
Dystonia and Cerebellum: From Bench to Bedside

 ,
,  , and
, and 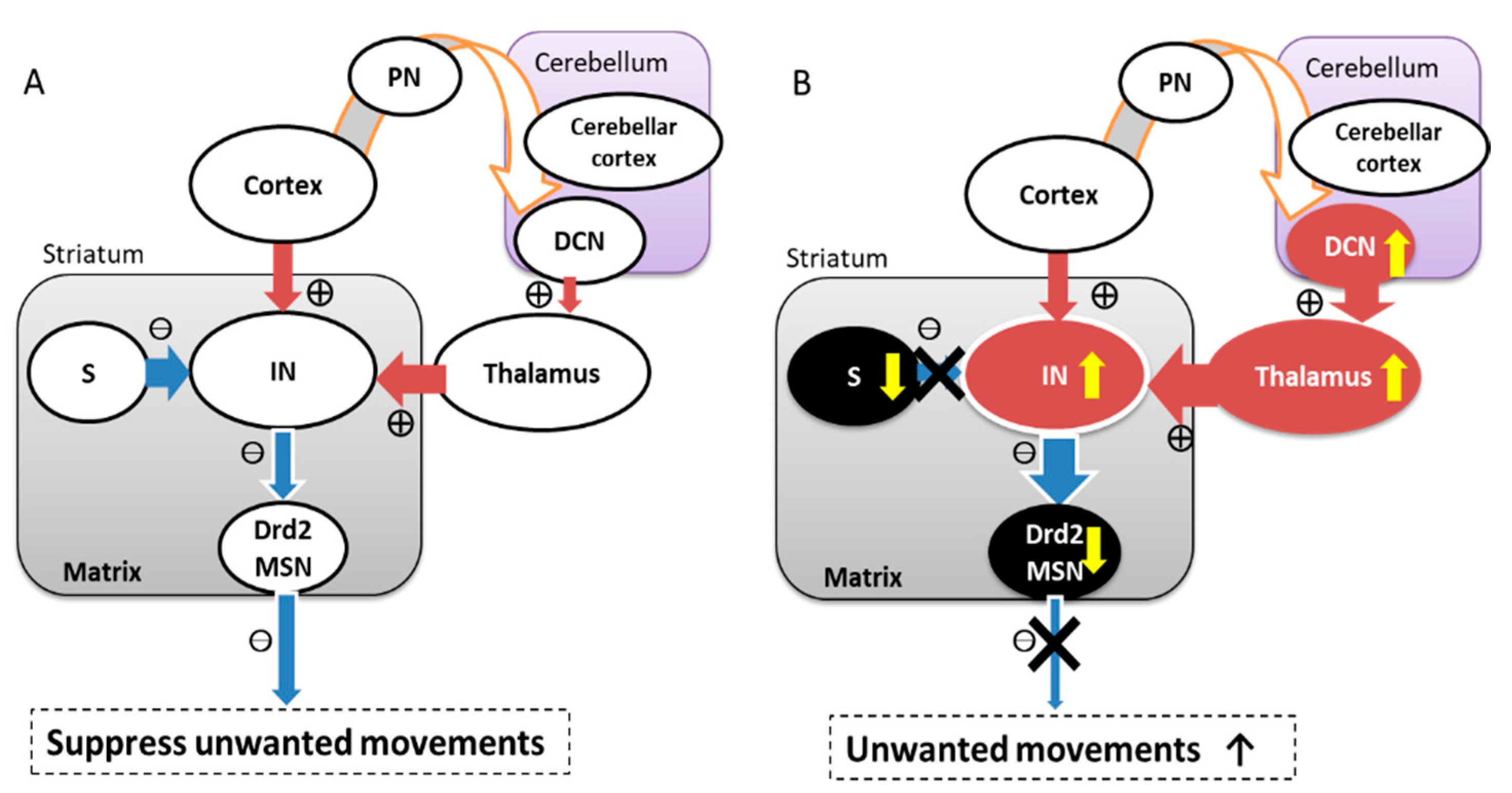{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Neuroanatomical Consideration: Interaction between Basal Ganglia and the Cerebellum
3. Research Regarding the Role of the Cerebellum in Dystonia Genesis
3.1. Evidence from Animal Models of Dystonia
3.2. Evidence from Clinical Research in Patients with Dystonia
3.3. The Effect of Cerebellar Stimulation for Dystonia
4. What Are the Roles of Two Distinct Loops?
5. Hypothesis for Dystonia Genesis
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jinnah, H.A.; Factor, S.A. Diagnosis and treatment of dystonia. Neurol. Clin. 2015, 33, 77–100. [Google Scholar] [CrossRef] [Green Version]
- Albanese, A.; Asmus, F.; Bhatia, K.P.; Elia, A.E.; Elibol, B.; Filippini, G.; Gasser, T.; Krauss, J.K.; Nardocci, N.; Newton, A.; et al. EFNS guidelines on diagnosis and treatment of primary dystonias. Eur. J. Neurol. 2011, 18, 5–18. [Google Scholar] [CrossRef]
- Geyer, H.L.; Bressman, S.B. The diagnosis of dystonia. Lancet Neurol. 2006, 5, 780–790. [Google Scholar] [CrossRef]
- Kaji, R.; Hasegawa, K.; Ugawa, Y.; Osawa, M.; Kashihara, K.; Kawarai, T.; Kobayashi, T.; Sakamoto, T.; Taira, T.; Tamagawa, S.; et al. Practical Guideline for Dystonia 2018, 1st ed.; Nankodo Co., Ltd.: Tokyo, Japan, 2018. [Google Scholar]
- Latorre, A.; Rocchi, L.; Bhatia, K.P. Delineating the electrophysiological signature of dystonia. Exp. Brain Res. 2020, 238, 1685–1692. [Google Scholar] [CrossRef] [PubMed]
- Kaji, R.; Bhatia, K.; Graybiel, A.M. Pathogenesis of dystonia: Is it of cerebellar or basal ganglia origin? J. Neurol. Neurosurg. Psychiatry 2018, 89, 488–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jinnah, H.A.; Hess, E.J. A new twist on the anatomy of dystonia: The basal ganglia and the cerebellum? Neurology 2006, 67, 1740–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neychev, V.K.; Fan, X.; Mitev, V.I.; Hess, E.J.; Jinnah, H.A. The basal ganglia and cerebellum interact in the expression of dystonic movement. Brain 2008, 131, 2499–2509. [Google Scholar] [CrossRef] [PubMed]
- Percheron, G.; Francois, C.; Talbi, B.; Yelnik, J.; Fenelon, G. The primate motor thalamus. Brain Res. Brain Res. Rev. 1996, 22, 93–181. [Google Scholar] [CrossRef]
- Voogd, J.; Glickstein, M. The anatomy of the cerebellum. Trends Cogn. Sci. 1998, 2, 307–313. [Google Scholar] [CrossRef]
- Grimaldi, G.; Manto, M. Topography of cerebellar deficits in humans. Cerebellum 2012, 11, 336–351. [Google Scholar] [CrossRef]
- Miterko, L.N.; Baker, K.B.; Beckinghausen, J.; Bradnam, L.V.; Cheng, M.Y.; Cooperrider, J.; DeLong, M.R.; Gornati, S.V.; Hallett, M.; Heck, D.H.; et al. Consensus paper: Experimental neurostimulation of the cerebellum. Cerebellum 2019, 18, 1064–1097. [Google Scholar] [CrossRef] [Green Version]
- Voogd, J. The human cerebellum. J. Chem. Neuroanat. 2003, 26, 243–252. [Google Scholar] [CrossRef]
- Pong, M.; Horn, K.M.; Gibson, A.R. Pathways for control of face and neck musculature by the basal ganglia and cerebellum. Brain Res. Rev. 2008, 58, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Manto, M. Mechanisms of human cerebellar dysmetria: Experimental evidence and current conceptual bases. J. Neuroeng. Rehabil. 2009, 6, 10. [Google Scholar] [CrossRef]
- Thach, W.T.; Goodkin, H.P.; Keating, J.G. The cerebellum and the adaptive coordination of movement. Annu. Rev. Neurosci. 1992, 15, 403–442. [Google Scholar] [CrossRef] [PubMed]
- Bond, K.M.; Brinjikji, W.; Eckel, L.J.; Kallmes, D.F.; McDonald, R.J.; Carr, C.M. Dentate update: Imaging features of entities that affect the dentate nucleus. AJNR Am. J. Neuroradiol. 2017, 38, 1467–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleine, J.F.; Guan, Y.; Kipiani, E.; Glonti, L.; Hoshi, M.; Buttner, U. Trunk position influences vestibular responses of fastigial nucleus neurons in the alert monkey. J. Neurophysiol. 2004, 91, 2090–2100. [Google Scholar] [CrossRef] [Green Version]
- Shaikh, A.G.; Meng, H.; Angelaki, D.E. Multiple reference frames for motion in the primate cerebellum. J. Neurosci. 2004, 24, 4491–4497. [Google Scholar] [CrossRef] [Green Version]
- Brooks, J.X.; Cullen, K.E. Multimodal integration in rostral fastigial nucleus provides an estimate of body movement. J. Neurosci. 2009, 29, 10499–10511. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Wang, J.J.; Zhu, J.N. Cerebellar fastigial nucleus: From anatomic construction to physiological functions. Cerebellum Ataxias 2016, 3, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshi, E.; Tremblay, L.; Feger, J.; Carras, P.L.; Strick, P.L. The cerebellum communicates with the basal ganglia. Nat. Neurosci. 2005, 8, 1491–1493. [Google Scholar] [CrossRef]
- Chen, C.H.; Fremont, R.; Arteaga-Bracho, E.E.; Khodakhah, K. Short latency cerebellar modulation of the basal ganglia. Nat. Neurosci. 2014, 17, 1767–1775. [Google Scholar] [CrossRef]
- Bostan, A.C.; Dum, R.P.; Strick, P.L. The basal ganglia communicate with the cerebellum. Proc. Natl. Acad. Sci. USA 2010, 107, 8452–8456. [Google Scholar] [CrossRef] [Green Version]
- Carta, I.; Chen, C.H.; Schott, A.L.; Dorizan, S.; Khodakhah, K. Cerebellar modulation of the reward circuitry and social behavior. Science 2019, 363. [Google Scholar] [CrossRef]
- Ratcheson, R.A.; Li, C.L. Effect of dentate stimulation on neuronal activity in the caudate nucleus. Exp. Neurol. 1969, 25, 268–281. [Google Scholar] [CrossRef]
- Raos, V.C.; Dermon, C.R.; Savaki, H.E. Functional anatomy of the thalamic centrolateral nucleus as revealed with the [14C] deoxyglucose method following electrical stimulation and electrolytic lesion. Neuroscience 1995, 68, 299–313. [Google Scholar] [CrossRef]
- Kim, J.E.; Chae, S.; Kim, S.; Jung, Y.J.; Kang, M.G.; Do Heo, W.; Kim, D. Cerebellar 5HT-2A receptor mediates stress-induced onset of dystonia. Sci. Adv. 2021, 7, eabb5735. [Google Scholar] [CrossRef] [PubMed]
- Ichinohe, N.; Mori, F.; Shoumura, K. A di-synaptic projection from the lateral cerebellar nucleus to the laterodorsal part of the striatum via the central lateral nucleus of the thalamus in the rat. Brain Res. 2000, 880, 191–197. [Google Scholar] [CrossRef]
- Ding, J.B.; Guzman, J.N.; Peterson, J.D.; Goldberg, J.A.; Surmeier, D.J. Thalamic gating of corticostriatal signaling by cholinergic interneurons. Neuron 2010, 67, 294–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doig, N.M.; Magill, P.J.; Apicella, P.; Bolam, J.P.; Sharott, A. Cortical and thalamic excitation mediate the multiphasic responses of striatal cholinergic interneurons to motivationally salient stimuli. J. Neurosci. 2014, 34, 3101–3117. [Google Scholar] [CrossRef] [Green Version]
- Assous, M.; Kaminer, J.; Shah, F.; Garg, A.; Koos, T.; Tepper, J.M. Differential processing of thalamic information via distinct striatal interneuron circuits. Nat. Commun. 2017, 8, 15860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapper, S.R.; Bolam, J.P. Input from the frontal cortex and the parafascicular nucleus to cholinergic interneurons in the dorsal striatum of the rat. Neuroscience 1992, 51, 533–545. [Google Scholar] [CrossRef]
- Meredith, G.E.; Wouterlood, F.G. Hippocampal and midline thalamic fibers and terminals in relation to the choline acetyltransferase-immunoreactive neurons in nucleus accumbens of the rat: A light and electron microscopic study. J. Comp. Neurol. 1990, 296, 204–221. [Google Scholar] [CrossRef] [PubMed]
- Sciamanna, G.; Ponterio, G.; Mandolesi, G.; Bonsi, P.; Pisani, A. Optogenetic stimulation reveals distinct modulatory properties of thalamostriatal vs corticostriatal glutamatergic inputs to fast-spiking interneurons. Sci. Rep. 2015, 5, 16742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bostan, A.C.; Strick, P.L. The cerebellum and basal ganglia are interconnected. Neuropsychol. Rev. 2010, 20, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, M.B.; Carleton, S.C.; Keller, J.T.; Conte, P. Connections of the subthalamic nucleus in the monkey. Brain Res. 1981, 224, 1–29. [Google Scholar] [CrossRef]
- Carpenter, M.B.; Baton, R.R., 3rd; Carleton, S.C.; Keller, J.T. Interconnections and organization of pallidal and subthalamic nucleus neurons in the monkey. J. Comp. Neurol. 1981, 197, 579–603. [Google Scholar] [CrossRef]
- Giolli, R.A.; Gregory, K.M.; Suzuki, D.A.; Blanks, R.H.; Lui, F.; Betelak, K.F. Cortical and subcortical afferents to the nucleus reticularis tegmenti pontis and basal pontine nuclei in the macaque monkey. Vis. Neurosci. 2001, 18, 725–740. [Google Scholar] [CrossRef] [Green Version]
- Hendry, S.H.; Jones, E.G.; Graham, J. Thalamic relay nuclei for cerebellar and certain related fiber systems in the cat. J. Comp. Neurol. 1979, 185, 679–713. [Google Scholar] [CrossRef]
- Ilinsky, I.A.; Kultas-Ilinsky, K. An autoradiographic study of topographical relationships between pallidal and cerebellar projections to the cat thalamus. Exp. Brain Res. 1984, 54, 95–106. [Google Scholar] [CrossRef]
- Pelzer, E.A.; Hintzen, A.; Goldau, M.; von Cramon, D.Y.; Timmermann, L.; Tittgemeyer, M. Cerebellar networks with basal ganglia: Feasibility for tracking cerebello-pallidal and subthalamo-cerebellar projections in the human brain. Eur. J. Neurosci. 2013, 38, 3106–3114. [Google Scholar] [CrossRef] [PubMed]
- Asanuma, C.; Thach, W.R.; Jones, E.G. Anatomical evidence for segregated focal groupings of efferent cells and their terminal ramifications in the cerebellothalamic pathway of the monkey. Brain Res. 1983, 286, 267–297. [Google Scholar] [CrossRef]
- Middleton, F.A.; Strick, P.L. Anatomical evidence for cerebellar and basal ganglia involvement in higher cognitive function. Science 1994, 266, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Middleton, F.A.; Strick, P.L. Cerebellar output channels. Int. Rev. Neurobiol. 1997, 41, 61–82. [Google Scholar] [PubMed]
- Middleton, F.A.; Strick, P.L. Dentate output channels: Motor and cognitive components. Prog. Brain Res. 1997, 114, 553–566. [Google Scholar] [PubMed]
- Dum, R.P.; Li, C.; Strick, P.L. Motor and nonmotor domains in the monkey dentate. Ann. N. Y. Acad. Sci. 2002, 978, 289–301. [Google Scholar] [CrossRef]
- Ilinsky, I.A.; Kultas-Ilinsky, K. Sagittal cytoarchitectonic maps of the macaca mulatta thalamus with a revised nomenclature of the motor-related nuclei validated by observations on their connectivity. J. Comp. Neurol. 1987, 262, 331–364. [Google Scholar] [CrossRef]
- Nakano, K. Neural circuits and topographic organization of the basal ganglia and related regions. Brain Dev. 2000, 22 (Suppl. 1), S5–S16. [Google Scholar] [CrossRef]
- Kuo, J.S.; Carpenter, M.B. Organization of pallidothalamic projections in the rhesus monkey. J. Comp. Neurol. 1973, 151, 201–236. [Google Scholar] [CrossRef]
- Sidibe, M.; Bevan, M.D.; Bolam, J.P.; Smith, Y. Efferent connections of the internal globus pallidus in the squirrel monkey: I. Topography and synaptic organization of the pallidothalamic projection. J. Comp. Neurol. 1997, 382, 323–347. [Google Scholar] [CrossRef]
- Sakai, S.T.; Inase, M.; Tanji, J. Comparison of cerebellothalamic and pallidothalamic projections in the monkey (macaca fuscata): A double anterograde labeling study. J. Comp. Neurol. 1996, 368, 215–228. [Google Scholar] [CrossRef]
- Akkal, D.; Dum, R.P.; Strick, P.L. Supplementary motor area and presupplementary motor area: Targets of basal ganglia and cerebellar output. J. Neurosci. 2007, 27, 10659–10673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbruzzese, G.; Berardelli, A. Further progress in understanding the pathophysiology of primary dystonia. Mov. Disord. 2011, 26, 1185–1186. [Google Scholar] [CrossRef] [PubMed]
- Quartarone, A.; Cacciola, A.; Milardi, D.; Ghilardi, M.F.; Calamuneri, A.; Chillemi, G.; Anastasi, G.; Rothwell, J. New insights into cortico-basal-cerebellar connectome: Clinical and physiological considerations. Brain 2020, 143, 396–406. [Google Scholar] [CrossRef]
- Beier, K.T.; Gao, X.J.; Xie, S.; DeLoach, K.E.; Malenka, R.C.; Luo, L. Topological organization of ventral tegmental area connectivity revealed by viral-genetic dissection of input-output relations. Cell Rep. 2019, 26, 159–167.e6. [Google Scholar] [CrossRef] [Green Version]
- Beier, K.T.; Steinberg, E.E.; DeLoach, K.E.; Xie, S.; Miyamichi, K.; Schwarz, L.; Gao, X.J.; Kremer, E.J.; Malenka, R.C.; Luo, L. Circuit architecture of VTA dopamine neurons revealed by systematic input-output mapping. Cell 2015, 162, 622–634. [Google Scholar] [CrossRef] [Green Version]
- Trutti, A.C.; Mulder, M.J.; Hommel, B.; Forstmann, B.U. Functional neuroanatomical review of the ventral tegmental area. Neuroimage 2019, 191, 258–268. [Google Scholar] [CrossRef] [Green Version]
- Watabe-Uchida, M.; Zhu, L.; Ogawa, S.K.; Vamanrao, A.; Uchida, N. Whole-brain mapping of direct inputs to midbrain dopamine neurons. Neuron 2012, 74, 858–873. [Google Scholar] [CrossRef] [Green Version]
- Bourdy, R.; Sanchez-Catalan, M.J.; Kaufling, J.; Balcita-Pedicino, J.J.; Freund-Mercier, M.J.; Veinante, P.; Sesack, S.R.; Georges, F.; Barrot, M. Control of the nigrostriatal dopamine neuron activity and motor function by the tail of the ventral tegmental area. Neuropsychopharmacology 2014, 39, 2788–2798. [Google Scholar] [CrossRef] [Green Version]
- Flaherty, A.W.; Graybiel, A.M. Input-output organization of the sensorimotor striatum in the squirrel monkey. J. Neurosci. 1994, 14, 599–610. [Google Scholar] [CrossRef]
- Filip, P.; Lungu, O.V.; Bares, M. Dystonia and the cerebellum: A new field of interest in movement disorders? Clin. Neurophysiol. 2013, 124, 1269–1276. [Google Scholar] [CrossRef]
- Stratton, S.E.; Lorden, J.F. Effect of harmaline on cells of the inferior olive in the absence of tremor: Differential response of genetically dystonic and harmaline-tolerant rats. Neuroscience 1991, 41, 543–549. [Google Scholar] [CrossRef]
- Isaacs, K.R.; Abbott, L.C. Cerebellar volume decreases in the tottering mouse are specific to the molecular layer. Brain Res. Bull. 1995, 36, 309–314. [Google Scholar] [CrossRef]
- Heckroth, J.A.; Abbott, L.C. Purkinje cell loss from alternating sagittal zones in the cerebellum of leaner mutant mice. Brain Res. 1994, 658, 93–104. [Google Scholar] [CrossRef]
- Matsui, K.; Mukoyama, M.; Adachi, K.; Ando, K. Fundamental study on ataxic mice (wriggle mouse sagami). Jikken Dobutsu 1987, 36, 185–189. [Google Scholar] [PubMed] [Green Version]
- Fremont, R.; Tewari, A.; Angueyra, C.; Khodakhah, K. A role for cerebellum in the hereditary dystonia DYT1. eLife 2017, 6, e22775. [Google Scholar] [CrossRef] [PubMed]
- Raike, R.S.; Pizoli, C.E.; Weisz, C.; van den Maagdenberg, A.M.; Jinnah, H.A.; Hess, E.J. Limited regional cerebellar dysfunction induces focal dystonia in mice. Neurobiol. Dis. 2013, 49, 200–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, L.L.; Lorden, J.F. Regional cerebral glucose utilization reveals widespread abnormalities in the motor system of the rat mutant dystonic. J. Neurosci. 1989, 9, 4033–4041. [Google Scholar] [CrossRef] [Green Version]
- Calderon, D.P.; Fremont, R.; Kraenzlin, F.; Khodakhah, K. The neural substrates of rapid-onset dystonia-parkinsonism. Nat. Neurosci. 2011, 14, 357–365. [Google Scholar] [CrossRef]
- Ulug, A.M.; Vo, A.; Argyelan, M.; Tanabe, L.; Schiffer, W.K.; Dewey, S.; Dauer, W.T.; Eidelberg, D. Cerebellothalamocortical pathway abnormalities in torsinA DYT1 knock-in mice. Proc. Natl. Acad. Sci. USA 2011, 108, 6638–6643. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Sharma, N.; LeDoux, M.S. The DYT1 carrier state increases energy demand in the olivocerebellar network. Neuroscience 2011, 177, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Campbell, D.B.; Hess, E.J. Cerebellar circuitry is activated during convulsive episodes in the tottering (tg/tg) mutant mouse. Neuroscience 1998, 85, 773–783. [Google Scholar] [CrossRef]
- Campbell, D.B.; North, J.B.; Hess, E.J. Tottering mouse motor dysfunction is abolished on the purkinje cell degeneration (pcd) mutant background. Exp. Neurol. 1999, 160, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Devanagondi, R.; Egami, K.; LeDoux, M.S.; Hess, E.J.; Jinnah, H.A. Neuroanatomical substrates for paroxysmal dyskinesia in lethargic mice. Neurobiol. Dis. 2007, 27, 249–257. [Google Scholar] [CrossRef] [PubMed]
- LeDoux, M.S.; Lorden, J.F. Abnormal cerebellar output in the genetically dystonic rat. Adv. Neurol. 1998, 78, 63–78. [Google Scholar]
- LeDoux, M.S.; Hurst, D.C.; Lorden, J.F. Single-unit activity of cerebellar nuclear cells in the awake genetically dystonic rat. Neuroscience 1998, 86, 533–545. [Google Scholar] [CrossRef]
- Fremont, R.; Calderon, D.P.; Maleki, S.; Khodakhah, K. Abnormal high-frequency burst firing of cerebellar neurons in rapid-onset dystonia-parkinsonism. J. Neurosci. 2014, 34, 11723–11732. [Google Scholar] [CrossRef] [Green Version]
- Fremont, R.; Tewari, A.; Khodakhah, K. Aberrant purkinje cell activity is the cause of dystonia in a shRNA-based mouse model of rapid onset dystonia-parkinsonism. Neurobiol. Dis. 2015, 82, 200–212. [Google Scholar] [CrossRef] [Green Version]
- Stratton, S.E.; Lorden, J.F.; Mays, L.E.; Oltmans, G.A. Spontaneous and harmaline-stimulated purkinje cell activity in rats with a genetic movement disorder. J. Neurosci. 1988, 8, 3327–3336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J.; Ledoux, M.S. Caytaxin deficiency causes generalized dystonia in rats. Brain Res. Mol. Brain Res. 2005, 141, 181–192. [Google Scholar] [CrossRef]
- Alvarez-Fischer, D.; Grundmann, M.; Lu, L.; Samans, B.; Fritsch, B.; Moller, J.C.; Schaefer, M.K.; Hartmann, A.; Oertel, W.H.; Bandmann, O. Prolonged generalized dystonia after chronic cerebellar application of kainic acid. Brain Res. 2012, 1464, 82–88. [Google Scholar] [CrossRef]
- LeDoux, M.S.; Lorden, J.F.; Ervin, J.M. Cerebellectomy eliminates the motor syndrome of the genetically dystonic rat. Exp. Neurol. 1993, 120, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Hughes, K.E.; Jinnah, H.A.; Hess, E.J. Selective and sustained alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor activation in cerebellum induces dystonia in mice. J. Pharm. Exp. 2012, 340, 733–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizoli, C.E.; Jinnah, H.A.; Billingsley, M.L.; Hess, E.J. Abnormal cerebellar signaling induces dystonia in mice. J. Neurosci. 2002, 22, 7825–7833. [Google Scholar] [CrossRef] [Green Version]
- Ellen, J.H.; Jinnah, H.A. Mouse model of dystonia. In Animal Models of Movement Disorder, 1st ed.; LeDoux, M., Ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005; pp. 265–277. [Google Scholar]
- White, J.J.; Sillitoe, R.V. Genetic silencing of olivocerebellar synapses causes dystonia-like behaviour in mice. Nat. Commun. 2017, 8, 14912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeDoux, M.S.; Brady, K.A. Secondary cervical dystonia associated with structural lesions of the central nervous system. Mov. Disord. 2003, 18, 60–69. [Google Scholar] [CrossRef]
- Seidel, K.; Siswanto, S.; Brunt, E.R.; den Dunnen, W.; Korf, H.W.; Rub, U. Brain pathology of spinocerebellar ataxias. Acta Neuropathol. 2012, 124, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, N.A.; Stell, R.; Harding, A.E.; Marsden, C.D. Degenerative cerebellar ataxia and focal dystonia. Mov. Disord. 1988, 3, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Manto, M.U. The wide spectrum of spinocerebellar ataxias (SCAs). Cerebellum 2005, 4, 2–6. [Google Scholar] [CrossRef]
- Anheim, M.; Tranchant, C.; Koenig, M. The autosomal recessive cerebellar ataxias. N. Engl. J. Med. 2012, 366, 636–646. [Google Scholar] [CrossRef]
- Muzaimi, M.B.; Wiles, C.M.; Robertson, N.P.; Ravine, D.; Compston, D.A. Task specific focal dystonia: A presentation of spinocerebellar ataxia type 6. J. Neurol. Neurosurg. Psychiatry 2003, 74, 1444–1445. [Google Scholar] [CrossRef] [Green Version]
- Saunders-Pullman, R.; Raymond, D.; Stoessl, A.J.; Hobson, D.; Nakamura, K.; Pullman, S.; Lefton, D.; Okun, M.S.; Uitti, R.; Sachdev, R.; et al. Variant ataxia-telangiectasia presenting as primary-appearing dystonia in Canadian Mennonites. Neurology 2012, 78, 649–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodensteiner, J.B.; Goldblum, R.M.; Goldman, A.S. Progressive dystonia masking ataxia in ataxia-telangiectasia. Arch. Neurol. 1980, 37, 464–465. [Google Scholar] [CrossRef] [PubMed]
- Krauss, J.K.; Seeger, W.; Jankovic, J. Cervical dystonia associated with tumors of the posterior fossa. Mov. Disord. 1997, 12, 443–447. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, K.; O’Riordan, S.; Gallagher, J.; Hutchinson, M. Paroxysmal torticollis and blepharospasm following bilateral cerebellar infarction. J. Neurol. 2006, 253, 1644–1645. [Google Scholar] [CrossRef]
- Le Ber, I.; Clot, F.; Vercueil, L.; Camuzat, A.; Viemont, M.; Benamar, N.; De Liege, P.; Ouvrard-Hernandez, A.M.; Pollak, P.; Stevanin, G.; et al. Predominant dystonia with marked cerebellar atrophy: A rare phenotype in familial dystonia. Neurology 2006, 67, 1769–1773. [Google Scholar] [CrossRef]
- Miyamoto, R.; Sumikura, H.; Takeuchi, T.; Sanada, M.; Fujita, K.; Kawarai, T.; Mure, H.; Morigaki, R.; Goto, S.; Murayama, S.; et al. Autopsy case of severe generalized dystonia and static ataxia with marked cerebellar atrophy. Neurology 2015, 85, 1522–1524. [Google Scholar] [CrossRef]
- Hagenah, J.; Reetz, K.; Zuhlke, C.; Rolfs, A.; Binkofski, F.; Klein, C. Predominant dystonia with marked cerebellar atrophy: A rare phenotype in familial dystonia. Neurology 2006, 67, 1769–1773. [Google Scholar] [CrossRef]
- van de Warrenburg, B.P.; Giunti, P.; Schneider, S.A.; Quinn, N.P.; Wood, N.W.; Bhatia, K.P. The syndrome of (predominantly cervical) dystonia and cerebellar ataxia: New cases indicate a distinct but heterogeneous entity. J. Neurol. Neurosurg. Psychiatry 2007, 78, 774–775. [Google Scholar] [CrossRef] [Green Version]
- Batla, A.; Sanchez, M.C.; Erro, R.; Ganos, C.; Stamelou, M.; Balint, B.; Brugger, F.; Antelmi, E.; Bhatia, K.P. The role of cerebellum in patients with late onset cervical/segmental dystonia?--evidence from the clinic. Parkinsonism Relat. Disord. 2015, 21, 1317–1322. [Google Scholar] [CrossRef]
- Prudente, C.N.; Pardo, C.A.; Xiao, J.; Hanfelt, J.; Hess, E.J.; Ledoux, M.S.; Jinnah, H.A. Neuropathology of cervical dystonia. Exp. Neurol. 2013, 241, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Ma, K.; Babij, R.; Cortes, E.; Vonsattel, J.P.; Louis, E.D. Cerebellar pathology of a dual clinical diagnosis: Patients with essential tremor and dystonia. Tremor Other Hyperkinet Mov. 2012, 2, tre-12-107-677-1. [Google Scholar] [CrossRef]
- Rossi, M.; Perez-Lloret, S.; Cerquetti, D.; Merello, M. Movement disorders in autosomal dominant cerebellar ataxias: A systematic review. Mov. Disord. Clin. Pr. 2014, 1, 154–160. [Google Scholar] [CrossRef] [PubMed]
- van Gaalen, J.; Giunti, P.; van de Warrenburg, B.P. Movement disorders in spinocerebellar ataxias. Mov. Disord. 2011, 26, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Cancel, G.; Durr, A.; Didierjean, O.; Imbert, G.; Burk, K.; Lezin, A.; Belal, S.; Benomar, A.; Abada-Bendib, M.; Vial, C.; et al. Molecular and clinical correlations in spinocerebellar ataxia 2: A study of 32 families. Hum. Mol. Genet. 1997, 6, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.H.; Gan, S.R.; Wang, J.; Lo, R.Y.; Figueroa, K.P.; Tomishon, D.; Pulst, S.M.; Perlman, S.; Wilmot, G.; Gomez, C.M.; et al. Dystonia and ataxia progression in spinocerebellar ataxias. Parkinsonism Relat. Disord. 2017, 45, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Neychev, V.K.; Gross, R.E.; Lehericy, S.; Hess, E.J.; Jinnah, H.A. The functional neuroanatomy of dystonia. Neurobiol. Dis. 2011, 42, 185–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoons, E.; Booij, J.; Nederveen, A.J.; Dijk, J.M.; Tijssen, M.A. Structural, functional and molecular imaging of the brain in primary focal dystonia—A review. Neuroimage 2011, 56, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Tewari, A.; Fremont, R.; Khodakhah, K. It’s not just the basal ganglia: Cerebellum as a target for dystonia therapeutics. Mov. Disord. 2017, 32, 1537–1545. [Google Scholar] [CrossRef]
- Draganski, B.; Thun-Hohenstein, C.; Bogdahn, U.; Winkler, J.; May, A. "Motor circuit" gray matter changes in idiopathic cervical dystonia. Neurology 2003, 61, 1228–1231. [Google Scholar] [CrossRef]
- Obermann, M.; Yaldizli, O.; De Greiff, A.; Lachenmayer, M.L.; Buhl, A.R.; Tumczak, F.; Gizewski, E.R.; Diener, H.C.; Maschke, M. Morphometric changes of sensorimotor structures in focal dystonia. Mov. Disord. 2007, 22, 1117–1123. [Google Scholar] [CrossRef]
- Delmaire, C.; Vidailhet, M.; Elbaz, A.; Bourdain, F.; Bleton, J.P.; Sangla, S.; Meunier, S.; Terrier, A.; Lehericy, S. Structural abnormalities in the cerebellum and sensorimotor circuit in writer’s cramp. Neurology 2007, 69, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Prudente, C.N.; Hess, E.J.; Jinnah, H.A. Dystonia as a network disorder: What is the role of the cerebellum? Neuroscience 2014, 260, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argyelan, M.; Carbon, M.; Niethammer, M.; Ulug, A.M.; Voss, H.U.; Bressman, S.B.; Dhawan, V.; Eidelberg, D. Cerebellothalamocortical connectivity regulates penetrance in dystonia. J. Neurosci. 2009, 29, 9740–9747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbon, M.; Kingsley, P.B.; Tang, C.; Bressman, S.; Eidelberg, D. Microstructural white matter changes in primary torsion dystonia. Mov. Disord. 2008, 23, 234–239. [Google Scholar] [CrossRef]
- Eidelberg, D.; Moeller, J.R.; Antonini, A.; Kazumata, K.; Nakamura, T.; Dhawan, V.; Spetsieris, P.; deLeon, D.; Bressman, S.B.; Fahn, S. Functional brain networks in DYT1 dystonia. Ann. Neurol. 1998, 44, 303–312. [Google Scholar] [CrossRef]
- Ceballos-Baumann, A.O.; Passingham, R.E.; Marsden, C.D.; Brooks, D.J. Motor reorganization in acquired hemidystonia. Ann. Neurol. 1995, 37, 746–757. [Google Scholar] [CrossRef] [PubMed]
- Kluge, A.; Kettner, B.; Zschenderlein, R.; Sandrock, D.; Munz, D.L.; Hesse, S.; Meierkord, H. Changes in perfusion pattern using ecd-spect indicate frontal lobe and cerebellar involvement in exercise-induced paroxysmal dystonia. Mov. Disord. 1998, 13, 125–134. [Google Scholar] [CrossRef]
- Odergren, T.; Stone-Elander, S.; Ingvar, M. Cerebral and cerebellar activation in correlation to the action-induced dystonia in writer’s cramp. Mov. Disord. 1998, 13, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Preibisch, C.; Berg, D.; Hofmann, E.; Solymosi, L.; Naumann, M. Cerebral activation patterns in patients with writer’s cramp: A functional magnetic resonance imaging study. J. Neurol. 2001, 248, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Galardi, G.; Perani, D.; Grassi, F.; Bressi, S.; Amadio, S.; Antoni, M.; Comi, G.C.; Canal, N.; Fazio, F. Basal ganglia and thalamo-cortical hypermetabolism in patients with spasmodic torticollis. Acta Neurol. Scand. 1996, 94, 172–176. [Google Scholar] [CrossRef]
- Hutchinson, M.; Nakamura, T.; Moeller, J.R.; Antonini, A.; Belakhlef, A.; Dhawan, V.; Eidelberg, D. The metabolic topography of essential blepharospasm: A focal dystonia with general implications. Neurology 2000, 55, 673–677. [Google Scholar] [CrossRef] [Green Version]
- Carbon, M.; Raymond, D.; Ozelius, L.; Saunders-Pullman, R.; Frucht, S.; Dhawan, V.; Bressman, S.; Eidelberg, D. Metabolic changes in DYT11 myoclonus-dystonia. Neurology 2013, 80, 385–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehericy, S.; Gerardin, E.; Poline, J.B.; Meunier, S.; Van de Moortele, P.F.; Le Bihan, D.; Vidailhet, M. Motor execution and imagination networks in post-stroke dystonia. Neuroreport 2004, 15, 1887–1890. [Google Scholar] [CrossRef]
- Asanuma, K.; Ma, Y.; Huang, C.; Carbon-Correll, M.; Edwards, C.; Raymond, D.; Bressman, S.B.; Moeller, J.R.; Eidelberg, D. The metabolic pathology of dopa-responsive dystonia. Ann. Neurol. 2005, 57, 596–600. [Google Scholar] [CrossRef]
- Thobois, S.; Ballanger, B.; Xie-Brustolin, J.; Damier, P.; Durif, F.; Azulay, J.P.; Derost, P.; Witjas, T.; Raoul, S.; Le Bars, D.; et al. Globus pallidus stimulation reduces frontal hyperactivity in tardive dystonia. J. Cereb. Blood Flow Metab 2008, 28, 1127–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bostan, A.C.; Strick, P.L. The basal ganglia and the cerebellum: Nodes in an integrated network. Nat. Rev. Neurosci. 2018, 19, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Mazere, J.; Dilharreguy, B.; Catheline, G.; Vidailhet, M.; Deffains, M.; Vimont, D.; Ribot, B.; Barse, E.; Cif, L.; Mazoyer, B.; et al. Striatal and cerebellar vesicular acetylcholine transporter expression is disrupted in human DYT1 dystonia. Brain 2021, 144, 909–923. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.; Porcacchia, P.; Ponzo, V.; Carrillo, F.; Caceres-Redondo, M.T.; Brusa, L.; Desiato, M.T.; Arciprete, F.; Di Lorenzo, F.; Pisani, A.; et al. Effects of two weeks of cerebellar theta burst stimulation in cervical dystonia patients. Brain Stimul. 2014, 7, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Brighina, F.; Romano, M.; Giglia, G.; Saia, V.; Puma, A.; Giglia, F.; Fierro, B. Effects of cerebellar TMS on motor cortex of patients with focal dystonia: A preliminary report. Exp. Brain Res. 2009, 192, 651–656. [Google Scholar] [CrossRef]
- Bradnam, L.V.; McDonnell, M.N.; Ridding, M.C. Cerebellar intermittent theta-burst stimulation and motor control training in individuals with cervical dystonia. Brain Sci. 2016, 6, 56. [Google Scholar] [CrossRef] [Green Version]
- Hoffland, B.S.; Kassavetis, P.; Bologna, M.; Teo, J.T.; Bhatia, K.P.; Rothwell, J.C.; Edwards, M.J.; van de Warrenburg, B.P. Cerebellum-dependent associative learning deficits in primary dystonia are normalized by rTMS and practice. Eur. J. Neurosci. 2013, 38, 2166–2171. [Google Scholar] [CrossRef] [PubMed]
- Bradnam, L.V.; Frasca, J.; Kimberley, T.J. Direct current stimulation of primary motor cortex and cerebellum and botulinum toxin a injections in a person with cervical dystonia. Brain Stimul. 2014, 7, 909–911. [Google Scholar] [CrossRef] [PubMed]
- Bradnam, L.V.; Graetz, L.J.; McDonnell, M.N.; Ridding, M.C. Anodal transcranial direct current stimulation to the cerebellum improves handwriting and cyclic drawing kinematics in focal hand dystonia. Front. Hum. Neurosci. 2015, 9, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadnicka, A.; Hamada, M.; Bhatia, K.P.; Rothwell, J.C.; Edwards, M.J. Cerebellar stimulation fails to modulate motor cortex plasticity in writing dystonia. Mov. Disord. 2014, 29, 1304–1307. [Google Scholar] [CrossRef] [PubMed]
- Cooper, I.S. Dystonia: Slrgical approaches to treatment and physiologic implications. Res. Publ. Assoc. Res. Nerv. Ment. Dis. 1976, 55, 369–383. [Google Scholar]
- Cooper, I.S. 20-year followup study of the neurosurgical treatment of dystonia musculorum deformans. Adv. Neurol. 1976, 14, 423–452. [Google Scholar]
- Cooper, I.S.; Riklan, M.; Amin, I.; Waltz, J.M.; Cullinan, T. Chronic cerebellar stimulation in cerebral palsy. Neurology 1976, 26, 744–753. [Google Scholar] [CrossRef]
- Penn, R.D.; Gottlieb, G.L.; Agarwal, G.C. Cerebellar stimulation in man. Quantitative changes in spasticity. J. Neurosurg. 1978, 48, 779–786. [Google Scholar] [CrossRef]
- Galanda, M.; Hovath, S. Different effect of chronic electrical stimulation of the region of the superior cerebellar peduncle and the nucleus ventralis intermedius of the thalamus in the treatment of movement disorders. Stereotact. Funct. Neurosurg. 1997, 69, 116–120. [Google Scholar] [CrossRef]
- Galanda, M.; Mistina, L.; Zoltan, O. Behavioural responses to cerebellar stimulation in cerebral palsy. Acta Neurochir. Suppl. 1989, 46, 37–38. [Google Scholar]
- Galanda, M.; Zoltan, O. Motor and psychological responses to deep cerebellar stimulation in cerebral palsy (correlation with organization of cerebellum into zones). Acta Neurochir. Suppl. 1987, 39, 129–131. [Google Scholar]
- Schulman, J.H.; Davis, R.; Nanes, M. Cerebellar stimulation for spastic cerebral palsy: Preliminary report; on-going double blind study. Pacing Clin. Electrophysiol. 1987, 10, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.; Schulman, J.; Delehanty, A. Cerebellar stimulation for cerebral palsy--double blind study. Acta Neurochir. Suppl. 1987, 39, 126–128. [Google Scholar] [PubMed]
- Davis, R.; Gray, E.; Ryan, T.; Schulman, J. Bioengineering changes in spastic cerebral palsy groups following cerebellar stimulation. Appl. Neurophysiol. 1985, 48, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.; Cullen, R.F., Jr.; Flitter, M.A.; Duenas, D.; Engle, H.; Papazian, O.; Weis, B. Control of spasticity and involuntary movements--cerebellar stimulation. Appl. Neurophysiol. 1977, 40, 135–140. [Google Scholar] [CrossRef]
- Davis, R.; Barolat-Romana, G.; Engle, H. Chronic cerebellar stimulation for cerebral palsy--five-year study. Acta Neurochir. Suppl. 1980, 30, 317–332. [Google Scholar]
- Davis, R. Cerebellar stimulation for cerebral palsy spasticity, function, and seizures. Arch. Med. Res. 2000, 31, 290–299. [Google Scholar] [CrossRef]
- Galanda, M.; Horvath, S. Effect of stereotactic high-frequency stimulation in the anterior lobe of the cerebellum in cerebral palsy: A new suboccipital approach. Stereotact. Funct. Neurosurg. 2003, 80, 102–107. [Google Scholar] [CrossRef]
- Galanda, M.; Horvath, S. Stereotactic stimulation of the anterior lobe of the cerebellum in cerebral palsy from a suboccipital approach. Acta Neurochir. Suppl. 2007, 97, 239–243. [Google Scholar]
- Harat, M.; Radziszewski, K.; Rudas, M.; Okon, M.; Galanda, M. Clinical evaluation of deep cerebellar stimulation for spasticity in patients with cerebral palsy. Neurol. Neurochir. Pol. 2009, 43, 36–44. [Google Scholar]
- Cooper, I.S. Effect of chronic stimulation of anterior cerebellum on neurological disease. Lancet 1973, 1, 206. [Google Scholar] [CrossRef]
- Rosenow, J.; Das, K.; Rovit, R.L.; Couldwell, W.T. Irving s. Cooper and his role in intracranial stimulation for movement disorders and epilepsy. Stereotact. Funct. Neurosurg. 2002, 78, 95–112. [Google Scholar] [CrossRef]
- Vidailhet, M.; Vercueil, L.; Houeto, J.L.; Krystkowiak, P.; Benabid, A.L.; Cornu, P.; Lagrange, C.; Tezenas du Montcel, S.; Dormont, D.; Grand, S.; et al. Bilateral deep-brain stimulation of the globus pallidus in primary generalized dystonia. N. Engl. J. Med. 2005, 352, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Vidailhet, M.; Vercueil, L.; Houeto, J.L.; Krystkowiak, P.; Lagrange, C.; Yelnik, J.; Bardinet, E.; Benabid, A.L.; Navarro, S.; Dormont, D.; et al. Bilateral, pallidal, deep-brain stimulation in primary generalised dystonia: A prospective 3 year follow-up study. Lancet Neurol. 2007, 6, 223–229. [Google Scholar] [CrossRef]
- Volkmann, J.; Wolters, A.; Kupsch, A.; Muller, J.; Kuhn, A.A.; Schneider, G.H.; Poewe, W.; Hering, S.; Eisner, W.; Muller, J.U.; et al. Pallidal deep brain stimulation in patients with primary generalised or segmental dystonia: 5-year follow-up of a randomised trial. Lancet Neurol. 2012, 11, 1029–1038. [Google Scholar] [CrossRef]
- Volkmann, J.; Mueller, J.; Deuschl, G.; Kuhn, A.A.; Krauss, J.K.; Poewe, W.; Timmermann, L.; Falk, D.; Kupsch, A.; Kivi, A.; et al. Pallidal neurostimulation in patients with medication-refractory cervical dystonia: A randomised, sham-controlled trial. Lancet Neurol. 2014, 13, 875–884. [Google Scholar] [CrossRef]
- Fukaya, C.; Katayama, Y.; Kano, T.; Nagaoka, T.; Kobayashi, K.; Oshima, H.; Yamamoto, T. Thalamic deep brain stimulation for writer’s cramp. J. Neurosurg. 2007, 107, 977–982. [Google Scholar] [CrossRef]
- Goto, S.; Shimazu, H.; Matsuzaki, K.; Tamura, T.; Murase, N.; Nagahiro, S.; Kaji, R. Thalamic Vo-complex vs pallidal deep brain stimulation for focal hand dystonia. Neurology 2008, 70, 1500–1501. [Google Scholar] [CrossRef]
- Goto, S.; Tsuiki, H.; Soyama, N.; Okamura, A.; Yamada, K.; Yoshikawa, M.; Hashimoto, Y.; Ushio, Y. Stereotactic selective Vo-complex thalamotomy in a patient with dystonic writer’s cramp. Neurology 1997, 49, 1173–1174. [Google Scholar] [CrossRef]
- Mure, H.; Morigaki, R.; Koizumi, H.; Okita, S.; Kawarai, T.; Miyamoto, R.; Kaji, R.; Nagahiro, S.; Goto, S. Deep brain stimulation of the thalamic ventral lateral anterior nucleus for DYT6 dystonia. Stereotact. Funct. Neurosurg. 2014, 92, 393–396. [Google Scholar] [CrossRef]
- Horisawa, S.; Ochiai, T.; Goto, S.; Nakajima, T.; Takeda, N.; Fukui, A.; Hanada, T.; Kawamata, T.; Taira, T. Safety and long-term efficacy of ventro-oral thalamotomy for focal hand dystonia: A retrospective study of 171 patients. Neurology 2019, 92, e371–e377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horisawa, S.; Taira, T.; Goto, S.; Ochiai, T.; Nakajima, T. Long-term improvement of musician’s dystonia after stereotactic ventro-oral thalamotomy. Ann. Neurol. 2013, 74, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Horisawa, S.; Goto, S.; Nakajima, T.; Ochiai, T.; Kawamata, T.; Taira, T. Stereotactic thalamotomy for hairdresser’s dystonia: A case series. Stereotact. Funct. Neurosurg. 2016, 94, 201–206. [Google Scholar] [CrossRef]
- Horisawa, S.; Goto, S.; Nakajima, T.; Kawamata, T.; Taira, T. Bilateral stereotactic thalamotomy for bilateral musician’s hand dystonia. World Neurosurg. 2016, 92, 585.e21–585.e25. [Google Scholar] [CrossRef]
- Shimizu, T.; Maruo, T.; Miura, S.; Kishima, H.; Ushio, Y.; Goto, S. Stereotactic lesioning of the thalamic Vo nucleus for the treatment of writer’s cramp (focal hand dystonia). Front. Neurol. 2018, 9, 1008. [Google Scholar] [CrossRef]
- Morishita, T.; Foote, K.D.; Haq, I.U.; Zeilman, P.; Jacobson, C.E.; Okun, M.S. Should we consider Vim thalamic deep brain stimulation for select cases of severe refractory dystonic tremor. Stereotact. Funct. Neurosurg. 2010, 88, 98–104. [Google Scholar] [CrossRef]
- Hedera, P.; Phibbs, F.T.; Dolhun, R.; Charles, P.D.; Konrad, P.E.; Neimat, J.S.; Davis, T.L. Surgical targets for dystonic tremor: Considerations between the globus pallidus and ventral intermediate thalamic nucleus. Parkinsonism Relat. Disord. 2013, 19, 684–686. [Google Scholar] [CrossRef]
- Morigaki, R.; Nagahiro, S.; Kaji, R.; Goto, S. Current use of thalamic surgeries for treating movement disorders. In Thalamus: Anatomy, Functions and Disorders; Song, J.L., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2011; pp. 1–31. [Google Scholar]
- Lin, S.; Zhang, C.; Li, H.; Wang, Y.; Wu, Y.; Wang, T.; Pan, Y.; Sun, B.; Wu, Y.; Li, D. High frequency deep brain stimulation of superior cerebellar peduncles in a patient with cerebral palsy. Tremor. Other Hyperkinet. Mov. 2020, 10, 38. [Google Scholar] [CrossRef]
- Nicholson, C.L.; Coubes, P.; Poulen, G. Dentate nucleus as target for deep brain stimulation in dystono-dyskinetic syndromes. Neurochirurgie 2020, 66, 258–265. [Google Scholar] [CrossRef]
- Horisawa, S.; Kohara, K.; Nonaka, T.; Mochizuki, T.; Kawamata, T.; Taira, T. Case report: Deep cerebellar stimulation for tremor and dystonia. Front. Neurol. 2021, 12, 642904. [Google Scholar] [CrossRef] [PubMed]
- Horisawa, S.; Arai, T.; Suzuki, N.; Kawamata, T.; Taira, T. The striking effects of deep cerebellar stimulation on generalized fixed dystonia: Case report. J. Neurosurg. 2019, 132, 712–716. [Google Scholar] [CrossRef]
- Sokal, P.; Rudas, M.; Harat, M.; Szylberg, L.; Zielinski, P. Deep anterior cerebellar stimulation reduces symptoms of secondary dystonia in patients with cerebral palsy treated due to spasticity. Clin. Neurol. Neurosurg. 2015, 135, 62–68. [Google Scholar] [CrossRef]
- Elia, A.E.; Bagella, C.F.; Ferre, F.; Zorzi, G.; Calandrella, D.; Romito, L.M. Deep brain stimulation for dystonia due to cerebral palsy: A review. Eur. J. Paediatr. Neurol. 2018, 22, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.G.; Bledsoe, I.O.; Luthra, N.S.; Miocinovic, S.; Starr, P.A.; Ostrem, J.L. Cerebellar deep brain stimulation for acquired hemidystonia. Mov. Disord. Clin. Pr. 2020, 7, 188–193. [Google Scholar] [CrossRef]
- Lin, S.; Wu, Y.; Li, H.; Zhang, C.; Wang, T.; Pan, Y.; He, L.; Shen, R.; Deng, Z.; Sun, B.; et al. Deep brain stimulation of the globus pallidus internus versus the subthalamic nucleus in isolated dystonia. J. Neurosurg. 2019, 132, 721–732. [Google Scholar] [CrossRef]
- Macerollo, A.; Sajin, V.; Bonello, M.; Barghava, D.; Alusi, S.H.; Eldridge, P.R.; Osman-Farah, J. Deep brain stimulation in dystonia: State of art and future directions. J. Neurosci. Methods 2020, 340, 108750. [Google Scholar] [CrossRef]
- Wu, Y.S.; Ni, L.H.; Fan, R.M.; Yao, M.Y. Meta-regression analysis of the long-term effects of pallidal and subthalamic deep brain stimulation for the treatment of isolated dystonia. World Neurosurg. 2019, 129, e409–e416. [Google Scholar] [CrossRef]
- Wagle Shukla, A.; Ostrem, J.L.; Vaillancourt, D.E.; Chen, R.; Foote, K.D.; Okun, M.S. Physiological effects of subthalamic nucleus deep brain stimulation surgery in cervical dystonia. J. Neurol. Neurosurg. Psychiatry 2018, 89, 1296–1300. [Google Scholar] [CrossRef]
- Toda, H.; Saiki, H.; Nishida, N.; Iwasaki, K. Update on deep brain stimulation for dyskinesia and dystonia: A literature review. Neurol. Med. Chir. 2016, 56, 236–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaias, I.U.; Alterman, R.L.; Tagliati, M. Outcome predictors of pallidal stimulation in patients with primary dystonia: The role of disease duration. Brain 2008, 131, 1895–1902. [Google Scholar] [CrossRef] [Green Version]
- Yianni, J.; Bain, P.G.; Gregory, R.P.; Nandi, D.; Joint, C.; Scott, R.B.; Stein, J.F.; Aziz, T.Z. Post-operative progress of dystonia patients following globus pallidus internus deep brain stimulation. Eur. J. Neurol. 2003, 10, 239–247. [Google Scholar] [CrossRef]
- Krauss, J.K.; Yianni, J.; Loher, T.J.; Aziz, T.Z. Deep brain stimulation for dystonia. J. Clin. Neurophysiol. 2004, 21, 18–30. [Google Scholar] [CrossRef]
- Yokochi, F.; Kato, K.; Iwamuro, H.; Kamiyama, T.; Kimura, K.; Yugeta, A.; Okiyama, R.; Taniguchi, M.; Kumada, S.; Ushiba, J. Resting-state pallidal-cortical oscillatory couplings in patients with predominant phasic and tonic dystonia. Front. Neurol. 2018, 9, 375. [Google Scholar] [CrossRef]
- Kupsch, A.; Benecke, R.; Muller, J.; Trottenberg, T.; Schneider, G.H.; Poewe, W.; Eisner, W.; Wolters, A.; Muller, J.U.; Deuschl, G.; et al. Pallidal deep-brain stimulation in primary generalized or segmental dystonia. N. Engl. J. Med. 2006, 355, 1978–1990. [Google Scholar] [CrossRef] [Green Version]
- Volkmann, J.; Benecke, R. Deep brain stimulation for dystonia: Patient selection and evaluation. Mov. Disord. 2002, 17 (Suppl. 3), S112–S115. [Google Scholar] [CrossRef] [PubMed]
- Hung, S.W.; Hamani, C.; Lozano, A.M.; Poon, Y.Y.; Piboolnurak, P.; Miyasaki, J.M.; Lang, A.E.; Dostrovsky, J.O.; Hutchison, W.D.; Moro, E. Long-term outcome of bilateral pallidal deep brain stimulation for primary cervical dystonia. Neurology 2007, 68, 457–459. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, X.; Yianni, J.; Green, A.L.; Joint, C.; Stein, J.F.; Bain, P.G.; Gregory, R.; Aziz, T.Z. Use of surface electromyography to assess and select patients with idiopathic dystonia for bilateral pallidal stimulation. J. Neurosurg. 2006, 105, 21–25. [Google Scholar] [CrossRef]
- Liu, X.; Griffin, I.C.; Parkin, S.G.; Miall, R.C.; Rowe, J.G.; Gregory, R.P.; Scott, R.B.; Aziz, T.Z.; Stein, J.F. Involvement of the medial pallidum in focal myoclonic dystonia: A clinical and neurophysiological case study. Mov. Disord. 2002, 17, 346–353. [Google Scholar] [CrossRef]
- Liu, X.; Wang, S.; Yianni, J.; Nandi, D.; Bain, P.G.; Gregory, R.; Stein, J.F.; Aziz, T.Z. The sensory and motor representation of synchronized oscillations in the globus pallidus in patients with primary dystonia. Brain 2008, 131, 1562–1573. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yianni, J.; Wang, S.; Bain, P.G.; Stein, J.F.; Aziz, T.Z. Different mechanisms may generate sustained hypertonic and rhythmic bursting muscle activity in idiopathic dystonia. Exp. Neurol. 2006, 198, 204–213. [Google Scholar] [CrossRef]
- Chen, C.C.; Kuhn, A.A.; Hoffmann, K.T.; Kupsch, A.; Schneider, G.H.; Trottenberg, T.; Krauss, J.K.; Wohrle, J.C.; Bardinet, E.; Yelnik, J.; et al. Oscillatory pallidal local field potential activity correlates with involuntary EMG in dystonia. Neurology 2006, 66, 418–420. [Google Scholar] [CrossRef]
- Chen, C.C.; Kuhn, A.A.; Trottenberg, T.; Kupsch, A.; Schneider, G.H.; Brown, P. Neuronal activity in globus pallidus interna can be synchronized to local field potential activity over 3–12 Hz in patients with dystonia. Exp. Neurol. 2006, 202, 480–486. [Google Scholar] [CrossRef]
- Silberstein, P.; Kuhn, A.A.; Kupsch, A.; Trottenberg, T.; Krauss, J.K.; Wohrle, J.C.; Mazzone, P.; Insola, A.; Di Lazzaro, V.; Oliviero, A.; et al. Patterning of globus pallidus local field potentials differs between parkinson’s disease and dystonia. Brain 2003, 126, 2597–2608. [Google Scholar] [CrossRef]
- Weinberger, M.; Hutchison, W.D.; Alavi, M.; Hodaie, M.; Lozano, A.M.; Moro, E.; Dostrovsky, J.O. Oscillatory activity in the globus pallidus internus: Comparison between parkinson’s disease and dystonia. Clin. Neurophysiol. 2012, 123, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Barow, E.; Neumann, W.J.; Brucke, C.; Huebl, J.; Horn, A.; Brown, P.; Krauss, J.K.; Schneider, G.H.; Kuhn, A.A. Deep brain stimulation suppresses pallidal low frequency activity in patients with phasic dystonic movements. Brain 2014, 137, 3012–3024. [Google Scholar] [CrossRef]
- Groen, J.L.; Ritz, K.; Contarino, M.F.; van de Warrenburg, B.P.; Aramideh, M.; Foncke, E.M.; van Hilten, J.J.; Schuurman, P.R.; Speelman, J.D.; Koelman, J.H.; et al. Dyt6 dystonia: Mutation screening, phenotype, and response to deep brain stimulation. Mov. Disord. 2010, 25, 2420–2427. [Google Scholar] [CrossRef]
- Panov, F.; Tagliati, M.; Ozelius, L.J.; Fuchs, T.; Gologorsky, Y.; Cheung, T.; Avshalumov, M.; Bressman, S.B.; Saunders-Pullman, R.; Weisz, D.; et al. Pallidal deep brain stimulation for DYT6 dystonia. J. Neurol. Neurosurg. Psychiatry 2012, 83, 182–187. [Google Scholar] [CrossRef]
- Zittel, S.; Moll, C.K.; Bruggemann, N.; Tadic, V.; Hamel, W.; Kasten, M.; Lohmann, K.; Lohnau, T.; Winkler, S.; Gerloff, C.; et al. Clinical neuroimaging and electrophysiological assessment of three DYT6 dystonia families. Mov. Disord. 2010, 25, 2405–2412. [Google Scholar] [CrossRef]
- Quartarone, A.; Hallett, M. Emerging concepts in the physiological basis of dystonia. Mov. Disord. 2013, 28, 958–967. [Google Scholar] [CrossRef]
- Benabid, A.L.; Pollak, P.; Gao, D.; Hoffmann, D.; Limousin, P.; Gay, E.; Payen, I.; Benazzouz, A. Chronic electrical stimulation of the ventralis intermedius nucleus of the thalamus as a treatment of movement disorders. J. Neurosurg. 1996, 84, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Tasker, R.R. Deep brain stimulation is preferable to thalamotomy for tremor suppression. Surg. Neurol. 1998, 49, 145–153. [Google Scholar] [CrossRef]
- Pilitsis, J.G.; Metman, L.V.; Toleikis, J.R.; Hughes, L.E.; Sani, S.B.; Bakay, R.A. Factors involved in long-term efficacy of deep brain stimulation of the thalamus for essential tremor. J. Neurosurg. 2008, 109, 640–646. [Google Scholar] [CrossRef]
- Morigaki, R.; Goto, S. Deep brain stimulation for essential tremor. In Deep Brain Stimulation for Neurological Disorders; Itakura, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 135–155. [Google Scholar]
- Breakefield, X.O.; Blood, A.J.; Li, Y.; Hallett, M.; Hanson, P.I.; Standaert, D.G. The pathophysiological basis of dystonias. Nat. Rev. Neurosci. 2008, 9, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Morigaki, R.; Goto, S. Striatal vulnerability in huntington’s disease: Neuroprotection versus neurotoxicity. Brain Sci. 2017, 7, 63. [Google Scholar] [CrossRef]
- Kawarai, T.; Morigaki, R.; Kaji, R.; Goto, S. Clinicopathological phenotype and genetics of x-linked dystonia-parkinsonism (XDP; DYT3; lubag). Brain Sci. 2017, 7, 72. [Google Scholar] [CrossRef]
- Goto, S.; Lee, L.V.; Munoz, E.L.; Tooyama, I.; Tamiya, G.; Makino, S.; Ando, S.; Dantes, M.B.; Yamada, K.; Matsumoto, S.; et al. Functional anatomy of the basal ganglia in X-linked recessive dystonia-parkinsonism. Ann. Neurol. 2005, 58, 7–17. [Google Scholar] [CrossRef]
- Goto, S.; Nagahiro, S.; Kaji, R. Striosome-matrix pathology of dystonias: A new hypothesis for dystonia genesis. In Dystonia: Causes, Symptoms, and Treatment; Kurstot, J., Forsström, M., Eds.; Nova Science: New York, NY, USA, 2010; pp. 1–22. [Google Scholar]
- Shakkottai, V.G.; Batla, A.; Bhatia, K.; Dauer, W.T.; Dresel, C.; Niethammer, M.; Eidelberg, D.; Raike, R.S.; Smith, Y.; Jinnah, H.A.; et al. Current opinions and areas of consensus on the role of the cerebellum in dystonia. Cerebellum 2017, 16, 577–594. [Google Scholar] [CrossRef] [Green Version]
- Morigaki, R.; Okita, S.; Goto, S. Dopamine-induced changes in Galphaolf protein levels in striatonigral and striatopallidal medium spiny neurons underlie the genesis of l-dopa-induced dyskinesia in parkinsonian mice. Front. Cell Neurosci. 2017, 11, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assous, M.; Tepper, J.M. Excitatory extrinsic afferents to striatal interneurons and interactions with striatal microcircuitry. Eur. J. Neurosci. 2019, 49, 593–603. [Google Scholar] [CrossRef]
- Koos, T.; Tepper, J.M. Inhibitory control of neostriatal projection neurons by GABAergic interneurons. Nat. Neurosci. 1999, 2, 467–472. [Google Scholar] [CrossRef]
- Mink, J.W. The basal ganglia: Focused selection and inhibition of competing motor programs. Prog. Neurobiol. 1996, 50, 381–425. [Google Scholar] [CrossRef]
- Amemori, K.; Gibb, L.G.; Graybiel, A.M. Shifting responsibly: The importance of striatal modularity to reinforcement learning in uncertain environments. Front. Hum. Neurosci. 2011, 5, 47. [Google Scholar] [CrossRef] [Green Version]
- Pisani, A.; Martella, G.; Tscherter, A.; Bonsi, P.; Sharma, N.; Bernardi, G.; Standaert, D.G. Altered responses to dopaminergic D2 receptor activation and N-type calcium currents in striatal cholinergic interneurons in a mouse model of DYT1 dystonia. Neurobiol. Dis. 2006, 24, 318–325. [Google Scholar] [CrossRef]
- Martella, G.; Tassone, A.; Sciamanna, G.; Platania, P.; Cuomo, D.; Viscomi, M.T.; Bonsi, P.; Cacci, E.; Biagioni, S.; Usiello, A.; et al. Impairment of bidirectional synaptic plasticity in the striatum of a mouse model of DYT1 dystonia: Role of endogenous acetylcholine. Brain 2009, 132, 2336–2349. [Google Scholar] [CrossRef] [PubMed]
- Bonsi, P.; Cuomo, D.; Martella, G.; Madeo, G.; Schirinzi, T.; Puglisi, F.; Ponterio, G.; Pisani, A. Centrality of striatal cholinergic transmission in basal ganglia function. Front. Neuroanat. 2011, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Bruggemann, N. Contemporary functional neuroanatomy and pathophysiology of dystonia. J. Neural Transm. 2021, 128, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Smith, Y.; Raju, D.V.; Pare, J.F.; Sidibe, M. The thalamostriatal system: A highly specific network of the basal ganglia circuitry. Trends Neurosci. 2004, 27, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Sidibe, M.; Smith, Y. Thalamic inputs to striatal interneurons in monkeys: Synaptic organization and co-localization of calcium binding proteins. Neuroscience 1999, 89, 1189–1208. [Google Scholar] [CrossRef]
- Rudkin, T.M.; Sadikot, A.F. Thalamic input to parvalbumin-immunoreactive GABAergic interneurons: Organization in normal striatum and effect of neonatal decortication. Neuroscience 1999, 88, 1165–1175. [Google Scholar] [CrossRef]
- Bennay, M.; Gernert, M.; Richter, A. Spontaneous remission of paroxysmal dystonia coincides with normalization of entopeduncular activity in dt(SZ) mutants. J. Neurosci. 2001, 21, RC153. [Google Scholar] [CrossRef]
- Gernert, M.; Richter, A.; Loscher, W. In vivo extracellular electrophysiology of pallidal neurons in dystonic and nondystonic hamsters. J. Neurosci. Res. 1999, 57, 894–905. [Google Scholar] [CrossRef]
- Gernert, M.; Richter, A.; Loscher, W. Alterations in spontaneous single unit activity of striatal subdivisions during ontogenesis in mutant dystonic hamsters. Brain Res. 1999, 821, 277–285. [Google Scholar] [CrossRef]
- Gernert, M.; Hamann, M.; Bennay, M.; Loscher, W.; Richter, A. Deficit of striatal parvalbumin-reactive GABAergic interneurons and decreased basal ganglia output in a genetic rodent model of idiopathic paroxysmal dystonia. J. Neurosci. 2000, 20, 7052–7058. [Google Scholar] [CrossRef]
- Gittis, A.H.; Leventhal, D.K.; Fensterheim, B.A.; Pettibone, J.R.; Berke, J.D.; Kreitzer, A.C. Selective inhibition of striatal fast-spiking interneurons causes dyskinesias. J. Neurosci. 2011, 31, 15727–15731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Threlfell, S.; Lalic, T.; Platt, N.J.; Jennings, K.A.; Deisseroth, K.; Cragg, S.J. Striatal dopamine release is triggered by synchronized activity in cholinergic interneurons. Neuron 2012, 75, 58–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crittenden, J.R.; Tillberg, P.W.; Riad, M.H.; Shima, Y.; Gerfen, C.R.; Curry, J.; Housman, D.E.; Nelson, S.B.; Boyden, E.S.; Graybiel, A.M. Striosome-dendron bouquets highlight a unique striatonigral circuit targeting dopamine-containing neurons. Proc. Natl. Acad. Sci. USA 2016, 113, 11318–11323. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morigaki, R.; Miyamoto, R.; Matsuda, T.; Miyake, K.; Yamamoto, N.; Takagi, Y. Dystonia and Cerebellum: From Bench to Bedside. Life 2021, 11, 776. https://doi.org/10.3390/life11080776
Morigaki R, Miyamoto R, Matsuda T, Miyake K, Yamamoto N, Takagi Y. Dystonia and Cerebellum: From Bench to Bedside. Life. 2021; 11(8):776. https://doi.org/10.3390/life11080776
Chicago/Turabian StyleMorigaki, Ryoma, Ryosuke Miyamoto, Taku Matsuda, Kazuhisa Miyake, Nobuaki Yamamoto, and Yasushi Takagi. 2021. "Dystonia and Cerebellum: From Bench to Bedside" Life 11, no. 8: 776. https://doi.org/10.3390/life11080776
APA StyleMorigaki, R., Miyamoto, R., Matsuda, T., Miyake, K., Yamamoto, N., & Takagi, Y. (2021). Dystonia and Cerebellum: From Bench to Bedside. Life, 11(8), 776. https://doi.org/10.3390/life11080776