
Experimental Approaches for Testing the Hypothesis of the Emergence of Life at Submarine Alkaline Vents


Abstract
:1. Introduction
1.1. Context of Origins of Life Studies and Bases of the AHV Model
2. The Conceptual Background for the EoL in Alkaline Hydrothermal Systems
2.1. Overall Geochemical Context in the AHV Model
2.2. Concepts from Complexity Sciences and Far-From-Equilibrium Thermodynamics in the AHV Model
2.3. The Early Chemiosmotic Coupling and the Geoelectrochemical Driven EoL
3. Experimental Setups and Results Investigating the AHV Model
3.1. Reactors Simulating AHV Chemistry and Geochemistry
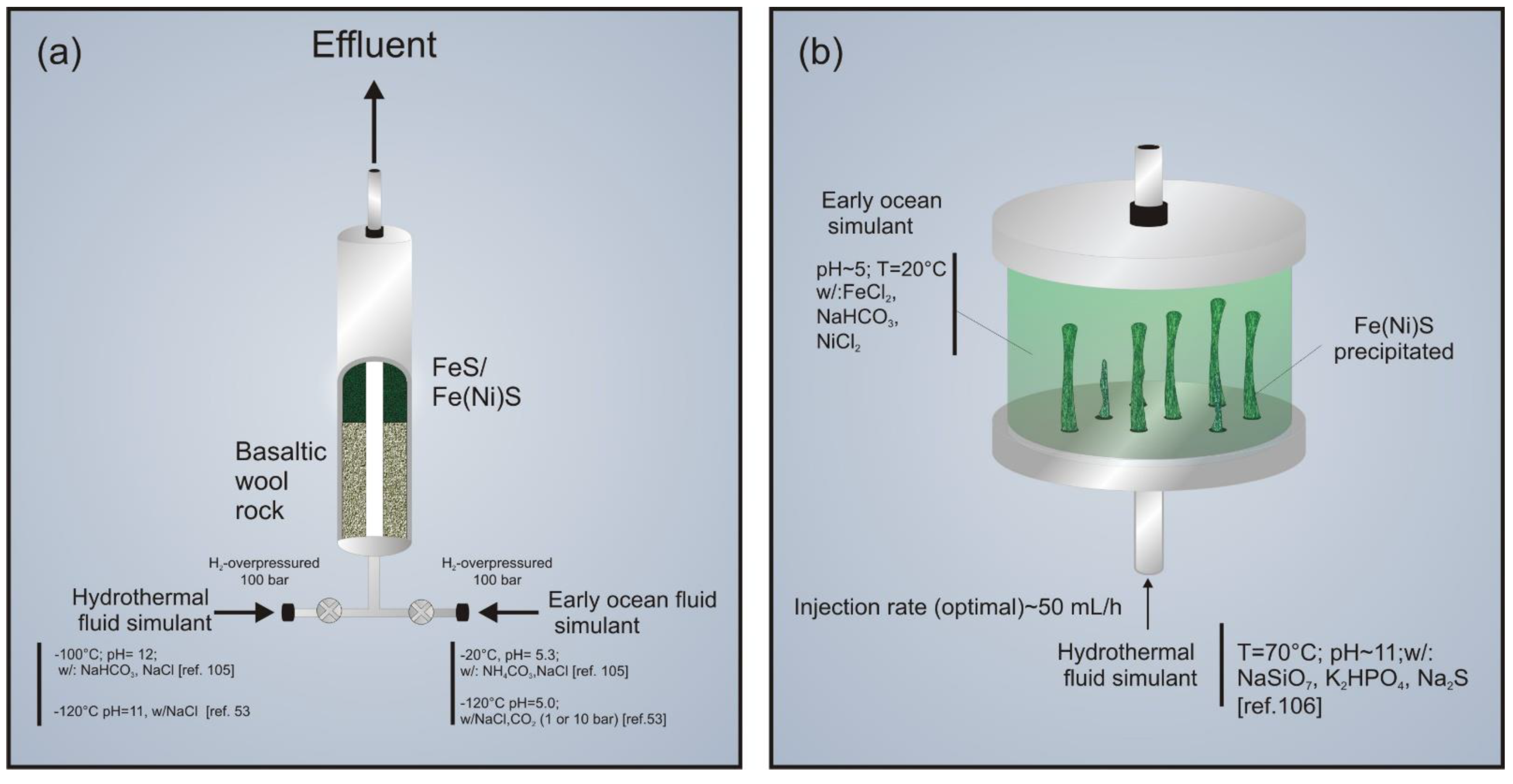
3.1.1. Hydrothermal High Pressure Flow Reactors
3.1.2. The Origin-Of-Life Reactor
3.2. Interfacial Chemistry and Electrochemistry as Key Factors for the Emergence of Life
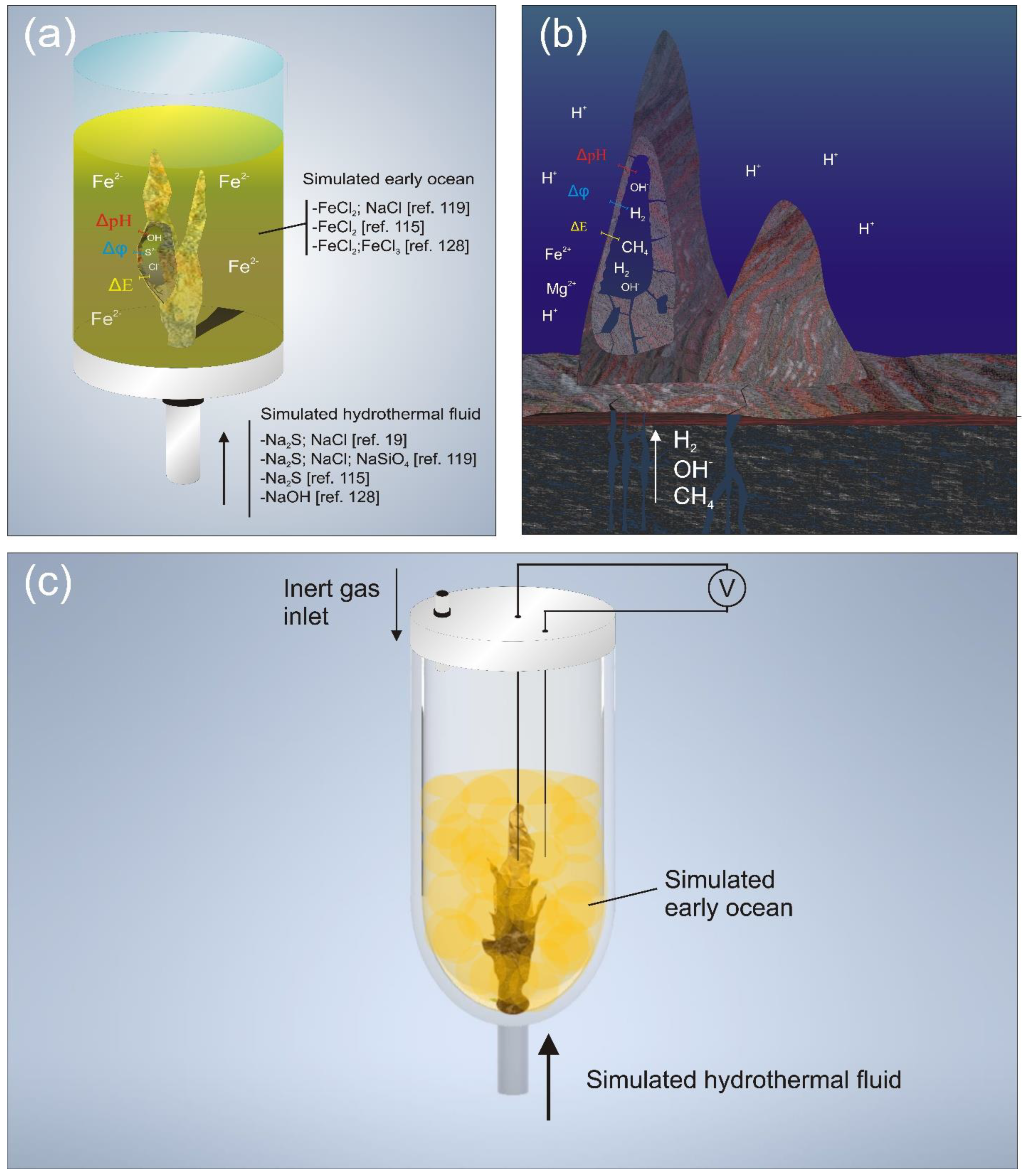
3.2.1. Chemical Gardens and the Simulation of Early AHVs
3.2.2. Electrochemically Activated Mineral Surfaces
3.2.3. Microfluidic Scale Setup Simulating the Hydrothermal Vent-Ocean Interface
4. Overall Perspectives and Trends for the Model
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lazcano, A. Historical development of origins research. Cold Spring Harb. Perspect. Biol. 2010, 2, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazen, R.M. Emergence and the experimental pursuit of the origin of life. In Exploring the Origin, Extent, and Future of Life; Bertka, C.M., Ed.; Cambridge University Press: Cambridge, UK, 2009; pp. 21–46. [Google Scholar]
- Domagal-Goldman, S.D.; Wright, K.E.; Adamala, K.; De La Rubia, L.A.; Bond, J.; Dartnell, L.R.; Goldman, A.D.; Lynch, K.; Naud, M.-E.E.; Paulino-Lima, I.G.; et al. The astrobiology primer v2.0. Astrobiology 2016, 16, 561–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Mirazo, K.; Briones, C.; De La Escosura, A. Prebiotic systems chemistry: New perspectives for the origins of life. Chem. Rev. 2014, 114, 285–366. [Google Scholar] [CrossRef]
- Branscomb, E.; Biancalani, T.; Goldenfeld, N.; Russell, M. Escapement mechanisms and the conversion of disequilibria; the engines of creation. Phys. Rep. 2017, 677, 1–60. [Google Scholar] [CrossRef]
- Gargaud, M.; Martin, H.; López-García, P.; Montmerle, T.; Pascal, R. Young Sun, Early Earth and the Origins of Life; Springer: Berlin/Heidelberg, Germany, 2012; ISBN 978-3-642-22551-2. [Google Scholar]
- Kauffman, S.A. The Origins of Order: Self-Organization and Selection in Evolution; Oxford University Press: New York, NY, USA, 1993; ISBN 0195058119. [Google Scholar]
- Orgel, L.E. The origin of life—A review of facts and speculations. Trends Biochem. Sci. 1998, 23, 491–495. [Google Scholar] [CrossRef]
- Miller, S.L.; Urey, H.C. Organic Compound Synthesis on the Primitive Earth. Science 1959, 130, 245–251. [Google Scholar] [CrossRef]
- Russell, M.J.; Barge, L.M.; Bhartia, R.; Bocanegra, D.; Bracher, P.J.; Branscomb, E.; Kidd, R.; McGlynn, S.; Meier, D.H.; Nitschke, W.; et al. The drive to life on wet and Icy Worlds. Astrobiology 2014, 14, 308–343. [Google Scholar] [CrossRef]
- Silantyev, S.A.; Mironenko, M.V.; Novoselov, A.A. Hydrothermal systems in peridotites of slow-spreading mid-oceanic ridges. Modeling phase transitions and material balance: Downwelling limb of a hydrothermal circulation cell. Petrology 2009, 17, 138–157. [Google Scholar] [CrossRef]
- Lowell, R.P.; Rona, P.A. Seafloor hydrothermal systems driven by the serpentinization of peridotite. Geophys. Res. Lett. 2002, 29, 26-1–26-4. [Google Scholar] [CrossRef]
- Von Damm, K.L. Seafloor hydrothermal activity: Black smoker chemistry and chimneys. Annu. Rev. Earth Planet. Sci. 1990, 18, 173–204. [Google Scholar] [CrossRef]
- Janecky, D.R.; Seyfried, W.E. Hydrothermal serpentinization of peridotite within the oceanic crust: Experimental investigations of mineralogy and major element chemistry. Geochim. Cosmochim. Acta 1986, 50, 1357–1378. [Google Scholar] [CrossRef]
- Kelley, D.S.; Karson, J.A.; Blackman, D.K.; Früh-Green, G.L.; Butterfield, D.A.; Lilley, M.D.; Olson, E.J.; Schrenk, M.O.; Roe, K.K.; Lebon, G.T.; et al. An off-axis hydrothermal vent field near the mid-atlantic ridge at 30° n. Nature 2001, 412, 145–149. [Google Scholar] [CrossRef]
- Kelley, D.S.; Karson, J.A.; Früh-Green, G.L.; Yoerger, D.R.; Shank, T.M.; Butterfield, D.A.; Hayes, J.M.; Schrenk, M.O.; Olson, E.J.; Proskurowski, G.; et al. A serpentinite-hosted ecosystem: The Lost City hydrothermal field. Science 2005, 307, 1428–1434. [Google Scholar] [CrossRef]
- Marteinsson, V.T.; Kristjánsson, J.K.; Kristmannsdóttir, H.; Dahlkvist, M.; Sæmundsson, K.; Hannington, M.; Pétursdóttir, S.K.; Geptner, A.; Stoffers, P. Discovery and Description of Giant Submarine Smectite Cones on the Seafloor in Eyjafjordur, Northern Iceland, and a Novel Thermal Microbial Habitat. Appl. Environ. Microbiol. 2001, 67, 827–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brazelton, W.J.; Schrenk, M.O.; Kelley, D.S.; Baross, J.A. Methane- and sulfur-metabolizing microbial communities dominate the lost city hydrothermal field ecosystem. Appl. Environ. Microbiol. 2006, 72, 6257–6270. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.J.; Hall, A.J.; Turner, D. In vitro growth of iron sulphide chimneys: Possible culture chambers for origin-of-life experiments. Terra Nov. 1989, 1, 238–241. [Google Scholar] [CrossRef]
- Russell, M.J.; Arndt, N.T. Geodynamic and metabolic cycles in the Hadean. Biogeosciences 2005, 2, 97–111. [Google Scholar] [CrossRef] [Green Version]
- McNichol, J. Primordial soup, fool’s gold, and spontaneous generation: A brief introduction to the theory, history, and philosophy of the search for the origin of life. Biochem. Mol. Biol. Educ. 2008, 36, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Oparin, A.I. The Origin of Life, 1st ed.; Dover: New York, NY, USA, 1924. [Google Scholar]
- Haldane JBS The orgin of life. Ration. Annu. 1929, 3, 3–10. [CrossRef]
- De Duve, C.; Miller, S.L. Two-dimensional life? Proc. Natl. Acad. Sci. USA 1991, 88, 10014–10017. [Google Scholar] [CrossRef] [Green Version]
- Wächtershäuser, G. Before enzymes and templates: Theory of surface metabolism. Microbiol. Rev. 1988, 52, 452–484. [Google Scholar] [CrossRef]
- Russell, M.J.; Martin, W. The rocky roots of the acetyl-CoA pathway. Trends Biochem. Sci. 2004, 29, 358–363. [Google Scholar] [CrossRef] [Green Version]
- Wächtershäuser, G. Pyrite Formation, the First Energy Source for Life: A Hypothesis. Syst. Appl. Microbiol. 1988, 10, 207–210. [Google Scholar] [CrossRef]
- Wächtershäuser, G. Evolution of the first metabolic cycles (chemoautotrophy/reductive citric acid cycle/origin of life/pyrite). Proc. Natl. Acad. Sci. USA 1990, 87, 200–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, M.J.; Daniel, R.M.; Hall, A.J.; Sherringham, J.A. A hydrothermally precipitated catalytic iron sulphide membrane as a first step toward life. J. Mol. Evol. 1994, 39, 231–243. [Google Scholar] [CrossRef]
- Russell, M.J.; Hall, A.J. The emergence of life from iron monosulphide bubbles at a submarine hydrothermal redox and pH front. J. Geol. Soc. Lond. 1997, 154, 377–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, G.J.T.; Surman, A.J.; McIver, J.; Colón-Santos, S.M.; Gromski, P.S.; Buchwald, S.; Suárez Marina, I.; Cronin, L. Miller-Urey Spark-Discharge Experiments in the Deuterium World. Angew. Chem. 2017, 129, 8191–8194. [Google Scholar] [CrossRef] [Green Version]
- Urey, H.C. On the Early Chemical History of the Earth and the Origin of Life. Proc. Natl. Acad. Sci. USA 1952, 38, 351–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, S.L.; Urey, H.C.; Oró, J. Origin of organic compounds on the primitive earth and in meteorites. J. Mol. Evol. 1976, 9, 59–72. [Google Scholar] [CrossRef]
- Zahnle, K.; Schaefer, L.; Fegley, B. Earth’s Earliest Atmospheres. Cold Spring Harb. Perspect. Biol. 2010, 2, a004895. [Google Scholar] [CrossRef] [Green Version]
- Huber, C.; Wachterhauser, G. Activated Acetic Acid by Carbon Fixation on (Fe, Ni) S Under Primordial Conditions. Science 1997, 276, 245–248. [Google Scholar] [CrossRef] [Green Version]
- Schneider, E.D.; Kay, J.J.; Murphy, M.P.; Neill, L.A.J.O.; The, L.; Fifty, N.; Schneider, E.D.; Kay, J.J. Order from disorder: The thermodynamics of complexity in biology. In What is Life? Next Fifty Years; Cambridge University Press: Cambridge, UK, 2010; Volume 8, pp. 161–174. [Google Scholar] [CrossRef] [Green Version]
- Hazen, R.M.; Ewing, R.C.; Sverjensky, D.A. Evolution of uranium and thorium minerals. Am. Mineral. 2009, 94, 1293–1311. [Google Scholar] [CrossRef]
- Maruyama, S.; Kurokawa, K.; Ebisuzaki, T.; Sawaki, Y.; Suda, K.; Santosh, M. Nine requirements for the origin of Earth’s life: Not at the hydrothermal vent, but in a nuclear geyser system. Geosci. Front. 2019, 10, 1337–1357. [Google Scholar] [CrossRef]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Ebisuzaki, T.; Maruyama, S. Nuclear geyser model of the origin of life: Driving force to promote the synthesis of building blocks of life. Geosci. Front. 2017, 8, 275–298. [Google Scholar] [CrossRef] [Green Version]
- Fox, S.W. Thermal synthesis of amino acids and the origin of life. Geochim. Cosmochim. Acta 1995, 59, 1213–1214. [Google Scholar] [CrossRef]
- Segerer, A.H.; Burggraf, S.; Fiala, G.; Huber, G.; Huber, R.; Pley, U.; Stetter, K.O. Life in hot springs and hydrothermal vents. Orig. Life Evol. Biosph. 1993, 23, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Campbell, B.J.; Cary, S.C. Abundance of Reverse Tricarboxylic Acid Cycle Genes in Free-Living Microorganisms at Deep-Sea Hydrothermal Vents. Appl. Environ. Microbiol. 2004, 70, 6282–6289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cody, G.D. Geochemical Connections to Primitive Metabolism. Elements 2005, 1, 139–143. [Google Scholar] [CrossRef]
- Pinti, D.L.; Gargaud, M.; Irvine, W.M.; Pinti, D.L.; Gargaud, M.; Irvine, W.M.; Pinti, D.L. Encyclopedia of Astrobiology. Encycl. Astrobiol. 2011. [CrossRef] [Green Version]
- Huber, C.; Wachtershauser, G. Peptides by Activation of Amino Acids with CO on (Ni,Fe)S Surfaces: Implications for the Origin of Life. Science 1998, 281, 670–672. [Google Scholar] [CrossRef] [Green Version]
- Murphy, R.; Strongin, D. Surface reactivity of pyrite and related sulfides. Surf. Sci. Rep. 2009, 64, 1–45. [Google Scholar] [CrossRef]
- Cody, G.D. Primordial Carbonylated Iron-Sulfur Compounds and the Synthesis of Pyruvate. Science 2000, 289, 1337–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinen, W.; Lauwers, A.M. Organic sulfur compounds resulting from the interaction of iron sulfide, hydrogen sulfide and carbon dioxide in an anaerobic aqueous environment. Orig. Life Evol. Biosph. 1996, 26, 131–150. [Google Scholar] [CrossRef]
- Miller, S.L.; Bada, J.L. Submarine hot springs and the origin of life. Nature 1988, 334, 609–611. [Google Scholar] [CrossRef]
- Sousa, F.L.; Thiergart, T.; Landan, G.; Nelson-Sathi, S.; Pereira, I.A.C.; Allen, J.F.; Lane, N.; Martin, W.F. Early bioenergetic evolution. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20130088. [Google Scholar] [CrossRef] [Green Version]
- Coveney, R., Jr.; Goebel, E.D.; Zeller, E.J.; Dreschhoff, G.A.M.; Angino, E.E. Serpentinization and the Origin of Hydrogen Gas in Kansas. Am. Assoc. Pet. Geol. Bull. 1987, 71, 39–48. [Google Scholar] [CrossRef]
- White, L.M.; Shibuya, T.; Vance, S.D.; Christensen, L.E.; Bhartia, R.; Kidd, R.; Hoffmann, A.; Stucky, G.D.; Kanik, I.; Russell, M.J. Simulating serpentinization as it could apply to the emergence of life using the JPL hydrothermal reactor. Astrobiology 2020, 20, 307–326. [Google Scholar] [CrossRef]
- White, L.M.; Bhartia, R.; Stucky, G.D.; Kanik, I.; Russell, M.J. Mackinawite and greigite in ancient alkaline hydrothermal chimneys: Identifying potential key catalysts for emergent life. Earth Planet. Sci. Lett. 2015, 430, 105–114. [Google Scholar] [CrossRef]
- Macleod, G.; McKeown, C.; Hall, A.J.; Russell, M.J. Hydrothermal and oceanic pH conditions of possible relevance to the origin of life. Orig. Life Evol. Biosph. 1994, 24, 19–41. [Google Scholar] [CrossRef]
- Branscomb, E.; Russell, M.J. Turnstiles and bifurcators: The disequilibrium converting engines that put metabolism on the road. Biochim. Biophys. Acta Bioenerg. 2013, 1827, 62–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, M. Green Rust: The Simple Organizing ‘Seed’ of All Life? Life 2018, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J.; Daniel, R.M.; Hall, A.J. On the emergence of life via catalytic iron-sulphide membranes. Terra Nov. 1993, 5, 343–347. [Google Scholar] [CrossRef]
- Russell, M.J.; Nitschke, W.; Branscomb, E. The inevitable journey to being. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120254. [Google Scholar] [CrossRef]
- Ranjan, S.; Sasselov, D.D. Constraints on the Early Terrestrial Surface UV Environment Relevant to Prebiotic Chemistry. Astrobiology 2017, 17, 169–204. [Google Scholar] [CrossRef] [Green Version]
- Westall, F.; Hickman-Lewis, K.; Hinman, N.; Gautret, P.; Campbell, K.A.; Bréhéret, J.G.; Foucher, F.; Hubert, A.; Sorieul, S.; Dass, A.V.; et al. A Hydrothermal-Sedimentary Context for the Origin of Life. Astrobiology 2018, 18, 259–293. [Google Scholar] [CrossRef]
- Branscomb, E.; Russell, M.J. Frankenstein or a Submarine Alkaline Vent: Who Is Responsible for Abiogenesis?: Part 1: What is life–that it might create itself? BioEssays 2018, 40, 1–8. [Google Scholar] [CrossRef]
- Cottrell, A. The natural philosophy of engines. Contemp. Phys. 1979, 20, 1–10. [Google Scholar] [CrossRef]
- Leduc, S. The Mechanism of Life; Rebman: London, UK, 1911. [Google Scholar]
- Luisi, P. The Emergence of Life: From Chemical Origins to Synthetic Biology; Cambridge University Press: Cambridge, UK, 2006; ISBN 0521821177. [Google Scholar]
- Ruiz-Mirazo, K.; Peretó, J.; Moreno, A. A universal definition of life: Autonomy and open-ended evolution. Orig. Life Evol. Biosph. 2004, 34, 323–346. [Google Scholar] [CrossRef] [PubMed]
- Stano, P.; Luisi, P.L. Basic questions about the origins of life: Proceedings of the Erice international school of complexity (fourth course). Orig. Life Evol. Biosph. 2007, 37, 303–307. [Google Scholar] [CrossRef]
- Schneider, E.D.D.; Kay, J.J.J. Life as a manifestation of the second law of thermodynamics. Math. Comput. Model. 1994, 19, 25–48. [Google Scholar] [CrossRef]
- Boltzmann, L. The Second Law of Thermodynamics. In Theoretical Physics and Philosophical Problems; McGuiness, B., Ed.; Springer: Dordrecht, The Netherlands, 1974; Volume 5, pp. 13–32. ISBN 9788578110796. [Google Scholar]
- Schrodinger, E. What is Life? Cambridge University Press: Cambridge, UK, 1946; Volume 43, ISBN 9781139644129. [Google Scholar]
- De Wolf, T.; Holvoet, T. Emergence versus self-organisation: Different concepts but promising when combined. Lect. Notes Comput. Sci. 2005, 3464 LNAI, 1–15. [Google Scholar] [CrossRef]
- Lebon, L.; Non-equilibrium, J.U.; Kondepudi, D.K.; Prigogine, I. Modern Thermodynamics: From Heat Engines to Dissipative Structures, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Branscomb, E.; Russell, M.J. Frankenstein or a Submarine Alkaline Vent: Who is Responsible for Abiogenesis?: Part 2: As life is now, so it must have been in the beginning. BioEssays 2018, 40, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Michaelian, K. Entropy Production and the Origin of Life. J. Mod. Phys. 2011, 2, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Endres, R.G. Entropy production selects nonequilibrium states in multistable systems. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Kondepudi, D.; Kay, B.; Dixon, J. Dissipative structures, machines, and organisms: A perspective. Chaos An Interdiscip. J. Nonlinear Sci. 2017, 27, 104607. [Google Scholar] [CrossRef] [PubMed]
- Lane, N. Proton gradients at the origin of life. BioEssays 2017, 39, 1600217. [Google Scholar] [CrossRef] [Green Version]
- Lane, N. Why Are Cells Powered by Proton Gradients? Nat. Educ. 2010, 3, 2. [Google Scholar] [CrossRef]
- Barge, L.M.; Doloboff, I.J.; White, L.M.; Stucky, G.D.; Russell, M.J.; Kanik, I. Characterization of iron-phosphate-silicate chemical garden structures. Langmuir 2012, 28, 3714–3721. [Google Scholar] [CrossRef] [PubMed]
- Astumian, R.D.; Mukherjee, S.; Warshel, A. The Physics and Physical Chemistry of Molecular Machines. ChemPhysChem 2016, 17, 1719–1741. [Google Scholar] [CrossRef] [Green Version]
- Goodsell, D.S. The Machinery of Life; Copernicus: New York, NY, UK, 1998; ISBN 978-0-387-84924-9. [Google Scholar]
- Arrhenius, G.O.; Diego, S.; Jolla, L.; Arrhenius, G.O. Crystals and life. Helv. Chim. Acta 2003, 86, 1569–1586. [Google Scholar] [CrossRef]
- Duval, S.; Baymann, F.; Schoepp-cothenet, B.; Trolard, F.; Bourrié, G.; Grauby, O.; Branscomb, E.; Russell, M.J.; Nitschke, W.; Grauby, O.; et al. Fougerite: The not so simple progenitor of the first cells. Interface Focus 2019, 9, 20190063. [Google Scholar] [CrossRef] [Green Version]
- Lane, N.; Allen, J.F.; Martin, W. How did LUCA make a living? Chemiosmosis in the origin of life. BioEssays 2010, 32, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Sojo, V.; Herschy, B.; Whicher, A.; Camprubí, E.; Lane, N. The Origin of Life in Alkaline Hydrothermal Vents. Astrobiology 2016, 16, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Russell, M.J. On the origin of biochemistry at an alkaline hydrothermal vent. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1887–1926. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P. Coupling of Phosphorylation to Electron and Hydrogen Transfer by a Chemi-Osmotic type of Mechanism. Nature 1961, 191, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P. Chemiosmotic Coupling in Energy Transduction: A Logical DevelopDlent of Biochemical Knowledge. In Membrane Structure and Mechanisms of Biological Energy Transduction; Avery, J., Ed.; Springer: Boston, MA, USA, 1972; pp. 5–24. ISBN 978-1-4684-2018-0. [Google Scholar]
- Mitchell, P. Keilin’s respiratory chain concept and its chemiosmotic consequences. Science 1979, 206, 1148–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, P. Vectorial chemistry and the molecular mechanics of chemiosmotic coupling: Power transmission by proticity. Biochem. Soc. Trans. 1976, 4, 399–430. [Google Scholar] [CrossRef] [PubMed]
- Prebble, J.N. The philosophical origins of Mitchell’s chemiosmotic concepts: The personal factor in scientific theory formulation. J. Hist. Biol. 2001, 34, 433–460. [Google Scholar] [CrossRef]
- Ljungdahl, L.G. The Acetyl-CoA Pathway and the Chemiosmotic Generation of ATP during Acetogenesis. In Acetogenesis; Springer US: Boston, MA, USA, 1994; pp. 63–87. [Google Scholar]
- Martin, W.; Russell, M.J. On the origins of cells: A hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2003, 358, 59–85. [Google Scholar] [CrossRef] [Green Version]
- Pinti, D.L. The Origin and Evolution of the Oceans. In Lectures in Astrobiology; Gargaud, M., Barbier, B., Martin, H., Reisse, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 83–112. [Google Scholar]
- Zahnle, K.; Arndt, N.; Cockell, C.; Halliday, A.; Nisbet, E.; Selsis, F.; Sleep, N.H. Emergence of a habitable planet. Space Sci. Rev. 2007, 129, 35–78. [Google Scholar] [CrossRef]
- Major, T.A.; Burd, H.; Whitman, W.B. Abundance of 4Fe-4S motifs in the genomes of methanogens and other prokaryotes. FEMS Microbiol. Lett. 2004, 239, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Roldan, A.; Hollingsworth, N.; Roffey, A.; Islam, H.-U.U.; Goodall, J.B.M.M.; Catlow, C.R.A.A.; Darr, J.A.; Bras, W.; Sankar, G.; Holt, K.B.; et al. Bio-inspired CO2 conversion by iron sulfide catalysts under sustainable conditions. Chem. Commun. 2015, 51, 7501–7504. [Google Scholar] [CrossRef] [Green Version]
- Nitschke, W.; McGlynn, S.E.; Milner-White, E.J.; Russell, M.J. On the antiquity of metalloenzymes and their substrates in bioenergetics. Biochim. Biophys. Acta Bioenerg. 2013, 1827, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Kitadai, N.; Nakamura, R. Chemical diversity of metal sulfide minerals and its implications for the origin of life. Life 2018, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Dobbek, H. Crystal Structure of a Carbon Monoxide Dehydrogenase Reveals a [Ni-4Fe-5S] Cluster. Science 2001, 293, 1281–1285. [Google Scholar] [CrossRef] [Green Version]
- Darnault, C.; Volbeda, A.; Kim, E.J.; Legrand, P.; Vernède, X.; Lindahl, P.A.; Fontecilla-Camps, J.C. Ni-Zn-[Fe4-S4] and Ni-Ni-[Fe4-S4] clusters in closed and open α subunits of acetyl-CoA synthase/carbon monoxide dehydrogenase. Nat. Struct. Mol. Biol. 2003, 10, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Trolard, F.; Bourri, G. Fougerite a Natural Layered Double Hydroxide in Gley Soil: Habitus, Structure, and Some Properties. In Clay Minerals in Nature—Their Characterization, Modification and Application; Valaskova, M., Martynková, G.S., Eds.; InTech: Nappanee, IN, USA, 2012; pp. 171–188. [Google Scholar]
- Russell, M.J. The alkaline solution to the emergence of life: Energy, entropy and early evolution. Acta Biotheor. 2007, 55, 133–179. [Google Scholar] [CrossRef]
- Nitschke, W.; Russell, M.J. Beating the acetyl coenzyme a-pathway to the origin of life. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielke, R.E.; Russell, M.J.; Wilson, P.R.; McGlynn, S.E.; Coleman, M.; Kidd, R.; Kanik, I. Design, fabrication, and test of a hydrothermal reactor for origin-of-life experiments. Astrobiology 2010, 10, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Herschy, B.; Whicher, A.; Camprubi, E.; Watson, C.; Dartnell, L.; Ward, J.; Evans, J.R.G.; Lane, N. An Origin-of-Life Reactor to Simulate Alkaline Hydrothermal Vents. J. Mol. Evol. 2014, 79, 213–227. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Nakamura, R.; Takai, K. Deep-Sea Hydrothermal Fields as Natural Power Plants. ChemElectroChem 2018, 5, 2162–2166. [Google Scholar] [CrossRef]
- Nakamura, R.; Takashima, T.; Kato, S.; Takai, K.; Yamamoto, M.; Hashimoto, K. Electrical current generation across a black smoker chimney. Angew. Chem. Int. Ed. 2010, 49, 7692–7694. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, J.H.E.E.; García-Ruiz, J.M.; Novella, M.L.; Otálora, F.; Garc, J.M.; García-Ruiz, J.M.; Novella, M.L.; Otálora, F. Formation of chemical gardens. J. Colloid Interface Sci. 2002, 256, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Barge, L.M.; Cardoso, S.S.S.; Cartwright, J.H.E.; Cooper, G.J.T.; Cronin, L.; De Wit, A.; Doloboff, I.J.; Escribano, B.; Goldstein, R.E.; Haudin, F.; et al. From chemical gardens to chemobrionics. Chem. Rev. 2015, 115, 8652–8703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, A.L. La Plasmogenia: Nueva ciencia del origen de la vida. Cuad. Cult. 1932, 49, 1–38. [Google Scholar]
- Glaab, F.; Kellermeier, M.; Kunz, W.; Morallon, E.; García-Ruiz, J.M. Formation and Evolution of Chemical Gradients and Potential Differences Across Self-Assembling Inorganic Membranes. Angew. Chem. 2012, 124, 4393–4397. [Google Scholar] [CrossRef]
- Cartwright, J.H.E.; Escribano, B.; Sainz-Díaz, C.I. Chemical-Garden Formation, Morphology, and Composition. I. Effect of the Nature of the Cations. Langmuir 2011, 27, 3286–3293. [Google Scholar] [CrossRef]
- Barge, L.M.; Jones, J.-P.; Pagano, J.J.; Martinez, E.; Bescup, J. Three-Dimensional Analysis of a Simulated Prebiotic Hydrothermal Chimney. ACS Earth Sp. Chem. 2020, 4, 1663–1669. [Google Scholar] [CrossRef]
- Barge, L.M.; Abedian, Y.; Russell, M.J.; Doloboff, I.J.; Cartwright, J.H.E.; Kidd, R.D.; Kanik, I. From Chemical Gardens to Fuel Cells: Generation of Electrical Potential and Current Across Self-Assembling Iron Mineral Membranes. Angew. Chem. Int. Ed. 2015, 54, 8184–8187. [Google Scholar] [CrossRef] [Green Version]
- Barge, L.M.; Kee, T.P.; Doloboff, I.J.; Hampton, J.M.P.; Ismail, M.; Pourkashanian, M.; Zeytounian, J.; Baum, M.M.; Moss, J.A.; Lin, C.K.; et al. The fuel cell model of abiogenesis: A new approach to origin-of-life simulations. Astrobiology 2014, 14, 254–270. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.J.; Hall, A.J. The onset and early evolution of life. In Evolution of Early Earth’s Atmosphere, Hydrosphere, and Biosphere—Constraints from Ore Deposits; Stephen, E.K., Ohmoto, H., Eds.; Geological Society of America: Boulder, CO, USA, 2006; pp. 1–32. [Google Scholar]
- Barge, L.M.; Abedian, Y.; Doloboff, I.J.; Nuñez, J.E.; Russell, M.J.; Kidd, R.D.; Kanik, I. Chemical gardens as flow-through reactors simulating natural hydrothermal systems. J. Vis. Exp. 2015, 2015, 91109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielke, R.E.; Robinson, K.J.; White, L.M.; McGlynn, S.E.; McEachern, K.; Bhartia, R.; Kanik, I.; Russell, M.J. Iron-sulfide-bearing chimneys as potential catalytic energy traps at life’s emergence. Astrobiology 2011, 11, 933–950. [Google Scholar] [CrossRef]
- Russell, M.J.; Daia, D.E.; Hall, A.J. The Emergence of Life from FeS Bubbles at Alkaline Hot Springs in an Acid Ocean. In Thermophiles: The Keys to Molecular Evolution and the Origin of Life? Taylor & Francis: London, UK, 1998; pp. 77–126. [Google Scholar]
- Burcar, B.T.; Barge, L.M.; Trail, D.; Watson, E.B.; Russell, M.J.; McGown, L.B. RNA Oligomerization in Laboratory Analogues of Alkaline Hydrothermal Vent Systems. Astrobiology 2015, 15, 509–522. [Google Scholar] [CrossRef]
- Santosh, M.; Arai, T.; Maruyama, S. Hadean Earth and primordial continents: The cradle of prebiotic life. Geosci. Front. 2017, 8, 309–327. [Google Scholar] [CrossRef] [Green Version]
- McGlynn, S.E.; Kanik, I.; Russell, M.J. Peptide and RNA contributions to iron–sulphur chemical gardens as life’s first inorganic compartments, catalysts, capacitors and condensers. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2012, 370, 3007–3022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barge, L.M.; Doloboff, I.J.; Russell, M.J.; VanderVelde, D.; White, L.M.; Stucky, G.D.; Baum, M.M.; Zeytounian, J.; Kidd, R.; Kanik, I. Pyrophosphate synthesis in iron mineral films and membranes simulating prebiotic submarine hydrothermal precipitates. Geochim. Cosmochim. Acta 2014, 128, 1–12. [Google Scholar] [CrossRef]
- Lahav, N.; Nir, S.; Elitzur, A.C. The emergence of life on Earth. Prog. Biophys. Mol. Biol. 2001, 75, 75–120. [Google Scholar]
- Luoto, H.H.; Belogurov, G.A.; Baykov, A.A.; Lahti, R.; Malinen, A.M. Na+-translocating Membrane Pyrophosphatases Are Widespread in the Microbial World and Evolutionarily Precede H+-translocating Pyrophosphatases. J. Biol. Chem. 2011, 286, 21633–21642. [Google Scholar] [CrossRef] [Green Version]
- Baltscheffsky, H. Stepwise molecular evolution of bacterial photosynthetic energy conversion. Biosystems 1981, 14, 49–56. [Google Scholar] [CrossRef]
- Barge, L.M.; Flores, E.; Baum, M.M.; Velde, D.G.V.; Russell, M.J. Redox and pH gradients drive amino acid synthesis in iron oxyhydroxide mineral systems. Proc. Natl. Acad. Sci. USA 2019, 116, 4828–4833. [Google Scholar] [CrossRef] [Green Version]
- Filtness, M.J.; Butler, I.B.; Rickard, D. The origin of life: The properties of iron sulphide membranes. Trans. Inst. Min. Metall. Sect. B Appl. Earth Sci. 2003, 112, 21–23. [Google Scholar] [CrossRef]
- Russell, M.; Ponce, A. Six ‘Must-Have’ Minerals for Life’s Emergence: Olivine, Pyrrhotite, Bridgmanite, Serpentine, Fougerite and Mackinawite. Life 2020, 10, 291. [Google Scholar] [CrossRef]
- Koper, M.T.M. Theory of multiple proton-electron transfer reactions and its implications for electrocatalysis. Chem. Sci. 2013, 4, 2710–2723. [Google Scholar] [CrossRef] [Green Version]
- Rickard, D.; Luther, G.W. Chemistry of Iron Sulfides. Chem. Rev. 2007, 107, 514–562. [Google Scholar] [CrossRef] [PubMed]
- Mullet, M.; Boursiquot, S.; Abdelmoula, M.; Génin, J.M.; Ehrhardt, J.J. Surface chemistry and structural properties of mackinawite prepared by reaction of sulfide ions with metallic iron. Geochim. Cosmochim. Acta 2002, 66, 829–836. [Google Scholar] [CrossRef]
- Holmes, J. Fate of incorporated metals during mackinawite oxidation in sea water. Appl. Geochem. 1999, 14, 277–281. [Google Scholar] [CrossRef]
- Xiaoran, P. Investigatin the Lectrochemical Reduction of CO2 at Metal Sulfides; National University of Singapore: Singapore, 2006. [Google Scholar]
- Santos-Carballal, D.; Roldan, A.; Dzade, N.Y.; de Leeuw, N.H. Reactivity of CO2 on the surfaces of magnetite (Fe3O4), greigite (Fe3S4) and mackinawite (FeS). Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2018, 376, 20170065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzade, N.Y.; Roldan, A.; De Leeuw, N.H. Activation and dissociation of CO2 on the (001), (011), and (111) surfaces of mackinawite (FeS): A dispersion-corrected DFT study. J. Chem. Phys. 2015, 143, 094703. [Google Scholar] [CrossRef] [Green Version]
- Lennie, A.R.; Redfern, S.A.T.; Champness, P.E.; Stoddart, C.P.; Schofield, P.F.; Vaughan, D.J. Transformation of mackinawite to greigite: An in situ X-ray powder diffraction and transmission electron microscope study. Am. Mineral. 1997, 82, 302–309. [Google Scholar] [CrossRef]
- Zakaria, S.N.A. Insight into the Surface Structure of Iron (-Nickel) Sulfides in Aqueous Environments for CO2 Reduction Applications Using In-Situ Spectroelectrochemistry. Ph.D. Thesis, University College London, London, UK, 2017. [Google Scholar]
- Vladimirov, M.G.; Ryzhkov, Y.F.; Alekseev, V.A.; Bogdanovskaya, V.A.; Otroshchenko, V.A.; Kritsky, M.S. Electrochemical reduction of carbon dioxide on pyrite as a pathway for abiogenic formation of organic molecules. Orig. Life Evol. Biosph. 2004, 34, 347–360. [Google Scholar] [CrossRef]
- Yamaguchi, A.; Yamamoto, M.; Takai, K.; Ishii, T.; Hashimoto, K.; Nakamura, R. Electrochemical CO2 reduction by Ni-containing iron sulfides: How is CO2 electrochemically reduced at bisulfide-bearing deep-sea hydrothermal precipitates? Electrochim. Acta 2014, 141, 311–318. [Google Scholar] [CrossRef] [Green Version]
- Thornber, M.R. Mineralogical and electrochemical stability of the nickel-iron sulphides-pentlandite and violarite. J. Appl. Electrochem. 1983, 13, 253–267. [Google Scholar] [CrossRef]
- Warner, T.E.; Rice, N.M.; Taylor, N. Electrochemical study of oxidative dissolution of synthetic violarite in aqueous media. In Hydrometallurgy ’94; Springer: Dordrecht, The Netherlands, 1994; pp. 273–287. [Google Scholar]
- Kitadai, N.; Nakamura, R.; Yamamoto, M.; Takai, K.; Li, Y.; Yamaguchi, A.; Gilbert, A.; Ueno, Y.; Yoshida, N.; Oono, Y. Geoelectrochemical CO production: Implications for the autotrophic origin of life. Sci. Adv. 2018, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yamaguchi, A.; Yamamoto, M.; Takai, K.; Nakamura, R. Molybdenum Sulfide: A Bioinspired Electrocatalyst for Dissimilatory Ammonia Synthesis with Geoelectrical Current. J. Phys. Chem. C 2017, 121, 2154–2164. [Google Scholar] [CrossRef]
- Batista, B.C.; Steinbock, O. Growing Inorganic Membranes in Microfluidic Devices: Chemical Gardens Reduced to Linear Walls. J. Phys. Chem. C 2015, 119, 27045–27052. [Google Scholar] [CrossRef]
- Hudson, R.; de Graaf, R.; Strandoo Rodin, M.; Ohno, A.; Lane, N.; McGlynn, S.E.; Yamada, Y.M.A.; Nakamura, R.; Barge, L.M.; Braun, D.; et al. CO 2 reduction driven by a pH gradient. Proc. Natl. Acad. Sci. USA 2020, 117, 22873–22879. [Google Scholar] [CrossRef]
- Möller, F.M.; Kriegel, F.; Kieß, M.; Sojo, V.; Braun, D. Steep pH Gradients and Directed Colloid Transport in a Microfluidic Alkaline Hydrothermal Pore. Angew. Chem. 2017, 129, 2380–2384. [Google Scholar] [CrossRef]
- Jackson, J.B. Natural pH Gradients in Hydrothermal Alkali Vents Were Unlikely to Have Played a Role in the Origin of Life. J. Mol. Evol. 2016, 83, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Jackson, J.B. The “Origin-of-Life Reactor” and Reduction of CO2 by H2 in Inorganic Precipitates. J. Mol. Evol. 2017, 85, 1–7. [Google Scholar] [CrossRef]
- Jackson, J.B. Ancient living organisms escaping from, or imprisoned in, the vents? Life 2017, 7, 36. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.J. The “Water Problem”(sic), the Illusory Pond and Life’s Submarine Emergence—A Review. Life 2021, 11, 429. [Google Scholar] [CrossRef]
- Damer, B.; Deamer, D. The Hot Spring Hypothesis for an Origin of Life. Astrobiology 2020, 20, 429–452. [Google Scholar] [CrossRef] [Green Version]
- Milshteyn, D.; Damer, B.; Havig, J.; Deamer, D. Amphiphilic Compounds Assemble into Membranous Vesicles in Hydrothermal Hot Spring Water but Not in Seawater. Life 2018, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Ooka, H.; McGlynn, S.E.; Nakamura, R. Electrochemistry at Deep-Sea Hydrothermal Vents: Utilization of the Thermodynamic Driving Force towards the Autotrophic Origin of Life. ChemElectroChem 2019, 6, 1316–1323. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, A.; Li, Y.; Takashima, T.; Hashimoto, K.; Nakamura, R. CO2 reduction using an electrochemical approach from chemical, biological, and geological aspects in the ancient and modern earth. Lect. Notes Energy 2016, 32, 213–228. [Google Scholar] [CrossRef]
- Altair, T.; Sartori, L.M.; Rodrigues, F.; de Avellar, M.G.B.; Galante, D. Natural Radioactive Environments As Sources of Local Disequilibrium for the Emergence of Life. Astrobiology 2020, 20, 1489–1497. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Basic Injection Method | |||
| Reservoir Solution (Early Ocean Simulant) Composition | Injected Solution (Early Hydrothermal Fluid) Composition | Maximum Potential (mV) | |
| FeCl2 + FeCl3 + NaNO3 | NaOH + CH3OH | 1200 | |
| FeCl2 | Na2S | 1400 | |
| FeCl2 + NaNO3 | NaOH | 640 | |
| Substitution of the interior fluid after the membrane formation | |||
| Reservoir solution (early ocean simulant) composition | Injected solution (early hydrothermal fluid) composition | Maximum potential (mV) | Maximum potential after solution substitution a (mV) |
| FeCl2 + FeCl3 | NaOH | 431 | 881 |
| NaOH + K4P2O7 | 473 | 914 | |
| NaOH + alanine | 485 | 929 | |
| NaOH + K4P2O7 + alanine | 239 | 923 | |
| Surface | Electrochemical Surface CO2 Reduction Product | Condition | Current Efficiency (%) | Reference |
|---|---|---|---|---|
| Fe3S4 | CO | Room temperature; pH = 5.5; 760 torr CO2; 1.3 V (vs. NHE) | <0.01 | Yamaguchi et al. (2014) [141] |
| CH4 | <0.01 | |||
| Formic acid | Room temperature; room pressure; E from 0 to −1 V a | 0.1 (pH = 4.5) 1.51 (pH = 6.5) 22.43 (pH = 10.5) | Roldan et al. (2015) [97] | |
| Acetic acid | 0.23 (pH = 4.5) 2.61 (pH = 6.5) 0.5 (pH = 10.5) | |||
| Methanol | 0.21 (pH = 4.5) 1.21 (pH = 6.5) | |||
| Pyruvic acid | 2.78 (pH = 6.5) | |||
| Ni—Fe3S4 | CO | Room temperature; pH = 5.5; pCO2 = 1 torr; E = 1.3 V b | <0.015 ~0.15 (Fe/Ni = 1; w/TEA) c ~0.05 (Fe/Ni = 1; w/PAH) d | Yamaguchi et al. (2014) [141] |
| CH4 | <0.01 (Fe/Ni = 5) ~0.075 (Fe/Ni = 1) ~0.25 (Fe/Ni = 1; w/TEA) c ~0.35 (Fe/Ni = 1; w/PAH) d | |||
| Pyrite | Acetic acid | Room temperature; pH = 7; pCO2 = 50 atm | ~0.025 (E = 0.8 V) a 0.12 (E ~ 1 V) a ~0.09 (E = 1.2 V) a | Vladimirov et al. (2004) [140] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altair, T.; Borges, L.G.F.; Galante, D.; Varela, H. Experimental Approaches for Testing the Hypothesis of the Emergence of Life at Submarine Alkaline Vents. Life 2021, 11, 777. https://doi.org/10.3390/life11080777
Altair T, Borges LGF, Galante D, Varela H. Experimental Approaches for Testing the Hypothesis of the Emergence of Life at Submarine Alkaline Vents. Life. 2021; 11(8):777. https://doi.org/10.3390/life11080777
Chicago/Turabian StyleAltair, Thiago, Luiz G. F. Borges, Douglas Galante, and Hamilton Varela. 2021. "Experimental Approaches for Testing the Hypothesis of the Emergence of Life at Submarine Alkaline Vents" Life 11, no. 8: 777. https://doi.org/10.3390/life11080777
APA StyleAltair, T., Borges, L. G. F., Galante, D., & Varela, H. (2021). Experimental Approaches for Testing the Hypothesis of the Emergence of Life at Submarine Alkaline Vents. Life, 11(8), 777. https://doi.org/10.3390/life11080777