
Transcriptomic Analysis of Long Noncoding RNA and mRNA Expression Profiles in the Amygdala of Rats with Bone Cancer Pain-Depression Comorbidity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Surgical Procedure to Establish a Rat Model of BCP
2.3. Behavioral Tests
2.4. Tissue Collection, RNA Extraction, Library Construction, and Sequencing
2.5. Quantitative Reverse Transcription PCR (RT-qPCR)
2.6. Western Blot Analysis
2.7. Targets Prediction of DElncRNAs
2.8. Gene Ontology (GO) Annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analysis
2.9. Construction of a Protein–Protein Interaction (PPI) Network Using Data of the Top 200 Differentially Expressed mRNAs (DEmRNAs)
2.10. Statistical Analysis
3. Results
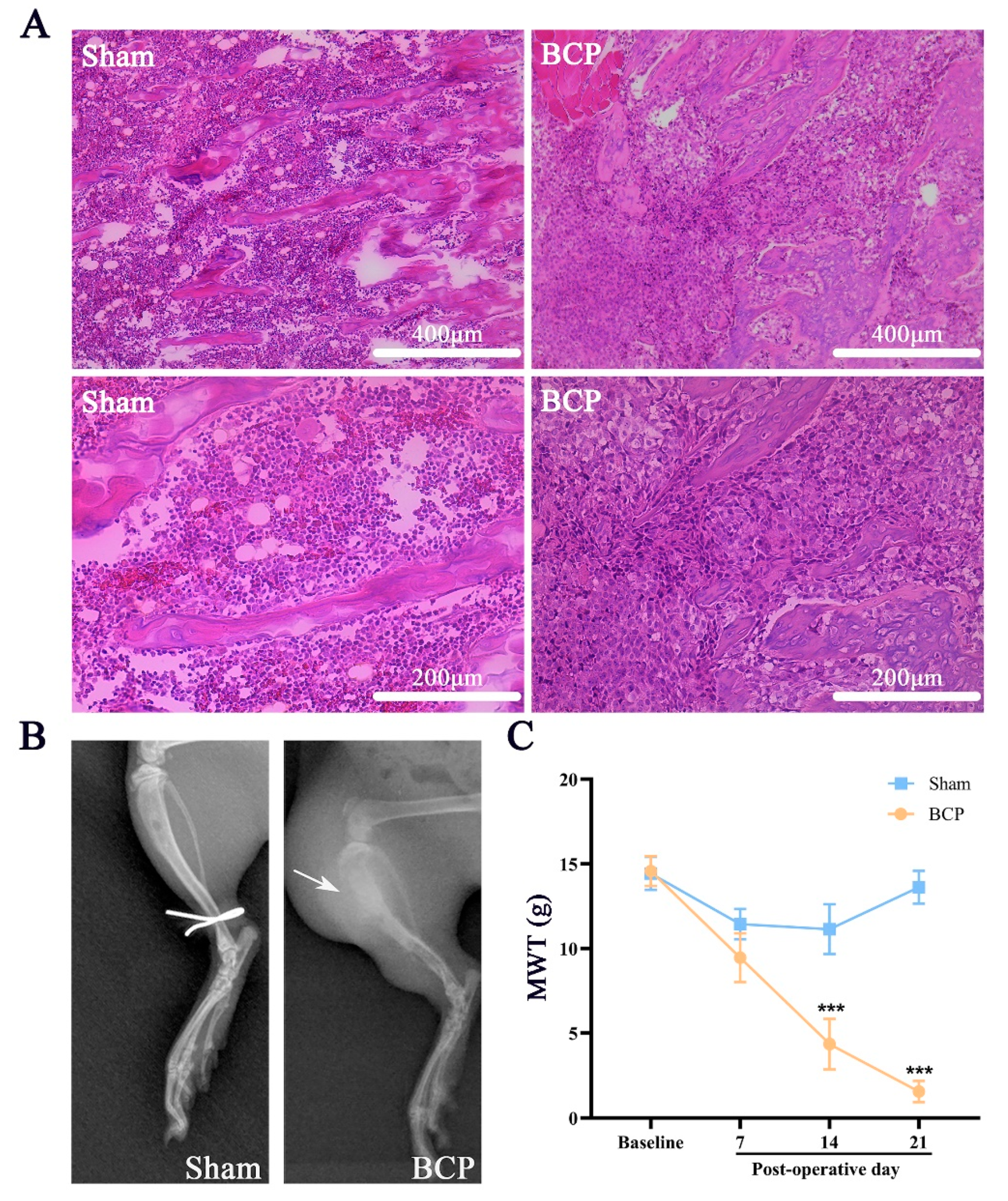
3.1. Implantation of MRMT-1 Carcinoma Cells Led to the Development of Mechanical Allodynia, Osteolytic Lesions, and Depressive Behaviors
3.2. Changes in Gene Expression Profiles Detected by High-Throughput RNA Sequencing
3.3. Validation of DElncRNA and DEmRNA Changes per RT-qPCR Test
3.4. Biological Functional Analysis of DEGs
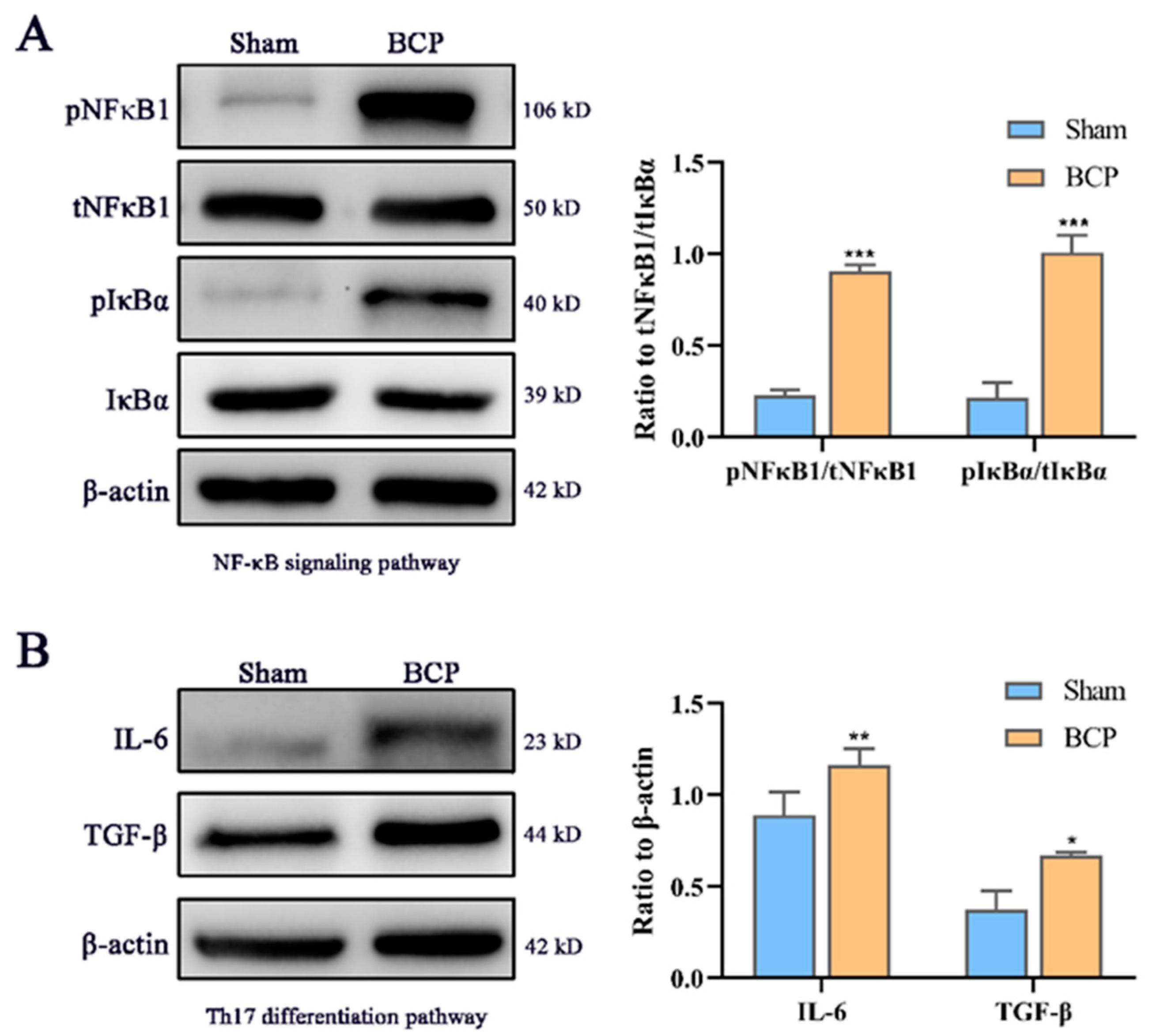
3.5. Western Blot Analyses of Proteins and Protein-Protein Interaction (PPI) Network Related to Immunity and Inflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zajączkowska, R.; Kocot-Kępska, M.; Leppert, W.; Wordliczek, J. Bone Pain in Cancer Patients: Mechanisms and Current Treatment. Int. J. Mol. Sci. 2019, 20, 6047. [Google Scholar] [CrossRef] [Green Version]
- Bortolato, B.; Hyphantis, T.N.; Valpione, S.; Perini, G.; Maes, M.; Morris, G.; Kubera, M.; Köhler, C.A.; Fernandes, B.; Stubbs, B.; et al. Depression in cancer: The many biobehavioral pathways driving tumor progression. Cancer Treat. Rev. 2017, 52, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Faron-Górecka, A.; Kuśmider, M.; Solich, J.; Kolasa, M.; Szafran, K.; Żurawek, D.; Pabian, P.; Dziedzicka-Wasylewska, M. Involvement of prolactin and somatostatin in depression and the mechanism of action of antidepressant drugs. Pharmacol. Rep. 2013, 65, 1640–1646. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Wang, Y.; Chen, J.; He, Y.; Zeng, Q.; Huang, Y.; Xu, X.; Lu, J.; Wang, Z.; Sun, X.; et al. The comorbidity of mental and physical disorders with self-reported chronic back or neck pain: Results from the China Mental Health Survey. J. Affect. Disord. 2020, 260, 334–341. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for the Pharmacological and Radiotherapeutic Management of Cancer Pain in Adults and Adolescents; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Lipton, A.; Uzzo, R.; Amato, R.J.; Ellis, G.K.; Hakimian, B.; Roodman, G.D.; Smith, M.R. The Science and Practice of Bone Health in Oncology: Managing Bone Loss and Metastasis in Patients With Solid Tumors. J. Natl. Compr. Cancer Netw. 2009, 7, S1–S29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Zhang, Y.; Xia, S. LncRNA NNT-AS1 promotes non-small cell lung cancer progression through regulating miR-22-3p/YAP1 axis. Thorac. Cancer 2020, 11, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, E.; Bagheri-Hosseinabadi, Z.; De Toma, I.; Jafarisani, M.; Sadeghi, I. The importance of long non-coding RNAs in neuropsychiatric disorders. Mol. Asp. Med. 2019, 70, 127–140. [Google Scholar] [CrossRef]
- Liu, Z.; Li, X.; Sun, N.; Xu, Y.; Meng, Y.; Yang, C.; Wang, Y.; Zhang, K. Microarray Profiling and Co-Expression Network Analysis of Circulating lncRNAs and mRNAs Associated with Major Depressive Disorder. PLoS ONE 2014, 9, e93388. [Google Scholar] [CrossRef] [PubMed]
- Willingham, A.T.; Orth, A.P.; Batalov, S.; Peters, E.C.; Wen, B.G.; Aza-Blanc, P.; Hogenesch, J.B.; Schultz, P.G. A Strategy for Probing the Function of Noncoding RNAs Finds a Repressor of NFAT. Science 2005, 309, 1570–1573. [Google Scholar] [CrossRef]
- Ng, S.-Y.; Johnson, R.; Stanton, L.W. Human long non-coding RNAs promote pluripotency and neuronal differentiation by association with chromatin modifiers and transcription factors. EMBO J. 2011, 31, 522–533. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Luo, Y.-L.; Mao, Y.-S.; Ji, J.-L. The link between long noncoding RNAs and depression. Prog. Neuro. Psychopharmacol. Biol. Psychiatry 2017, 73, 73–78. [Google Scholar] [CrossRef]
- Hou, X.; Weng, Y.; Guo, Q.; Ding, Z.; Wang, J.; Dai, J.; Wei, A.; Song, Z. Transcriptomic analysis of long noncoding RNAs and mRNAs expression profiles in the spinal cord of bone cancer pain rats. Mol. Brain 2020, 13, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulk, B.G.V.D.; Meens, P.H.F.; Van Lang, N.D.J.; De Voogd, E.L.; Van Der Wee, N.J.A.; Rombouts, S.A.R.B.; Crone, E.A.; Vermeiren, R.R.J.M. Amygdala activation during emotional face processing in adolescents with affective disorders: The role of underlying depression and anxiety symptoms. Front. Hum. Neurosci. 2014, 8, 393. [Google Scholar] [CrossRef] [Green Version]
- Corder, G.; Ahanonu, B.; Grewe, B.F.; Wang, D.; Schnitzer, M.J.; Scherrer, G. An amygdalar neural ensemble that encodes the unpleasantness of pain. Science 2019, 363, 276–281. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Jin, Y.; Meng, Q.; Zhu, X.; Bai, T.; Tian, Y.; Mao, Y.; Wang, L.; Xie, W.; Zhong, H.; et al. A neural circuit for comorbid depressive symptoms in chronic pain. Nat. Neurosci. 2019, 22, 1649–1658. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Xuan, C.; Li, Y.; Piao, L.; Li, J.; Zhao, H. Role of the Lateral Habenula in Pain-Associated Depression. Front. Behav. Neurosci. 2017, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef]
- Liu, M.; Yang, H.; Fang, D.; Yang, J.-J.; Cai, J.; Wan, Y.; Chui, D.-H.; Han, J.-S.; Xing, G.-G. Upregulation of P2X3 receptors by neuronal calcium sensor protein VILIP-1 in dorsal root ganglions contributes to the bone cancer pain in rats. Pain 2013, 154, 1551–1568. [Google Scholar] [CrossRef]
- Donovan-Rodriguez, T.; Dickenson, A.H.; Urch, C.E. Gabapentin Normalizes Spinal Neuronal Responses That Correlate with Behavior in a Rat Model of Cancer-induced Bone Pain. Anesthesiology 2005, 102, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-Y.; Ding, J.-Q.; Tang, Q.; Hao, M.-M.; Wang, B.-H.; Wu, J.; Yu, L.-Z.; Jiao, M.; Luo, B.-H.; Xie, M.; et al. SIRT1 activation by SRT1720 attenuates bone cancer pain via preventing Drp1-mediated mitochondrial fission. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 587–598. [Google Scholar] [CrossRef]
- Liu, X.; Bu, H.; Liu, C.; Gao, F.; Yang, H.; Tian, X.; Xu, A.; Chen, Z.; Cao, F.; Tian, Y. Inhibition of glial activation in rostral ventromedial medulla attenuates mechanical allodynia in a rat model of cancer-induced bone pain. Acta Acad. Med. Wuhan 2012, 32, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Zhang, Y.E.; Ye, Z.-Q.; Liu, X.-Q.; Zhao, S.-Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2007, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, S.K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein–protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, X.; Chen, X.; Li, S.; Ho, I.H.T.; Liu, X.; Chan, M.T.V.; Wu, W.K.K. Emerging roles of long non-coding RNAs in neuropathic pain. Cell Prolif. 2019, 52, e12528. [Google Scholar] [CrossRef] [Green Version]
- Chubb, E.J.; Bradshaw, N.J.; Soares, D.; Porteous, D.; Millar, J.K. The DISC locus in psychiatric illness. Mol. Psychiatry 2007, 13, 36–64. [Google Scholar] [CrossRef]
- Butler, A.A.; Webb, W.; Lubin, F.D. Regulatory RNAs and control of epigenetic mechanisms: Expectations for cognition and cognitive dysfunction. Epigenomics 2016, 8, 135–151. [Google Scholar] [CrossRef] [Green Version]
- Ng, S.Y.; Lin, L.; Soh, B.S.; Stanton, L.W. Long noncoding RNAs in development and disease of the central nervous system. Trends Genet. 2013, 29, 461–468. [Google Scholar] [CrossRef]
- Luongo, T.S.; Eller, J.M.; Lu, M.-J.; Niere, M.; Raith, F.; Perry, C.; Bornstein, M.R.; Oliphint, P.; Wang, L.; McReynolds, M.R.; et al. SLC25A51 is a mammalian mitochondrial NAD+ transporter. Nature 2020, 588, 174–179. [Google Scholar] [CrossRef]
- Choi, S.-H.; Koh, D.-I.; Ahn, H.; Kim, J.Y.; Kim, Y.; Hur, M.-W. Cell fate decisions by c-Myc depend on ZBTB5 and p. Biochem. Biophys. Res. Commun. 2020, 533, 1247–1254. [Google Scholar] [CrossRef]
- Shi, Y.; Sun, C.; Ge, W.; Du, Y.; Hu, N. Circular RNA VMA21 ameliorates sepsis-associated acute kidney injury by regulating miR-9-3p/SMG1/inflammation axis and oxidative stress. J. Cell. Mol. Med. 2020, 24, 11397–11408. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, A.; Uygun, D.S.; Yang, L.; McNally, J.; Lopez-Huerta, V.G.; Arias-Garcia, M.A.; Baez-Nieto, D.; Allen, A.; Fitzgerald, M.; Choi, S.; et al. Effects of a patient-derived de novo coding alteration of CACNA1I in mice connect a schizophrenia risk gene with sleep spindle deficits. Transl. Psychiatry 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Masih, S.; Moirangthem, A.; Phadke, S.R. Homozygous Missense Variation in PNPLA8 Causes Prenatal-Onset Severe Neurodegeneration. Mol. Syndr. 2021, 12, 174–178. [Google Scholar] [CrossRef]
- Obara, I.; Telezhkin, V.; AlRashdi, I.; Chazot, P. Histamine, histamine receptors, and neuropathic pain relief. Br. J. Pharmacol. 2020, 177, 580–599. [Google Scholar] [CrossRef]
- Mehta, P.; Miszta, P.; Rzodkiewicz, P.; Michalak, O.; Krzeczyński, P.; Filipek, S. Enigmatic Histamine Receptor H4 for Potential Treatment of Multiple Inflammatory, Autoimmune, and Related Diseases. Life 2020, 10, 50. [Google Scholar] [CrossRef]
- Mucci, F.; Marazziti, D.; Della Vecchia, A.; Baroni, S.; Morana, P.; Carpita, B.; Mangiapane, P.; Morana, F.; Morana, B.; Dell’Osso, L. State-of-the-Art: Inflammatory and Metabolic Markers in Mood Disorders. Life 2020, 10, 82. [Google Scholar] [CrossRef] [PubMed]
- Jaracz, J.; Gattner, K.; Jaracz, K.; Górna, K. Unexplained Painful Physical Symptoms in Patients with Major Depressive Disorder: Prevalence, Pathophysiology and Management. CNS Drugs 2016, 30, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Shi, D.; Zhou, Y.; Zheng, H.; Xiang, H.; Tian, X.; Gao, F.; Manyande, A.; Cao, F.; Tian, Y.; et al. MHC-I promotes apoptosis of GABAergic interneurons in the spinal dorsal horn and contributes to cancer induced bone pain. Exp. Neurol. 2016, 286, 12–20. [Google Scholar] [CrossRef]
- Berghoff, A.S.; Ricken, G.; Widhalm, G.; Rajky, O.; Dieckmann, K.; Birner, P.; Bartsch, R.; Höller, C.; Preusser, M. Tumour-infiltrating lymphocytes and expression of programmed death ligand 1 (PD-L1) in melanoma brain metastases. Histopathology 2014, 66, 289–299. [Google Scholar] [CrossRef]
- Banks, W. Blood-Brain Barrier Transport of Cytokines: A Mechanism for Neuropathology. Curr. Pharm. Des. 2005, 11, 973–984. [Google Scholar] [CrossRef]
- Walker, A.K.; Kavelaars, A.; Heijnen, C.J.; Dantzer, R. Neuroinflammation and Comorbidity of Pain and Depression. Pharmacol. Rev. 2014, 66, 80–101. [Google Scholar] [CrossRef] [Green Version]
- Huo, W.; Liu, Y.; Lei, Y.; Zhang, Y.; Huang, Y.; Mao, Y.; Wang, C.; Sun, Y.; Zhang, W.; Ma, Z.; et al. Imbalanced spinal infiltration of Th17/Treg cells contributes to bone cancer pain via promoting microglial activation. Brain Behav. Immun. 2019, 79, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Zorrilla, E.P.; Luborsky, L.; McKay, J.R.; Rosenthal, R.; Houldin, A.; Tax, A.; McCorkle, R.; Seligman, D.A.; Schmidt, K. The Relationship of Depression and Stressors to Immunological Assays: A Meta-Analytic Review. Brain Behav. Immun. 2001, 15, 199–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, I.I.; McKenzie, B.S.; Zhou, L.; Tadokoro, C.E.; Lepelley, A.; Lafaille, J.J.; Cua, D.J.; Littman, D.R. The Orphan Nuclear Receptor RORγt Directs the Differentiation Program of Proinflammatory IL-17+ T Helper Cells. Cell 2006, 126, 1121–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Neriah, Y.; Karin, M. Inflammation meets cancer, with NF-κB as the matchmaker. Nat. Immunol. 2011, 12, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-L.; Jiang, G.-Q.; Wei, J.; Zhang, H.-H.; Chen, W.; Zhu, H.; Hu, S.; Jiang, X.; Xu, G.-Y. Enhanced binding capability of nuclear factor-κB with demethylated P2X3 receptor gene contributes to cancer pain in rats. Pain 2015, 156, 1892–1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caviedes, A.; Lafourcade, C.; Soto, C.; Wyneken, U. BDNF/NF-κB Signaling in the Neurobiology of Depression. Curr. Pharm. Des. 2017, 23, 3154–3163. [Google Scholar] [CrossRef]
- Su, W.-J.; Zhang, Y.; Chen, Y.; Gong, H.; Lian, Y.-J.; Peng, W.; Liu, Y.-Z.; Wang, Y.-X.; You, Z.-L.; Feng, S.-J.; et al. NLRP3 gene knockout blocks NF-κB and MAPK signaling pathway in CUMS-induced depression mouse model. Behav. Brain Res. 2017, 322, 1–8. [Google Scholar] [CrossRef]
- Kravtsova-Ivantsiv, Y.; Shomer, I.; Cohen-Kaplan, V.; Snijder, B.; Superti-Furga, G.; Gonen, H.; Sommer, T.; Ziv, T.; Admon, A.; Naroditsky, I.; et al. KPC1-Mediated Ubiquitination and Proteasomal Processing of NF-κB1 p105 to p50 Restricts Tumor Growth. Cell 2015, 161, 333–347. [Google Scholar] [CrossRef] [Green Version]
- Cartwright, T.; Perkins, N.D.; Wilson, C.L. NFKB1: A suppressor of inflammation, ageing and cancer. FEBS J. 2016, 283, 1812–1822. [Google Scholar] [CrossRef] [Green Version]
- Oguiza, A.; Recio, C.; Lázaro, I.; Mallavia, B.; Blanco, J.; Egido, J.; Gomez-Guerrero, C. Peptide-based inhibition of IκB kinase/nuclear factor-κB pathway protects against diabetes-associated nephropathy and atherosclerosis in a mouse model of type 1 diabetes. Diabetologia 2015, 58, 1656–1667. [Google Scholar] [CrossRef] [Green Version]
- Tsuge, M.; Yasui, K.; Ichiyawa, T.; Saito, Y.; Nagaoka, Y.; Yashiro, M.; Yamashita, N.; Morishima, T. Increase of tumor necrosis factor-α in the blood induces the early activation of matrix metalloproteinase-9 in the brain. Microbiol. Immunol. 2010, 54, 417–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echeverry, S.; Shi, X.Q.; Rivest, S.; Zhang, J. Peripheral Nerve Injury Alters Blood-Spinal Cord Barrier Functional and Molecular Integrity through a Selective Inflammatory Pathway. J. Neurosci. 2011, 31, 10819–10828. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, Z.; Yang, Z.; Ruan, J.; Luo, L.; Long, F.; Tang, D. Chanling Gao Attenuates Bone Cancer Pain in Rats by the Ikkβ/Nf-Κb Signaling Pathway. Front. Pharmacol. 2020, 11, 525. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Wang, Z.; Zhi, Z.; Tian, J.; Zhao, Y.; Sun, J. Targeting the TLR4/NF-κB pathway in β-amyloid-stimulated microglial cells: A possible mechanism that oxysophoridine exerts anti-oxidative and anti-inflammatory effects in an in vitro model of Alzheimer’s disease. Brain Res. Bull. 2021, 175, 150–157. [Google Scholar] [CrossRef]
- Liu, Y.; Feng, L.; Ren, S.; Zhang, Y.; Xue, J. Inhibition of lncRNA DILC attenuates neuropathic pain via the SOCS3/JAK2/STAT3 pathway. Biosci. Rep. 2020, 40, BSR20194486. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Description | log2FC | p-Value |
|---|---|---|---|
| Upregulation | |||
| RGD1310951 | similar to RIKEN cDNA | >10 | 3.15 × 10 −11 |
| Timp3 | TIMP metallopeptidase inhibitor 3 | >10 | 4.60 × 10−10 |
| Dnaja4 | DnaJ heat shock protein family (Hsp40) member A4 | 9.60 | 1.46 × 10−7 |
| S100a8 | S100 calcium binding protein A8 | 6.09 | 1.82 × 10−14 |
| Cxcr2 | C-X-C motif chemokine receptor 2 | 6.07 | 9.09 × 10−7 |
| Cd177 | CD177 molecule | 5.91 | 1.14 × 10−4 |
| Rhag | Rh-associated glycoprotein | 5.77 | 3.99 × 10−4 |
| Gata1 | GATA binding protein 1 | 8.08 | 6.78 × 10−4 |
| Mmp8 | matrix metallopeptidase 8 | 5.54 | 1.21 × 10−8 |
| S100a9 | S100 calcium binding protein A9 | 8.07 | 2.41 × 10−9 |
| Arntl | aryl hydrocarbon receptor nuclear translocator-like | 8.00 | 2.11 × 10−5 |
| Ngp | neutrophilic granule protein | 5.45 | 7.08 × 10−5 |
| Mcemp1 | mast cell-expressed membrane protein 1 | 5.22 | 9.90 × 10−5 |
| Nlrp12 | NLR family, pyrin domain containing 12 | 5.13 | 3.1 × 10−4 |
| Nfkb1 | nuclear factor kappa B subunit 1 | 7.09 | 7.068 × 10−3 |
| Downregulation | |||
| Vamp1 | vesicle-associated membrane protein 1 | <−10 | 5.20 × 10−21 |
| NEWGENE_1310561 | programmed cell death 5 | <−10 | 1.63 × 10−5 |
| Gabbr2 | gamma-aminobutyric acid type B receptor subunit 2 | <−10 | 8.08 × 10−17 |
| Lgals3 | galectin 3 | <−10 | 4.33 × 10−7 |
| Tmem8b | transmembrane protein 8B | −9.92 | 2.12 × 10−5 |
| Kif26a | kinesin family member 26A | −9.81 | 3.65 × 10−11 |
| Paqr8 | progestin and adipoQ receptor family member 8 | −8.11 | 7.99 × 10−4 |
| Dcun1d2 | defective in cullin neddylation 1 domain containing 2 | −8.08 | 3.428 × 10−3 |
| Sertad2 | SERTA domain containing 2 | −7.98 | 3.115 × 10−3 |
| Ccdc92 | coiled-coil domain containing 92 | −7.71 | 1.756 × 10−3 |
| Vcam1 | vascular cell adhesion molecule 1 | −1.73 | 6.764 × 10−3 |
| Cpxm2 | carboxypeptidase X | −2.62 | 5.99 × 10−5 |
| Zfp280c | zinc finger protein 280C | −7.35 | 9.41 × 10−3 |
| Capn5 | calpain 5 | −6.82 | 3.8 × 10−4 |
| LOC100910708 | aldose reductase-related protein 1-like | −4.50 | 4.18 × 10−4 |
| LncRNA ID | Chromosomal Locus | log2FC | p-Value |
|---|---|---|---|
| Upregulation | |||
| MSTRG.5652.5 | 14:46638968-46643403 | >10 | 5.12 × 10−9 |
| MSTRG.4754.3 | 12:44294988-44296879 | 5.44 | 1.01 × 10−10 |
| MSTRG.16920.1 | 9:1433268-1436339 | 1.88 | 1.13 × 10−4 |
| MSTRG.1734.2 | 1:215913711-215968992 | 3.43 | 5.45 × 10−34 |
| MSTRG.7026.1 | 16:66316521-66324304 | 1.75 | 3.15 × 10−5 |
| MSTRG.13760.6 | 5:156317010-156325525 | 1.64 | 6.11 × 10−5 |
| MSTRG.13760.7 | 5:156317010-156325525 | 1.64 | 6.11 × 10−5 |
| ENSRNOT00000092949 | 1:221158098-221158792 | >10 | 6.117 × 10−3 |
| MSTRG.14298.2 | 6:69986056-69996290 | 2.57 | 8.838 × 10−3 |
| MSTRG.5.2 | 1:1671268-1675416 | 1.99 | 7.356 × 10−3 |
| ENSRNOT00000079224 | 16:60008769-60018283 | 1.74 | 6.637 × 10−3 |
| ENSRNOT00000088948 | 16:59997072-60000092 | 8.75 | 1.8916 × 10−2 |
| MSTRG.1478.10 | 1:194865923-194974619 | 7.45 | 1.088 × 10−2 |
| MSTRG.5207.3 | 13:84531041-84536203 | 10.38 | 2.3133 × 10−2 |
| MSTRG.4300.2 | 12:3340366-3341210 | 1.42 | 1.24 × 10−5 |
| Downregulation | |||
| MSTRG.74.1 | 1:11962604-11967166 | −3.66 | 6.00 × 10−18 |
| MSTRG.14169.1 | 6:30629736-30630859 | −2.06 | 2.34 × 10−10 |
| MSTRG.12891.1 | 5:23347609-23377614 | <−10 | 1.91 × 10−8 |
| MSTRG.5653.1 | 14:46644977-46686204 | −3.20 | 2.05 × 10−4 |
| MSTRG.5652.3 | 14:46635183-46683415 | −3.03 | 1.256 × 10−3 |
| MSTRG.14261.1 | 6:52444440-52459777 | −1.57 | 4.80 × 10−6 |
| MSTRG.10204.1 | 20:7065377-7156807 | −8.48 | 1.507 × 10−3 |
| MSTRG.1810.2 | 1:220139299-220144318 | −9.14 | 3.027 × 10−3 |
| MSTRG.16973.1 | 9:10509195-10537693 | −1.51 | 8.48 × 10−3 |
| MSTRG.13242.1 | 5:91131894-91135016 | −1.53 | 6.741 × 10−3 |
| MSTRG.12287.1 | 4:112115082-112140886 | −2.04 | 3.733 × 10−3 |
| ENSRNOT00000092903 | 5:60538928-60540538 | −2.98 | 1.4 × 10−3 |
| MSTRG.17595.5 | 9:113964702-113968568 | −3.45 | 3.773 × 10−3 |
| ENSRNOT00000092608 | 7:121511097-121619283 | −5.74 | 2.635 × 10−3 |
| ENSRNOT00000077174 | 4:16946938-16953847 | −7.02 | 2.78 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Chen, X.; Huang, F.; Lin, M.; Chen, P.; Wan, H.; Gao, F.; Zheng, T.; Zheng, X. Transcriptomic Analysis of Long Noncoding RNA and mRNA Expression Profiles in the Amygdala of Rats with Bone Cancer Pain-Depression Comorbidity. Life 2021, 11, 834. https://doi.org/10.3390/life11080834
Wu S, Chen X, Huang F, Lin M, Chen P, Wan H, Gao F, Zheng T, Zheng X. Transcriptomic Analysis of Long Noncoding RNA and mRNA Expression Profiles in the Amygdala of Rats with Bone Cancer Pain-Depression Comorbidity. Life. 2021; 11(8):834. https://doi.org/10.3390/life11080834
Chicago/Turabian StyleWu, Shuyan, Xiaohui Chen, Fengyi Huang, Mingxue Lin, Pinzhong Chen, Haiyang Wan, Fei Gao, Ting Zheng, and Xiaochun Zheng. 2021. "Transcriptomic Analysis of Long Noncoding RNA and mRNA Expression Profiles in the Amygdala of Rats with Bone Cancer Pain-Depression Comorbidity" Life 11, no. 8: 834. https://doi.org/10.3390/life11080834
APA StyleWu, S., Chen, X., Huang, F., Lin, M., Chen, P., Wan, H., Gao, F., Zheng, T., & Zheng, X. (2021). Transcriptomic Analysis of Long Noncoding RNA and mRNA Expression Profiles in the Amygdala of Rats with Bone Cancer Pain-Depression Comorbidity. Life, 11(8), 834. https://doi.org/10.3390/life11080834