
Metabolome Profiling of Eight Chinese Yam (Dioscorea polystachya Turcz.) Varieties Reveals Metabolite Diversity and Variety Specific Uses

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Sample Preparation and Extraction
2.3. UPLC Conditions and ESI-Q TRAP-MS/MS
2.4. Data Analyses
3. Results
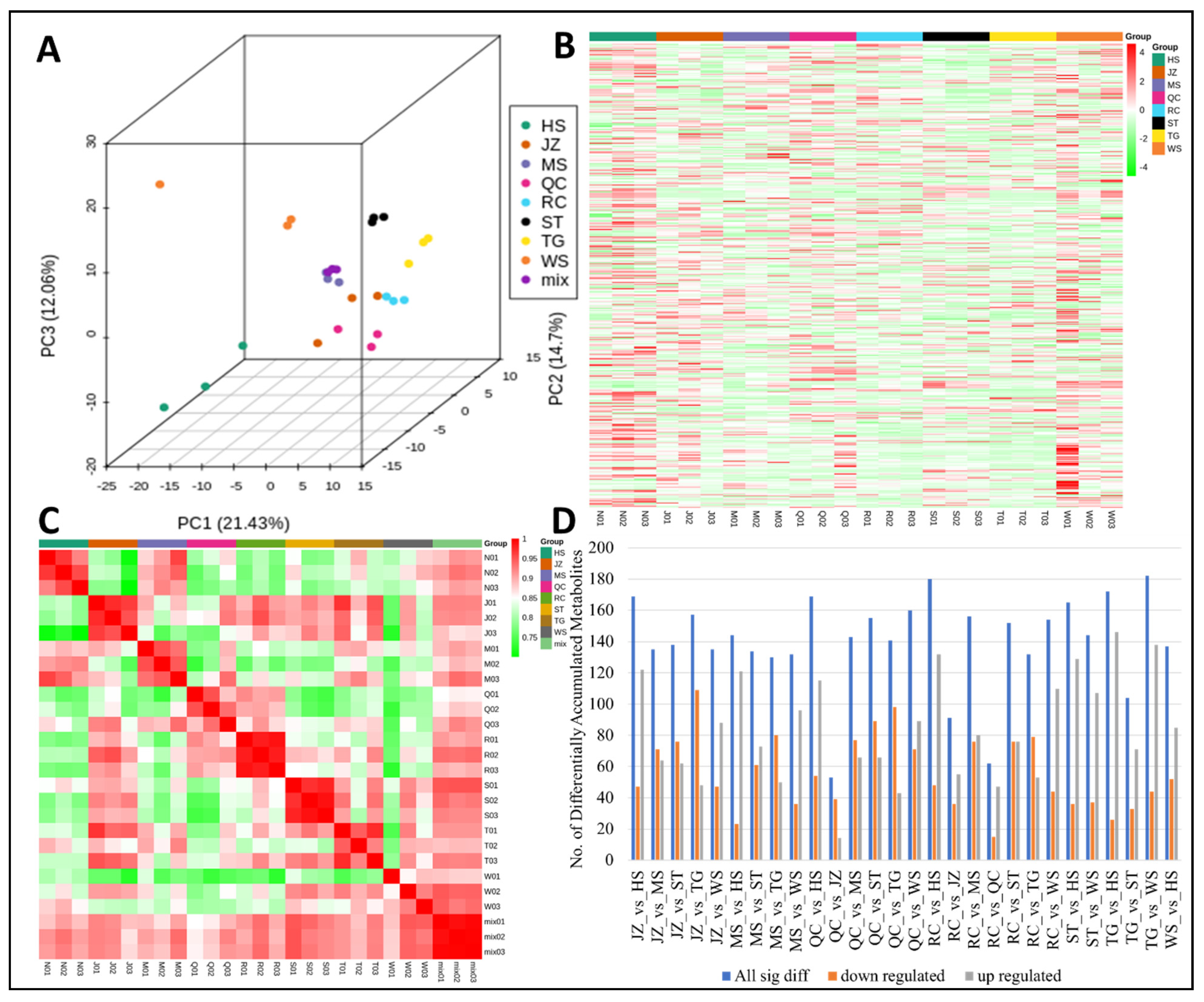
3.1. Metabolome Analysis
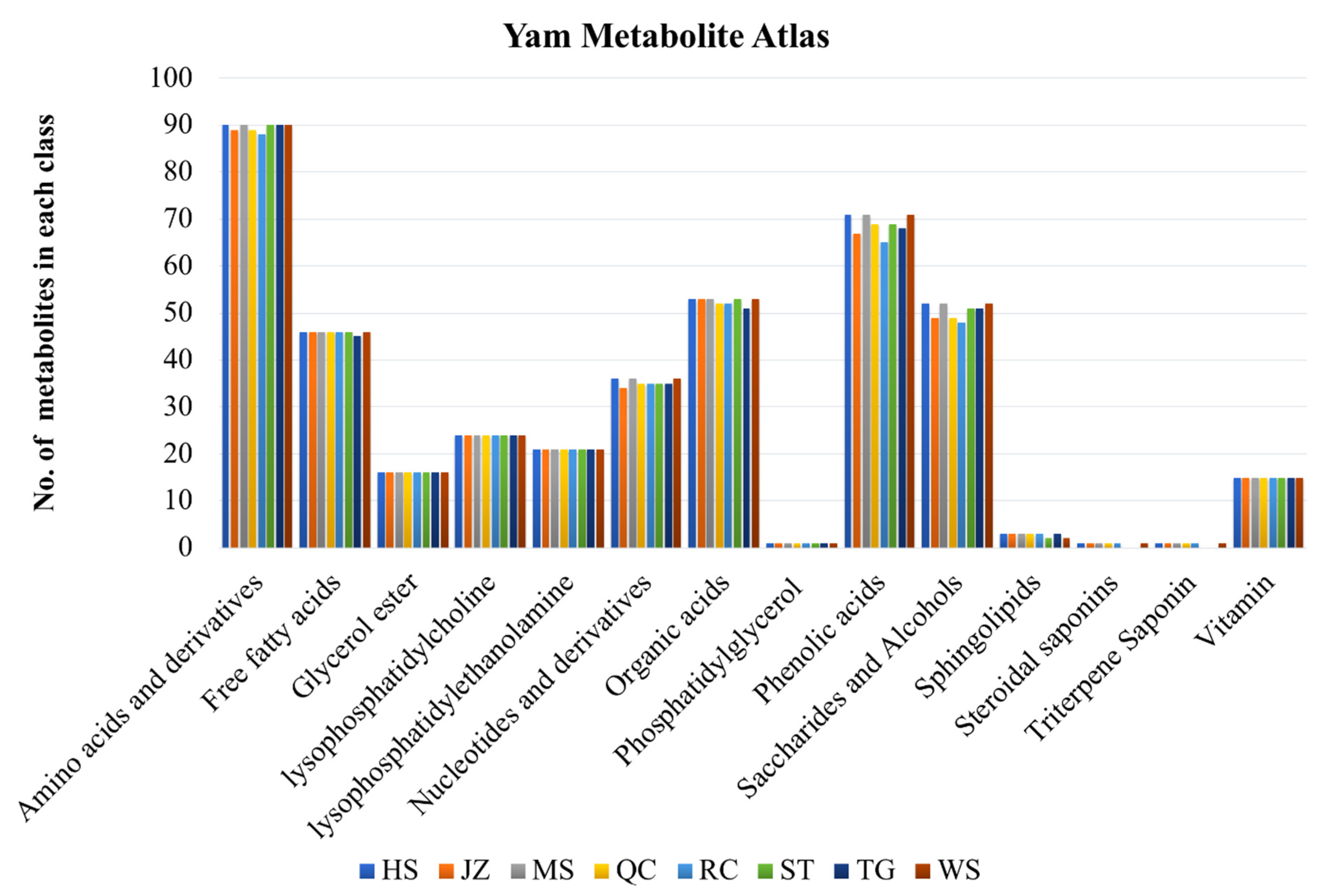
3.2. Yam Metabolite Atlas
3.3. Variety Demarcation and Specific Metabolites
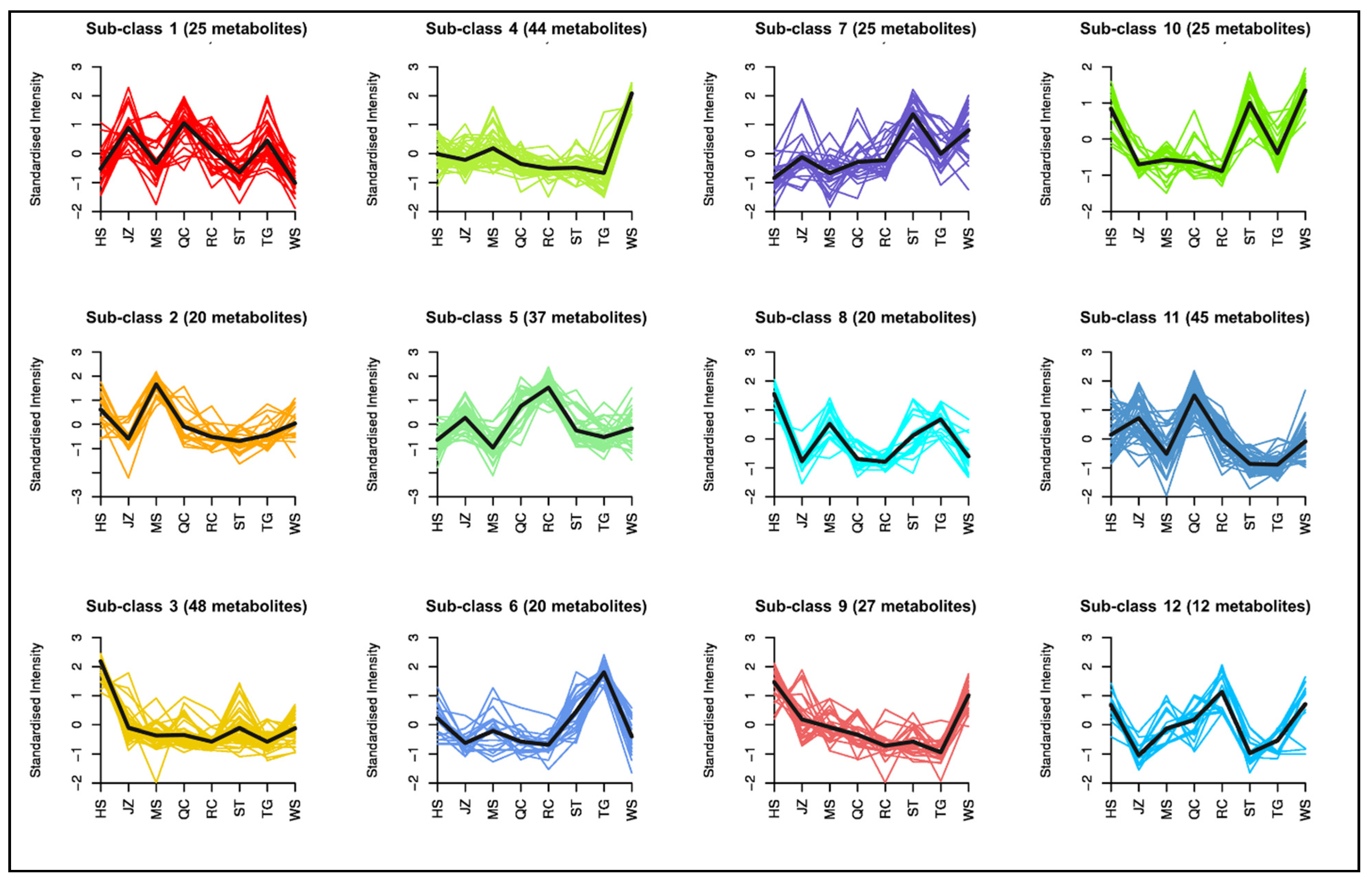
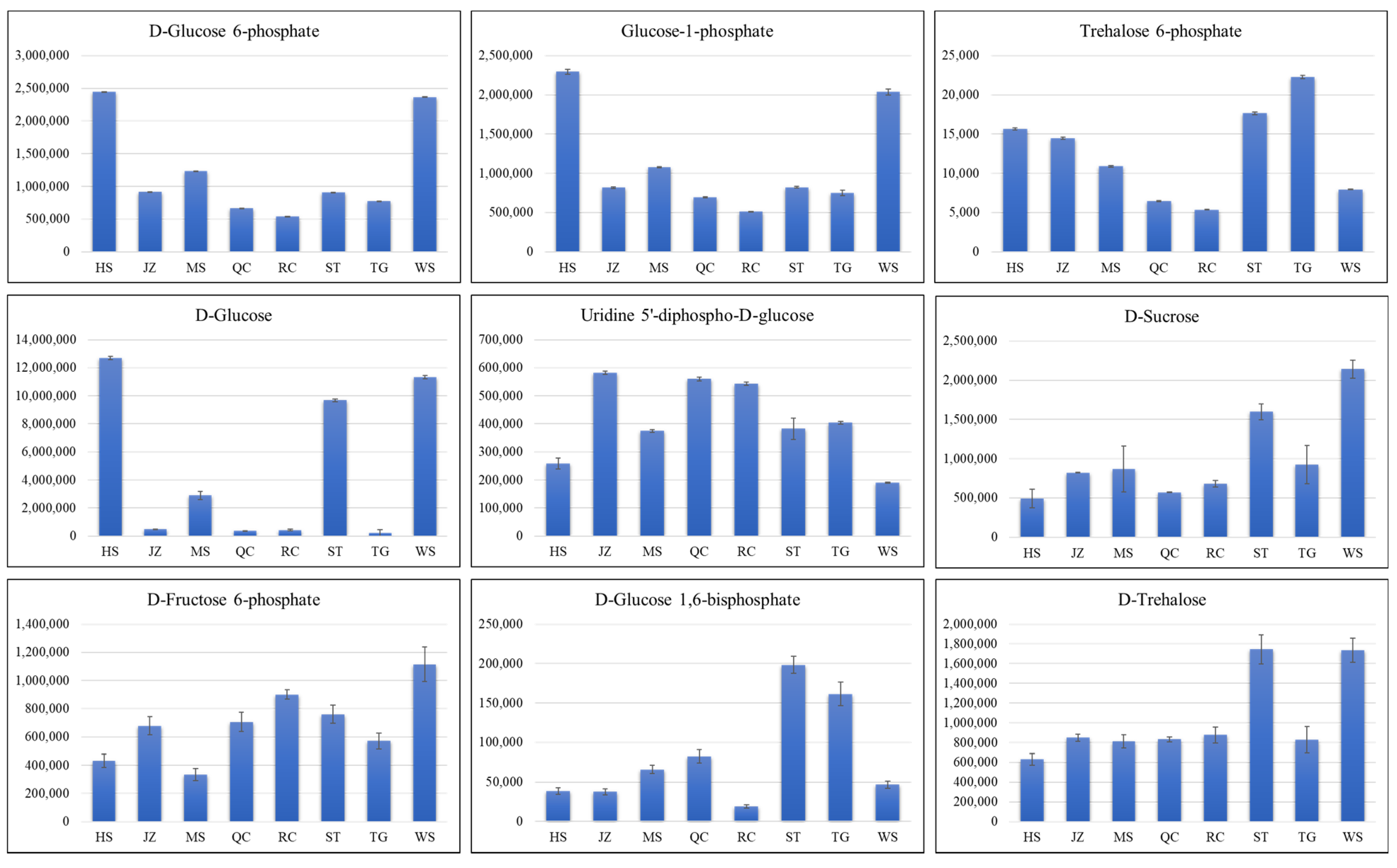
3.4. Accumulation of Metabolites Related to Amino Acid Biosynthesis Pathway
3.5. Key Medicinal Metabolites
4. Discussion
4.1. D. polystachya Is Rich in Amino Acids, Polysaccharides, and Phenolic Acids
4.2. D. polystachya Varieties Differ in Metabolome Profiles
4.3. Different D. polystachya Varieties Offer Different Utility Based on Metabolome Profiles and Contents
4.4. Future Research on D. polystachya
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coursey, D.G. Yams. An Account of the Nature, Origins, Cultivation and Utilisation of the Useful Members of the Dioscoreaceae; Humanities Press: New York, NY, USA, 1968. [Google Scholar]
- Zhang, L.; Bai, B.; Liu, X.; Wang, Y.; Li, M.; Zhao, D. α-Glucosidase inhibitors from Chinese yam (Dioscorea opposita Thunb.). Food Chem. 2011, 126, 203–206. [Google Scholar] [CrossRef]
- Zhang, Z.; Gao, W.; Wang, R.; Huang, L. Changes in main nutrients and medicinal composition of Chinese yam (Dioscorea opposita) tubers during storage. J. Food Sci. Technol. 2014, 51, 2535–2543. [Google Scholar] [CrossRef] [PubMed]
- Babil, P.; Kondo, S.; Iwata, H.; Kushikawa, S.; Shiwachi, H. Intra-specific ploidy variations in cultivated Chinese yam (Dioscorea polystachya Turcz.). Trop. Agric. Dev. 2013, 57, 101–107. [Google Scholar]
- Wu, Z.-G.; Jiang, W.; Nitin, M.; Bao, X.-Q.; Chen, S.-L.; Tao, Z.-M. Characterizing diversity based on nutritional and bioactive compositions of yam germplasm (Dioscorea spp.) commonly cultivated in China. J. Food Drug Anal. 2016, 24, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekara, A.; Josheph Kumar, T. Roots and tuber crops as functional foods: A review on phytochemical constituents and their potential health benefits. Int. J. Food Sci. 2016, 2016, 3631647. [Google Scholar] [CrossRef] [Green Version]
- Epping, J.; Laibach, N. An underutilized orphan tuber crop—Chinese yam: A review. Planta 2020, 252, 58. [Google Scholar]
- Sautour, M.; Mitaine-Offer, A.-C.; Lacaille-Dubois, M.-A. The Dioscorea genus: A review of bioactive steroid saponins. J. Nat. Med. 2007, 61, 91–101. [Google Scholar] [CrossRef]
- Leng, M.S.; Tobit, P.; Demasse, A.M.; Wolf, K.; Gouado, I.; Ndjouenkeu, R.; Rawel, H.M.; Schweigert, F.J. Nutritional and anti-oxidant properties of yam (Dioscorea schimperiana) based complementary food formulation. Sci. Afr. 2019, 5, e00132. [Google Scholar]
- Kanu, A.N.; Ezeocha, C.V.; Ogunka, N.P. A review on bioactive compounds of yam varieties for human disease management. Asian Food Sci. J. 2018, 1–10. [Google Scholar] [CrossRef]
- Yang, M.H.; Chin, Y.-W.; Yoon, K.D.; Kim, J. Phenolic compounds with pancreatic lipase inhibitory activity from Korean yam (Dioscorea opposita). J. Enzym. Inhib. Med. Chem. 2014, 29, 1–6. [Google Scholar] [CrossRef]
- Fan, Y.; He, Q.; Luo, A.; Wang, M.; Luo, A. Characterization and antihyperglycemic activity of a polysaccharide from Dioscorea opposita Thunb roots. Int. J. Mol. Sci. 2015, 16, 6391–6401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Hu, M.; Tao, J.; Yang, H.; Yan, P.; An, G.; Wang, H. The protective effects of Chinese yam polysaccharide against obesity-induced insulin resistance. J. Funct. Foods 2019, 55, 238–247. [Google Scholar] [CrossRef]
- Singh, K.; Kaushal, R. Comprehensive notes on commercial utilization, characteristics and status of steroid yielding plants in India. Ethnobot. Leafl. 2007, 2007, 9. [Google Scholar]
- Eymard, M. Composition Based on Diosgenin Ester for Topical Use. U.S. Patent Application 10/497,030, 10 February 2005. [Google Scholar]
- Price, E.J.; Bhattacharjee, R.; Lopez-Montes, A.; Fraser, P.D. Metabolite profiling of yam (Dioscorea spp.) accessions for use in crop improvement programmes. Metabolomics 2017, 13, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bino, R.J.; Hall, R.D.; Fiehn, O.; Kopka, J.; Saito, K.; Draper, J.; Nikolau, B.J.; Mendes, P.; Roessner-Tunali, U.; Beale, M.H. Potential of metabolomics as a functional genomics tool. Trends Plant Sci. 2004, 9, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Price, E.J.; Wilkin, P.; Sarasan, V.; Fraser, P.D. Metabolite profiling of Dioscorea (yam) species reveals underutilised biodiversity and renewable sources for high-value compounds. Sci. Rep. 2016, 6, 29136. [Google Scholar] [CrossRef] [Green Version]
- Champagne, A.; Bernillon, S.; Moing, A.; Rolin, D.; Legendre, L.; Lebot, V. Carotenoid profiling of tropical root crop chemotypes from Vanuatu, South Pacific. J. Food Compos. Anal. 2010, 23, 763–771. [Google Scholar] [CrossRef]
- Champagne, A.; Hilbert, G.; Legendre, L.; Lebot, V. Diversity of anthocyanins and other phenolic compounds among tropical root crops from Vanuatu, South Pacific. J. Food Compos. Anal. 2011, 24, 315–325. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Color attributes, betacyanin, and carotenoid profiles, bioactive components, and radical quenching capacity in selected Amaranthus gangeticus leafy vegetables. Sci. Rep. 2021, 11, 11559. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Leaf pigmentation, its profiles and radical scavenging activity in selected Amaranthus tricolor leafy vegetables. Sci. Rep. 2020, 10, 18617. [Google Scholar] [CrossRef]
- Sarker, U.; Hossain, M.N.; Iqbal, M.A.; Oba, S. Bioactive components and radical scavenging activity in selected advance lines of salt-tolerant vegetable amaranth. Front. Nutr. 2020, 7, 587257. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Nutritional and bioactive constituents and scavenging capacity of radicals in Amaranthus hypochondriacus. Sci. Rep. 2020, 10, 19962. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Phenolic profiles and antioxidant activities in selected drought-tolerant leafy vegetable amaranth. Sci. Rep. 2020, 10, 18287. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Nutraceuticals, phytochemicals, and radical quenching ability of selected drought-tolerant advance lines of vegetable amaranth. BMC Plant Biol. 2020, 20, 564. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S.; Daramy, M.A. Nutrients, minerals, antioxidant pigments and phytochemicals, and antioxidant capacity of the leaves of stem amaranth. Sci. Rep. 2020, 10, 3892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Oba, S. Nutrients, minerals, pigments, phytochemicals, and radical scavenging activity in Amaranthus blitum leafy vegetables. Sci. Rep. 2020, 10, 3868. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Hossain, M.M.; Oba, S. Nutritional and antioxidant components and antioxidant capacity in green morph Amaranthus leafy vegetable. Sci. Rep. 2020, 10, 1336. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Nutraceuticals, antioxidant pigments, and phytochemicals in the leaves of Amaranthus spinosus and Amaranthus viridis weedy species. Sci. Rep. 2019, 9, 20413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Oba, S. Protein, dietary fiber, minerals, antioxidant pigments and phytochemicals, and antioxidant activity in selected red morph Amaranthus leafy vegetable. PLoS ONE 2019, 14, e0222517. [Google Scholar]
- Abang, M.M.; Abraham, W.-R.; Asiedu, R.; Hoffmann, P.; Wolf, G.; Winter, S. Secondary metabolite profile and phytotoxic activity of genetically distinct forms of Colletotrichum gloeosporioides from yam (Dioscorea spp.). Mycol. Res. 2009, 113, 130–140. [Google Scholar] [CrossRef]
- Sheikh, N.; Kumar, Y.; Misra, A.; Pfoze, L. Phytochemical screening to validate the ethnobotanical importance of root tubers of Dioscorea species of Meghalaya, North East India. J. Med. Plants 2013, 1, 62–69. [Google Scholar]
- Barman, P.; Bhat, K.; Geeta, R. Phylogenetic analysis of Indian Dioscorea and comparison of secondary metabolite content with sampling across the tree. Genet. Resour. Crop Evol. 2018, 65, 1003–1012. [Google Scholar] [CrossRef]
- Kumar, S.; Mahanti, P.; Singh, N.R.; Rath, S.K.; Jena, P.K.; Patra, J.K. Antioxidant activity, antibacterial potential and characterization of active fraction of Dioscorea pentaphylla L. tuber extract collected from Similipal Biosphere Reserve, Odisha, India. Braz. J. Pharm. Sci. 2018, 53. [Google Scholar] [CrossRef]
- Lebot, V.; Malapa, R.; Abraham, K.; Molisalé, T.; Van Kien, N.; Gueye, B.; Waki, J. Secondary metabolites content may clarify the traditional selection process of the greater yam cultivars (Dioscorea alata L.). Genet. Resour. Crop Evol. 2018, 65, 1699–1709. [Google Scholar] [CrossRef]
- Price, E.J.; Drapal, M.; Perez-Fons, L.; Amah, D.; Bhattacharjee, R.; Heider, B.; Rouard, M.; Swennen, R.; Becerra Lopez-Lavalle, L.A.; Fraser, P.D. Metabolite database for root, tuber, and banana crops to facilitate modern breeding in understudied crops. Plant J. 2020, 101, 1258–1268. [Google Scholar] [CrossRef]
- Tan, L.; Jin, Z.; Ge, Y.; Nadeem, H.; Cheng, Z.; Azeem, F.; Zhan, R. Comprehensive ESI-Q TRAP-MS/MS based characterization of metabolome of two mango (Mangifera indica L) cultivars from China. Sci. Rep. 2020, 10, 20017. [Google Scholar] [CrossRef]
- Fraga, C.G.; Clowers, B.H.; Moore, R.J.; Zink, E.M. Signature-discovery approach for sample matching of a nerve-agent precursor using liquid chromatography—Mass spectrometry, XCMS, and chemometrics. Anal. Chem. 2010, 82, 4165–4173. [Google Scholar] [CrossRef]
- Rusilowicz, M.J.; Dickinson, M.; Charlton, A.J.; O’Keefe, S.; Wilson, J. MetaboClust: Using interactive time-series cluster analysis to relate metabolomic data with perturbed pathways. PLoS ONE 2018, 13, e0205968. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.-M.; Li, Y.; Zou, J.-H.; Guo, S.-Y.; Wang, F.; Yu, P.; Su, X.-J. Influence of Adding Chinese Yam (Dioscorea opposita Thunb.) Flour on Dough Rheology, Gluten Structure, Baking Performance, and Antioxidant Properties of Bread. Foods 2020, 9, 256. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Xie, J.; Yu, Y.; Shen, M. Recent progress in the research of yam mucilage polysaccharides: Isolation, structure and bioactivities. Int. J. Biol. Macromol. 2019. [Google Scholar] [CrossRef]
- Ketiku, A.; Oyenuga, V. Changes in the carbohydrate constituents of yam tuber (Dioscorea rotundata, Poir) during growth. J. Sci. Food Agric. 1973, 24, 367–373. [Google Scholar] [CrossRef]
- Doss, A.; Tresina, P.S.; Mohan, V.R. Amino acid composition of wild yam (Dioscorea spp.). Food Res. 2019, 3, 617–621. [Google Scholar] [CrossRef]
- Obidiegwu, J.E.; Lyons, J.B.; Chilaka, C.A. The Dioscorea Genus (Yam)—An Appraisal of Nutritional and Therapeutic Potentials. Foods 2020, 9, 1304. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, D.; Ji, H. Sucrose metabolism in developing oil-rich tubers of Cyperus esculentus: Comparative transcriptome analysis. BMC Plant Biol. 2018, 18, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamouz, K.; Lachman, J.; Hejtmánková, K.; Pazderů, K.; Čížek, M.; Dvořák, P. Effect of natural and growing conditions on the contentof phenolics in potatoes with different flesh colour. Plant Soil Environ. 2010, 56, 368–374. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Catalase, superoxide dismutase and ascorbate-glutathione cycle enzymes confer drought tolerance of Amaranthus tricolor. Sci. Rep. 2018, 8, 16496. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Response of nutrients, minerals, antioxidant leaf pigments, vitamins, polyphenol, flavonoid and antioxidant activity in selected vegetable amaranth under four soil water content. Food Chem. 2018, 252, 72–83. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. The response of salinity stress-induced A. tricolor to growth, anatomy, physiology, non-enzymatic and enzymatic antioxidants. Front. Plant Sci. 2020, 11, 559876. [Google Scholar] [CrossRef]
- Król, A.; Amarowicz, R.; Weidner, S. Changes in the composition of phenolic compounds and antioxidant properties of grapevine roots and leaves (Vitis vinifera L.) under continuous of long-term drought stress. Acta Physiol. Plant. 2014, 36, 1491–1499. [Google Scholar] [CrossRef] [Green Version]
- Romano, C.; Corsetti, G.; Flati, V.; Pasini, E.; Picca, A.; Calvani, R.; Marzetti, E.; Dioguardi, F.S. Influence of diets with varying essential/nonessential amino acid ratios on mouse lifespan. Nutrients 2019, 11, 1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tohda, C.; Yang, X.; Matsui, M.; Inada, Y.; Kadomoto, E.; Nakada, S.; Watari, H.; Shibahara, N. Diosgenin-rich yam extract enhances cognitive function: A placebo-controlled, randomized, double-blind, crossover study of healthy adults. Nutrients 2017, 9, 1160. [Google Scholar] [CrossRef]
- Jesus, M.; Martins, A.P.; Gallardo, E.; Silvestre, S. Diosgenin: Recent highlights on pharmacology and analytical methodology. J. Anal. Methods Chem. 2016, 2016, 4156293. [Google Scholar] [CrossRef] [PubMed]
- Adebowale, A.R.A.; Wahab, A.B.; Sobukola, P.O.; Obadina, A.O.; Kajihausa, E.O.; Adegunwa, O.M.; Sanni, O.L.; Tomlins, K. The antinutritional and vitamin composition of high-quality yam flour as affected by yam specie, pretreatment, and drying method. Food Sci. Nutr. 2018, 6, 1985–1990. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | HS | JZ | MS | QC | RC | ST | TG | WS |
|---|---|---|---|---|---|---|---|---|
| 2-Hydroxy-2-methyl-3-oxobutanoic acid | 20,189 | 2774 | 20,750 | 13,369 | 9100 | 8501 | 9 | 29,304 |
| L-Asparagine | 512,580 | 246,787 | 438,293 | 212,913 | 903,170 | 162,117 | 314,913 | 991,700 |
| Shikimic acid | 280,557 | 10,818 | 325,290 | 9 | 9 | 108,597 | 166,289 | 1,528,013 |
| L-Glycine | 54,524 | 31,847 | 99,641 | 46,566 | 166,227 | 31,485 | 61,560 | 91,819 |
| L-Aspartic Acid | 179,627 | 383,653 | 173,230 | 292,570 | 454,543 | 276,557 | 400,117 | 298,600 |
| L-Leucine | 1,023,030 | 217,523 | 504,200 | 1,086,607 | 883,730 | 264,000 | 655,027 | 1,113,757 |
| L-Threonine | 8,872,567 | 4,996,267 | 6,290,300 | 7,236,100 | 12,276,333 | 7,484,467 | 5,347,000 | 9,397,000 |
| L-Tyrosine | 7,563,567 | 4,342,400 | 3,939,633 | 23,188,433 | 30,096,333 | 2,100,900 | 1,447,500 | 6,041,733 |
| L-Histidine | 3,163,367 | 5,547,900 | 2,477,800 | 10,705,967 | 12,294,433 | 4,492,767 | 2,030,200 | 5,124,167 |
| L-Valine | 4,869,233 | 936,337 | 2,946,067 | 3,401,400 | 3,674,300 | 917,830 | 2,068,267 | 3,873,667 |
| L-Isoleucine | 1,031,653 | 216,743 | 489,500 | 1,065,467 | 876,860 | 287,447 | 669,460 | 1,132,440 |
| L-Arginine | 1,092,357 | 1,852,067 | 1,098,087 | 1,773,967 | 1,945,767 | 684,817 | 860,083 | 413,363 |
| L-Tryptophan | 3,141,013 | 820,333 | 308,770 | 15,000,333 | 2,424,500 | 13,194,333 | 3,090,397 | 9,606,433 |
| L-Homoserine | 9,104,733 | 5,182,267 | 6,424,467 | 7,368,867 | 1,295,6333 | 7,814,533 | 5,731,633 | 9,846,433 |
| O-Acetylserine | 56,170 | 56,702 | 76,012 | 42,476 | 51,966 | 23,477 | 35,622 | 66,658 |
| Phosphoenolpyruvate | 68,188 | 66,762 | 47,585 | 42,489 | 77,714 | 89,989 | 73,858 | 64,938 |
| Argininosuccinic acid | 95,243 | 75,388 | 93,438 | 59,969 | 60,440 | 30,598 | 46,636 | 165,560 |
| 2-Isopropylmalic Acid | 1,018,967 | 75,689 | 507,123 | 160,350 | 133,218 | 992,057 | 677,057 | 296,380 |
| L-Proline | 798,060 | 868,233 | 527,353 | 1,769,780 | 1,407,867 | 199,073 | 249,720 | 730,737 |
| L-Citrulline | 4,798,100 | 7,407,300 | 4,684,833 | 2,993,233 | 11,067,267 | 8,424,600 | 5,464,200 | 1,092,600 |
| L-Serine | 8,318,867 | 4,711,400 | 7,853,300 | 5,979,333 | 9,210,933 | 4,236,267 | 4,128,400 | 9,583,200 |
| L-Glutamic acid | 17,475,000 | 38,746,667 | 18,312,667 | 32,689,667 | 40,976,000 | 38,307,667 | 28,774,000 | 31,668,333 |
| L-Phenylalanine | 3,713,100 | 1,881,600 | 2,317,600 | 13,634,900 | 5,681,867 | 667,383 | 768,980 | 5,905,200 |
| L-Lysine | 197,727 | 297,223 | 146,947 | 395,507 | 468,727 | 181,067 | 109,435 | 161,207 |
| N-Acetyl-L-glutamic acid | 113,515 | 377,400 | 361,547 | 226,463 | 466,277 | 272,503 | 443,220 | 159,013 |
| L-Glutamine | 188,893 | 267,073 | 148,850 | 399,223 | 439,100 | 166,707 | 107,900 | 150,788 |
| L-Cysteine | 36,773 | 81,414 | 39,362 | 50,923 | 49,264 | 34,895 | 95,566 | 35,369 |
| 2,6-Diaminooimelic acid | 546,180 | 464,830 | 250,790 | 645,430 | 1,786,833 | 1,150,607 | 656,837 | 850,843 |
| L-Methionine | 1,532,673 | 248,787 | 925,987 | 1,233,383 | 1,294,523 | 257,847 | 884,340 | 1,868,263 |
| L-Saccharopine | 17,265 | 18,701 | 18,410 | 23,905 | 26,602 | 46,919 | 33,352 | 25,745 |
| L-Alanine | 7673 | 2188 | 17,661 | 6898 | 10,939 | 2125 | 3706 | 4032 |
| α-Ketoglutaric acid | 178,203 | 136,004 | 109,423 | 90,059 | 117,576 | 107,107 | 30,908 | 129,640 |
| L-Ornithine | 135,533 | 293,903 | 97,413 | 249,387 | 418,970 | 105,368 | 64,881 | 31,494 |
| S-Adenosyl-L-methionine | 42,219 | 19,932 | 26,620 | 34,835 | 30,260 | 19,435 | 14,517 | 16,205 |
| L-Cystathionine | 283,277 | 607,923 | 328,547 | 1,127,953 | 1,119,487 | 1,298,530 | 641,040 | 558,327 |
| N-α-Acetyl-L-ornithine | 6,323,067 | 17,055,667 | 6,126,967 | 13,141,000 | 18,058,667 | 4,866,900 | 5,826,533 | 1,909,933 |
| Dihydroxyacetone phosphate | 90,476 | 14,331 | 29,318 | 14,343 | 11,172 | 38,319 | 21,761 | 37,559 |
| D-Erythrose-4-phosphate | 31,412 | 6884 | 14,192 | 6210 | 4911 | 13,696 | 6532 | 21,288 |
| Compounds | HS | JZ | MS | QC | RC | ST | TG | WS |
|---|---|---|---|---|---|---|---|---|
| Riboflavin (Vitamin B2) | 3124 | 1860 | 2374 | 2300 | 2087 | 2722 | 1953 | 2406 |
| N-(beta-D-Glucosyl)nicotinate | 9360 | 8849 | 4451 | 7140 | 4050 | 3886 | 6298 | 4020 |
| L-Ascorbic acid (Vitamin C) | 52,139 | 3397 | 60,939 | 1751 | 1590 | 192,033 | 290,330 | 6761 |
| Biotin | 7817 | 12,808 | 12,286 | 11,324 | 14,041 | 10,141 | 8462 | 8709 |
| Delta-Tocopherol | 10,716 | 11,074 | 11,677 | 11,140 | 10,617 | 11,789 | 11,365 | 11,465 |
| Pyridoxal | 9895 | 15,313 | 8158 | 7995 | 7228 | 6595 | 6298 | 12,730 |
| Nicotinic acid (Vitamin B3) | 13,554 | 13,904 | 24,652 | 20,598 | 23,574 | 29,894 | 26,770 | 18,077 |
| Orotic acid | 496,593 | 37,893 | 43,963 | 23,132 | 16,767 | 213,033 | 46,264 | 58,923 |
| Pyridoxine-5′-O-glucoside | 244,953 | 446,547 | 131,430 | 367,150 | 129,638 | 193,710 | 187,913 | 94,560 |
| Menatetrenone (Vitamin K2) | 107,573 | 115,347 | 113,775 | 91,789 | 90,104 | 89,383 | 93,762 | 99,026 |
| Dehydroascorbic acid | 156,740 | 371,043 | 210,967 | 422,020 | 208,047 | 212,207 | 267,823 | 194,550 |
| Nicotinamide | 263,703 | 355,080 | 573,840 | 340,820 | 275,997 | 288,603 | 1,520,000 | 295,040 |
| Pyridoxine | 17,721 | 68,595 | 1,215,400 | 54,216 | 35,292 | 64,701 | 461,267 | 1,143,010 |
| D-Pantothenic acid | 1,330,137 | 1,817,033 | 2,018,067 | 2,388,933 | 1,475,200 | 2,063,467 | 2,351,233 | 1,976,033 |
| Nicotinate D-ribonucleoside | 1,136,437 | 2,357,433 | 2,083,867 | 2,711,567 | 1,604,133 | 3,770,933 | 3,500,067 | 2,302,200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, X.; Liu, D.; Huang, L. Metabolome Profiling of Eight Chinese Yam (Dioscorea polystachya Turcz.) Varieties Reveals Metabolite Diversity and Variety Specific Uses. Life 2021, 11, 687. https://doi.org/10.3390/life11070687
Zeng X, Liu D, Huang L. Metabolome Profiling of Eight Chinese Yam (Dioscorea polystachya Turcz.) Varieties Reveals Metabolite Diversity and Variety Specific Uses. Life. 2021; 11(7):687. https://doi.org/10.3390/life11070687
Chicago/Turabian StyleZeng, Xiaoxuan, Dahui Liu, and Luqi Huang. 2021. "Metabolome Profiling of Eight Chinese Yam (Dioscorea polystachya Turcz.) Varieties Reveals Metabolite Diversity and Variety Specific Uses" Life 11, no. 7: 687. https://doi.org/10.3390/life11070687
APA StyleZeng, X., Liu, D., & Huang, L. (2021). Metabolome Profiling of Eight Chinese Yam (Dioscorea polystachya Turcz.) Varieties Reveals Metabolite Diversity and Variety Specific Uses. Life, 11(7), 687. https://doi.org/10.3390/life11070687