
Dynamics of Silurian Plants as Response to Climate Changes

 ,
, 
Abstract
:1. Introduction
1.1. Character of Early Land Plants and Their Classification
1.2. Silurian Plant Assemblages
2. Materials and Methods
3. Results
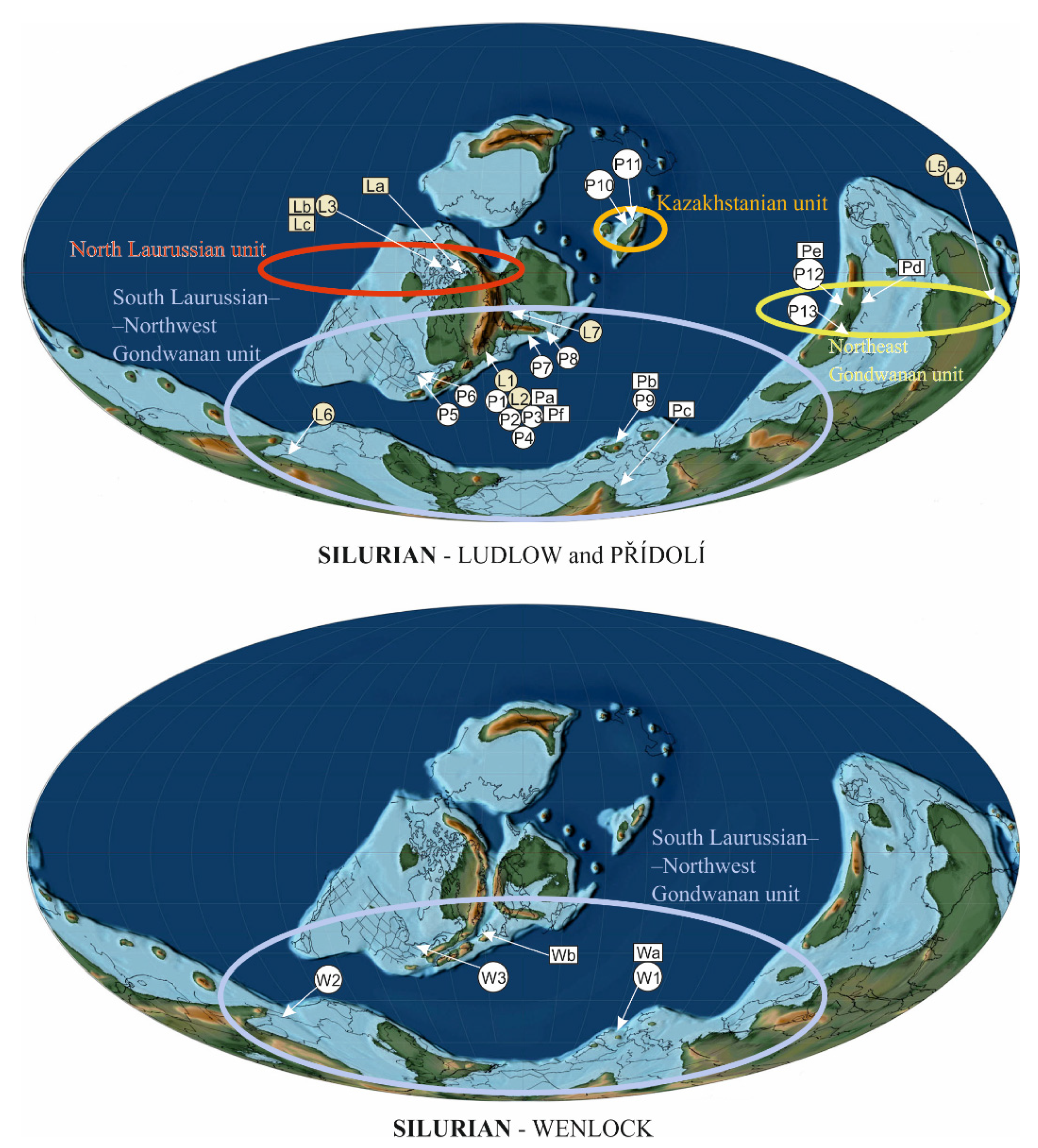
3.1. Environment of Silurian Plants
3.2. Diversity of Silurian Plants
3.2.1. North Laurussian Unit
3.2.2. South Laurussian–Northwest Gondwanan Unit
Northwest Gondwana


South Laurussia

3.2.3. Kazakhstanian Unit


3.2.4. Northeast Gondwanan Unit
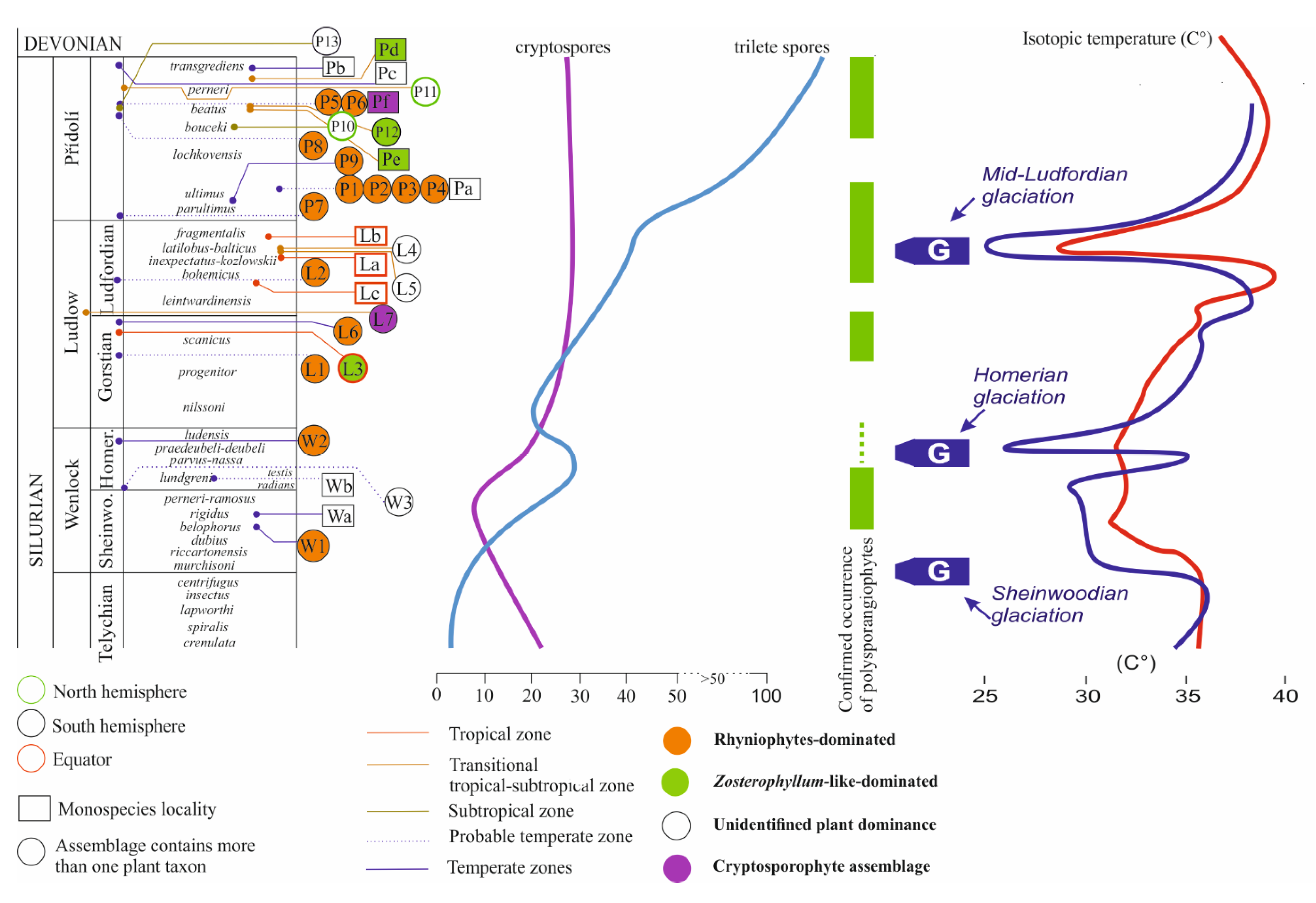
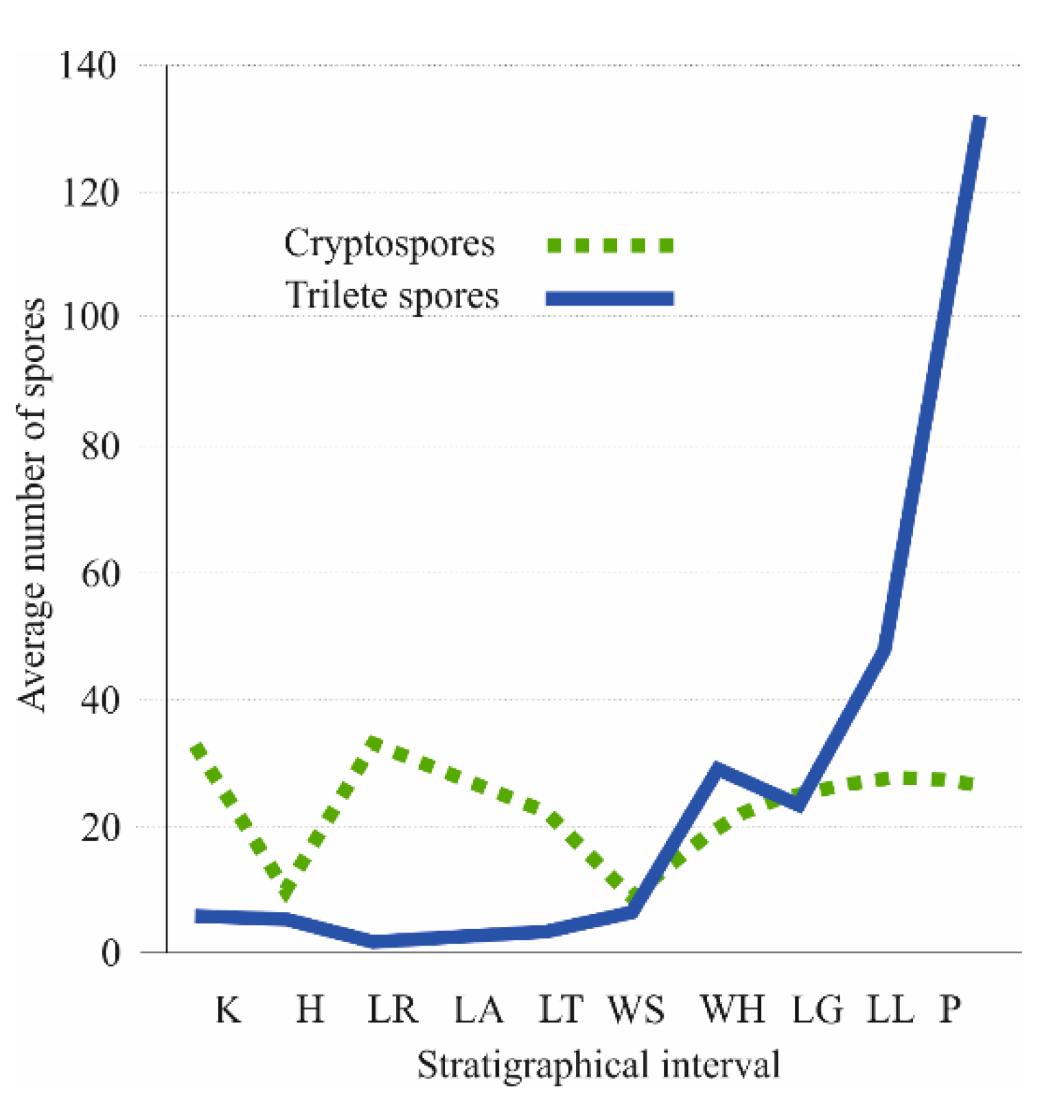
3.3. Palynological Aspects of Early Land Plants

4. Discussion
4.1. Response of Silurian Plants to Environmental Perturbations
4.1.1. Wenlock
4.1.2. Ludlow
4.1.3. The Importance of Mid-Ludfordian Glaciation
4.1.4. Přídolí
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wellman, C.H.; Strother, P.K. The terrestrial biota prior to the origin of land plants (embryophytes): A review of the evidence. Palaeontology 2015, 58, 601–627. [Google Scholar] [CrossRef]
- Kenrick, P. Changing expressions: A hypothesis for the origin of the vascular plant life cycle. Philos. Trans. R. Soc. B 2017, 373, 20170149. [Google Scholar] [CrossRef]
- Cooper, E.D.; Henwood, M.J.; Brown, E.A. Are the liverworts really that old? Cretaceous origins and Cenozoic diversifications in Lepidoziaceae reflect a recurrent theme in liverwort evolution. Biol. J. Linn. Soc. 2012, 107, 425–441. [Google Scholar] [CrossRef] [Green Version]
- Zhong, B.; Fong, R.; Collins, L.J.; McLenachan, P.A.; Penny, D. Two new fern chloroplasts and decelerated evolution linked to the long generation time in tree ferns. Genome Biol. Evol. 2014, 6, 1166–1173. [Google Scholar] [CrossRef] [Green Version]
- Willis, K.J.; McElwain, J.C. The Evolution of Plants; Oxford University Press: Oxford, UK, 2002; p. 378. [Google Scholar]
- Taylor, W.A.; Wellman, C.H. Ultrastructure of enigmatic phytoclasts (banded tubes) from the Silurian-lower Devonian: Evidence for affinities and role in early terrestrial ecosystems. Palaios 2009, 24, 167–180. [Google Scholar] [CrossRef]
- Edwards, D.; Morris, J.L.; Richardson, J.B.; Kenrick, P. Cryptospores and cryptophytes reveal hidden diversity in early land floras. New Phytol. 2014, 202, 50–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, D.; Kenrick, P. The early evolution of land plants, from fossils to genomics: A commentary on Lang (1937) ‘On the plant-remains from the Downtonian of England and Wales’. Philos. Trans. R. Soc. B 2015, 370, 20140343. [Google Scholar] [CrossRef] [Green Version]
- Tomescu, A.M.F.; Rothwell, G.W. Wetlands before tracheophytes: Thalloid terrestrial communities of the Early Silurian Passage Creek biota (Virginia). Geol. Soc. Am. Spec. Pap. 2006, 399, 41–56. [Google Scholar]
- Tomescu, A.M.F.; Honegger, R.; Rothwell, G.W. Earliest fossil record of bacterial–cyanobacterial mat consortia: The early Silurian Passage Creek biota (440 Ma, VA, USA). Geobiology 2008, 6, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Tomescu, A.M.F.; Tate, R.W.; Mack, N.G.; Calder, V.J. Simulating fossilization to resolve the taxonomic affinities of thalloid fossils in Early Silurian (ca. 425 Ma) terrestrial assemblages. Bibl. Lichenol. 2010, 105, 183–189. [Google Scholar]
- Rubinstein, C.V.; Gerrienne, P.; de la Puente, G.S.; Astini, R.A.; Steemans, P. Early middle Ordovician evidence for land plants in Argentina (eastern Gondwana). New Phytol. 2010, 188, 365–369. [Google Scholar] [CrossRef]
- Salamon, S.A.; Gerrienne, P.; Steemans, P.; Gorzelak, P.; Filipiak, P.; Le Hérissé, A.; Paris, F.; Cascales-Minana, B.; Brachaniec, T.; Misz-Kennan, M.; et al. Putative late Ordovician land plants. New Phytol. 2018, 218, 1305–1309. [Google Scholar] [CrossRef] [Green Version]
- Naugolnykh, S.V. Plants of the First Terrestrial Ecosystems. Her. Russ. Acad. Sci. 2019, 89, 502–511. [Google Scholar] [CrossRef]
- Morris, J.L.; Puttick, M.N.; Clark, J.W.; Edwards, D.; Kenrick, P.; Pressel, S.; Wellman, C.H.; Yang, Z.; Schneider, H.; Donoghue, P.C.J. The timescale of early land plant evolution. Proc. Natl. Acad. Sci. USA 2018, 115, E2274–E2283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steemans, P.; Le Hérissé, A.; Melvin, J.; Miller, M.A.; Paris, F.; Verniers, J.; Wellman, C.H. Origin and Radiation of the Earliest Vascular Land Plants. Science 2009, 324, 353. [Google Scholar] [CrossRef] [Green Version]
- Strother, P.K.; Beck, J. Spore–like microfossils from Middle Cambrian strata: Expanding the meaning of the term cryptospore, 413–424. In Pollen and Spores: Morphology and Biology; Harley, M.M., Morton, C.M., Blackmore, S., Eds.; Royal Botanic Gardens, Kew: Richmond, UK, 2000. [Google Scholar]
- Su, D.; Yang, L.; Shi, X.; Ma, X.; Zhou, X.; Hedges, S.B.; Zhong, B. Large-Scale Phylogenomic Analyses Reveal the Monophyly of Bryophytes and Neoproterozoic Origin of Land Plants. Mol. Biol. Evol. 2021, 1–13. [Google Scholar] [CrossRef]
- Schopf, J.M.; Mencher, E.; Boucot, A.J.; Andrews, H.N. Erect plants in the early Silurian of Maine. US Geol. Sur. Rest. VA Prof. Pap. 1966, 550-D, D69–D75. [Google Scholar]
- McAdams, N.E.B.; Cramer, B.D.; Bancroft, A.M.; Melchin, M.J.; Devera, J.A.; Day, J.E. Integrated δ13Ccarb, conodont, and graptolite biochemostratigraphy of the Silurian from the Illinois Basin and stratigraphic revision of the Bainbridge Group. GSA Bull. 2018, 131, 335–352. [Google Scholar] [CrossRef]
- Cramer, B.D.; Brett, C.E.; Melchin, M.J.; Männik, P.; Kleffner, M.A.; McLaughlin, P.I.; Lloydell, D.K.; Munnecke, A.; Jeppsson, L.; Corradini, C.; et al. Revised correlation of Silurian provincial series of North America with global and regional chronostratigraphic and δ13Ccarb chemostratigraphy. Lethaia 2011, 44, 185–202. [Google Scholar] [CrossRef]
- Melchin, M.J.; Sadler, P.M.; Cramer, B.D. Chapter 21—The Silurian period. In The Geological Time Scale; Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2020; pp. 695–732. [Google Scholar]
- Grossman, E.L.; Joachimski, M.M. Oxygen isotope stratigraphy. In The Geological Time Scale 2020; Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2020; pp. 279–307. [Google Scholar]
- Frýda, J.; Lehnert, O.; Joachimski, M.; Mannik, P.; Kubajko, M.; Mergl, M.; Farkaš, J.; Frýdová, B. The Mid-Ludfordian (late Silurian) Glaciation: A link with global changes in ocean chemistry and ecosystem overturns. Earth Sci. Rev. 2021, 220, 103652. [Google Scholar] [CrossRef]
- Gerrienne, P.; Servais, T.; Vecoli, M. Plant evolution and terrestrialization during Palaeozoic times—The phylogenetic context. Rev. Palaeobot. Palynol. 2016, 227, 4–18. [Google Scholar] [CrossRef]
- Edwards, D.; Axe, L.; Honeggert, R. Contributions to the diversity in cryptogamic covers in the mid-Palaeozoic: Nematothallus revisited. Bot. J. Linn. Soc. 2013, 173, 505–534. [Google Scholar] [CrossRef]
- Abbott, G.D.; Ewbank, G.; Edwards, D.; Wang, G.Y. Molecular characterization of some enigmatic Lower Devonian fossils. Geochim. Cosmochim. Acta 1998, 62, 1407–1418. [Google Scholar] [CrossRef]
- Edwards, D.; Axe, L. Evidence for a fungal affinity for Nematasketum, a closeally of Prototaxites. Bot. J. Linn. Soc. 2012, 168, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.Q.; Edwards, D.; Wu, P.C.; Duckett, J.G.; Hueber, F.M.; Li, C.S. Riccardiothallus devonicus gen. et sp. nov., the earliest simple thalloid liverwort from the Lower Devonian of Yunnan, China. Rev. Palaeobot. Palynol. 2012, 176–177, 35–40. [Google Scholar] [CrossRef]
- Strother, P.K.; Al-Hajri, S.; Traverse, A. New evidence for land plants from the lower Middle Ordovician of Saudi Arabia. Geology 1996, 24, 55–59. [Google Scholar] [CrossRef]
- Cox, C.J.; Li, B.; Foster, P.G.; Embley, T.M.; Civáň, P. Conflicting Phylogenies for Early Land Plants are Caused by Composition Biases among Synonymous Substitutions. Syst. Biol. 2014, 63, 272–279. [Google Scholar] [CrossRef]
- Kenrick, P.; Crane, P.R. Water-Conducting Cells in Early Fossil Land Plants: Implications for the Early Evolution of Tracheophytes. Bot. Gaz. 1991, 152, 335–356. [Google Scholar] [CrossRef]
- Cascales-Miñana, B.; Steemans, P.; Servais, T.; Lepot, K.; Gerrienne, P. An alternative model for the earliest evolution of vascular plants. Lethaia 2019, 52, 445–453. [Google Scholar] [CrossRef]
- Edwards, D. Embryophytic sporophytes in the Rhynie and Windyfield cherts. Trans. R. Soc. Edinb. Earth Sci. 2004, 94, 397–410. [Google Scholar] [CrossRef]
- Wellman, C.H. The invasion of the land by plants: When and where? New Phytol. 2010, 188, 306–309. [Google Scholar] [CrossRef]
- Graham, L.; Lewis, L.A.; Taylor, W.; Wellman, C.H.; Cook, M. Early terrestrialization: Transition from algal to bryophyte grade. In Photosynthesis in Bryophytes and Early Land Plants; Hanson, D.T., Rice, S.K., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 9–28. [Google Scholar]
- Harrison, C.J. Development and genetics in the evolution of land plant body plans. Philos. Trans. R. Soc. B 2017, 372, 20150490. [Google Scholar] [CrossRef] [Green Version]
- Kraft, P.; Pšenička, J.; Sakala, J.; Frýda, J. Initial plant diversification and dispersal event in upper Silurian of the Prague Basin. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 514, 144–155. [Google Scholar] [CrossRef]
- Kenrick, P.; Crane, P.R. The Origin and Early Diversification of Land Plants. Nature 1997, 389, 33–39. [Google Scholar] [CrossRef]
- Edwards, D. New insights into early land ecosystems: A glimpse of a Lilliputian world. Rev. Palaeobot. Palynol. 1996, 90, 159–174. [Google Scholar] [CrossRef]
- Gonez, P.; Gerrienne, P. Aberlemnia caledonica gen. et comb. nov., a new name for Cooksonia caledonica Edwards 1970. Rev. Palaeobot. Palynol. 2010, 163, 64–72. [Google Scholar] [CrossRef]
- Tims, J.D.J.; Chambers, T.C. Rhyniophytina and Trimerophytina from the early land flora of Victoria, Australia. Palaeontology 1984, 27, 265–279. [Google Scholar]
- Kraft, P.; Kvaček, Z. Where the lycophytes come from?—A piece of the story from the Silurian of peri-Gondwana. Gondwana Res. 2017, 45, 180–190. [Google Scholar] [CrossRef]
- Rickards, R.B. The age of the earliest club mosses: The Silurian Baragwanathia flora in Victoria, Australia. Geol. Mag. 2000, 137, 207–209. [Google Scholar] [CrossRef]
- Edwards, D.; Wellman, C.H. Embryophytes on Land: The Ordovician to Lochkovian (Lower Devonian) Record. In Plants Invade the land, Evolutionary and Environmental Perspectives; Gensel, P.G., Edwards, D., Eds.; Columbia University Press: New York, NY, USA, 2001; pp. 3–28. [Google Scholar]
- Edwards, D.; Richardson, J.B. Silurian and Lower Devonian plant assemblages from the Anglo-Welsh Basin: A palaeobotanical and palynological synthesis. Geol. J. 2004, 39, 375–402. [Google Scholar] [CrossRef]
- Raymond, A.; Gensel, P.; Stein, W.E. Phytogeography of Late Silurian macrofloras. Rev. Palaeobot. Palynol. 2006, 142, 165–192. [Google Scholar] [CrossRef]
- Wellman, C.H.; Steemans, P.; Vecoli, M. Paleophytogeography of Ordovician–Silurian land plants. In Early Palaeozoic Biogeography and Palaeogeography; Harper, D.T., Servais, T., Eds.; Geological Society, London, Memoirs: London, UK, 2013; Volume 38, pp. 461–476. [Google Scholar]
- Edwards, D. Constraints on Silurian and Early Devonian phytogeographic analysis based on megafossils. In Palaeozoic Palaeogeography and Biogeography; McKerrow, W.S., Scotese, C.R., Eds.; Geological Society, London, Memoirs: London, UK, 1990; pp. 233–242. [Google Scholar]
- Hao, S.; Xue, J.; Liu, Z.; Wang, D. Zosterophyllum penhallow around the Silurian-Devonian Boundary of Northeastern Yunnan, China. Int. J. Plant Sci. 2007, 168, 477–489. [Google Scholar] [CrossRef]
- Bateman, R.M.; Crane, P.R.; DiMichele, W.A.; Kenrick, P.R.; Rowe, N.P.; Speck, T.; Stein, W.E. Early evolution of land plants: Phylogeny, physiology, and ecology of the primary terrestrial radiation. Annul. Rev. Ecol. Syst. 1998, 29, 263–292. [Google Scholar] [CrossRef] [Green Version]
- Servais, T.; Cascales-Miñana, B.; Cleal, C.J.; Gerrienne, P.; Harper, D.A.T.; Neumann, M. Revisiting the Great Ordovician Diversification of land plants: Recent data and perspectives. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 534, 109280. [Google Scholar] [CrossRef]
- Servais, T.; Martin, R.E.; Nützel, A. The impact of the ‘terrestrialisation process’ in the late Palaeozoic: pCO2, pO2, and the ‘phytoplankton blackout’. Rev. Palaeobot. Palynol. 2016, 224, 26–37. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Boyce, C.K. Carbon sources for the Palaeozoic giant fungus Prototaxites inferred from modern analogues. Proc. R. Soc. B 2010, 277, 2149–2156. [Google Scholar] [CrossRef] [Green Version]
- Kenrick, P.; Wellman, C.H.; Schneider, H.; Edgecombe, G.D. A timeline for terrestrialization: Consequences for the carbon cycle in the Palaeozoic. Philos. Trans. R. Soc. London Ser. B 2012, 367, 519–536. [Google Scholar] [CrossRef] [PubMed]
- Trotter, J.A.; Williams, I.S.; Barnes, C.R.; Männik, P.; Simpson, A. New conodont δ18O records of Silurian climate change: Implications for environmental and biological events. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 443, 34–48. [Google Scholar] [CrossRef]
- Gambacorta, G.; Menichetti, E.; Trincianti, E.; Torricelli, S. The Silurian climatic transition recorded in the epicontinental Baltica Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 517, 16–29. [Google Scholar] [CrossRef]
- Signor, P.W.; Brett, C.E. The mid−Paleozoic precursor to the Mesozoic marine revolution. Paleobiology 1984, 10, 229–245. [Google Scholar] [CrossRef]
- Brett, C.E.; Walker, S.E. Predators and predation in Paleozoic marine environments. Paleontol. Soc. Pap. 2002, 8, 93–118. [Google Scholar] [CrossRef]
- Nützel, A.; Frýda, J. Palaeozoic plankton revolution: Evidence from early gastropod ontogeny. Geology 2003, 31, 829–831. [Google Scholar] [CrossRef]
- Klug, C.; Kroger, B.; Kiessling, W.; Mullins, G.L.; Servais, T.; Frýda, J.; Korn, D.; Turner, S. The Devonian nekton revolution. Lethaia 2010, 43, 465–477. [Google Scholar] [CrossRef] [Green Version]
- Berner, R.A. Phanerozoic atmospheric oxygen: New results using the GEOCARBSULF model. Am. J. Sci. 2009, 309, 603–606. [Google Scholar] [CrossRef]
- Berner, R.A.; Beerling, D.J.; Dudley, R.; Robinson, J.M.; Wildman, R.A. Phanerozoic atmospheric oxygen. Annu. Rev. Earth Planet. Sci. 2003, 31, 105–134. [Google Scholar] [CrossRef] [Green Version]
- Schachat, S.R.; Labandeira, C.C.; Saltzman, M.R.; Cramer, B.D.; Payne, J.L.; Boyce, C.K. Phanerozoic pO2 and the early evolution of terrestrial animals. Proc. R. Soc. B 2018, 285, 20172631. [Google Scholar] [CrossRef] [Green Version]
- Saltzman, M.R.; Thomas, E. Carbon isotope stratigraphy. In The Geologic Time Scale 2012; Gradstein, F., Ogg, J., Schmitz, M.D., Ogg, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 207–232. [Google Scholar] [CrossRef]
- Munnecke, A.; Samtleben, C.; Bickert, T. The Ireviken event in the Lower Silurian of Gotland, Sweden—Relation to similar Palaeozoic and Proterozoic events. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 195, 99–124. [Google Scholar] [CrossRef]
- Loydell, D.K. Reply to ‘Early Silurian positive δ13C excursions and their relationship to glaciations, sea-level changes and extinction events: Discussion’ by Bradley, D. Cramer and Axel Munnecke. Geol. J. 2008, 43, 511–515. [Google Scholar] [CrossRef]
- Calner, M. Silurian global events—At the tipping point of climate change. In Mass Extinctions; Ashraf, M.T., Ed.; Springer–Verlag: Berlin/Heidelberg, Germany, 2008; pp. 21–58. [Google Scholar]
- Lehnert, O.; Mannik, P.; Joachimski, M.M.; Calner, M.; Frýda, J. Palaeoclimate perturbations before the Sheinwoodian glaciation: A trigger for extinctions during the ‘Ireviken Event’. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 296, 320–331. [Google Scholar] [CrossRef]
- Veizer, J.; Ala, D.; Azmy, K.; Bruckschen, P.; Buhl, D.; Bruhn, F.; Carden, G.A.F.; Diener, A.; Ebneth, S.; Godderis, Y.; et al. 87Sr/86Sr, δ13C and δ18O evolution of Phanerozoic seawater. Chem. Geol. 1999, 161, 59–88. [Google Scholar] [CrossRef] [Green Version]
- Veizer, J.; Prokoph, A. Temperatures and oxygen isotopic composition of Phanerozoic oceans. Earth Sci. Rev. 2015, 146, 92–104. [Google Scholar] [CrossRef]
- Lécuyer, C.; Allemand, P. Modelling of the oxygen isotope evolution of seawater: Implication for the climate interpretation of the δ18O of marine sediments. Geochim. Cosmochim. Acta 1999, 63, 351–361. [Google Scholar] [CrossRef]
- Azmy, K.; Veizer, J.; Bassett, M.G.; Copper, P. Oxygen and carbon isotopic composition of Silurian brachiopods: Implications for coeval seawater and glaciations. Geol. Soc. Am. Bull. 1998, 100, 1499–1512. [Google Scholar] [CrossRef]
- Kotyk, M.E.; Basinger, J.F.; Gensel, P.G.; de Freitas, T.A. Morphologically complex plant macrofossils from the late Silurian of Arctic Canada. Am. J. Bot. 2002, 89, 1004–1013. [Google Scholar] [CrossRef]
- Larsen, P.-H.; Edwards, D.; Escher, J.C. Late Silurian plant megafossils from the Peary Land Group, North Greenland. Rapp. Gronlands Geol. Undersøgelse 1987, 133, 17–112. [Google Scholar]
- Scotese, C.R. Atlas of Silurian and Middle-Late Ordovician Paleogeographic Maps (Mollweide Projection), Maps 73–80, Volumes 5, The Early Paleozoic, PALEOMAP Atlas for ArcGIS; PALEOMAP Project: Evanston, IL, USA, 2014. [Google Scholar]
- Libertín, M.; Kvaček, J.; Bek, J.; Žárský, V.; Štorch, P. Sporophytes of polysporangiate land plants from the early Silurian period may have been photosynthetically autonomous. Nat. Plants 2018, 4, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Libertín, M.; Kvaček, J.; Bek, J. Plant diversity of the mid Silurian (lower Wenlock, Sheinwoodian) terrestrial vegetation preserved in marine sediments from the Barrandian area, the Czech Republic. Foss. Impr. 2018, 74, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Bek, J.; Štorch, P.; Tonarová, P.; Libertín, M. Early Silurian (mid-Sheinwoodian) palynomorphs from the Prague Basin, Czech Republic. Bull. Geosci. 2021. under review. [Google Scholar]
- Zicha, O. (Jugoslávských Partyzánů, Prague, Czech Republic). Personal communication, 2021. [Google Scholar]
- Honegger, R.; Edwards, D.; Axe, L.; Strullu-Derriens, C. Fertile Prototaxites taiti: A basal ascomycete with inoperculate, polysporous asci lacking croziers. Phil. Trans. R. Soc. B 2017, 373, 20170146. [Google Scholar] [CrossRef] [Green Version]
- Kříž, J. Silurian Field Excursions. Prague Basin (Barrandian), Bohemia; Geological Series of National Museum of Wales; National Museum of Wales: Cardiff, UK, 1992; Volume 13, pp. 1–111. [Google Scholar]
- Libertín, M.; Labuťa, R.; Dašková, J. The oldest vascular plants from the Bohemian Massif. Zprávy Geol. Výzkumech Roce 2002, 2002, 127. (In Czech) [Google Scholar]
- Horný, R. Studie o vrstvách budňanských v západní části barrandienského siluru. Sborník Ústředního Ust. Geol. 1955, 21, 315–448. [Google Scholar]
- Horný, R. Stratigraphy and tectonics of the western closures of the Silurian-Devonian synclinorium in the Barrandian area. Sborník Ústředního Ust. Geol. 1960, 26, 495–524. [Google Scholar]
- Obrhel, J. Die Flora der Pridoli-Schichten (Budnany-Stufe) des mittelböhmishcen Silurs. Geologie 1962, 11, 83–97. [Google Scholar]
- Schweitzer, H.J. Die Gattungen Renalia Gensel und Psilophyton Dawson im Unterdevon des Rheinlandes. Bonn. Palaeobot. Mitt. 1980, 6, 1–34. [Google Scholar]
- Kříž, J.; Jaeger, H.; Paris, F.; Schönlaub, H.P. Přídolí—The Fourth Subdivison of the Silurian. Jahrb. Geol. Bundesanst. 1986, 129, 291–360. [Google Scholar]
- Toro, M.; Chamon, O.; Salguero, R.; Vargas, C. Las plantas de la Formacion Kirusillas (Silurico) en la region de La Angostura Departamento de Cochabamba. In Memorias del XII Congreso Geológico de Bolivia–Tarija, Bolivia; Sociedad Geologica: Cochabamba, Boliviana, 1996; pp. 523–529. [Google Scholar]
- Edwards, D.; Banks, H.P.; Ciurca, S.J., Jr.; Laub, R.S. New Silurian cooksonias from dolostones of north-eastern North America. Bot. J. Linn. Soc. 2004, 146, 399–413. [Google Scholar] [CrossRef]
- Edwards, D.; Rogerson, E.C.W. New records of fertile Rhyniophytina from the late Silurian of Wales. Geol. Mag. 1979, 116, 93–98. [Google Scholar] [CrossRef]
- Douglas, J.; Holmes, F. The Baragwanathia story: An update. Foss. Collect. 2006, 77, 9–26. [Google Scholar]
- Edwards, D.; Morel, E.M.; Paredes, F.; Ganuza, D.G.; Zúniga, A. Plant assemblages from the Silurian of southern Bolivia and their palaeogeographic significance. Bot. J. Linn. Soc. 2001, 135, 229–250. [Google Scholar] [CrossRef]
- Hagström, J.; Mehlqvist, K. The dawn of terrestrial ecosystems on Baltica: First report on land plant remains and arthropod coprolites from the Upper Silurian of Gotland, Sweden. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 317–318, 162–170. [Google Scholar] [CrossRef]
- Rogerson, C.; Edwards, D.; Axe, L.; Davies, K.L. A new Embryophyte from the Upper Silurian of Shropshire, England. Spec. Pap. Palaeontol. 2002, 67, 233–249. [Google Scholar]
- Fanning, U.; Edwards, D.; Richardson, J.B. Further evidence for diversity in late Silurian land vegetation. J. Geol. Soc. Lond. 1990, 147, 725–728. [Google Scholar] [CrossRef]
- Fanning, U.; Edwards, D.; Richardson, J.B. A new rhyniophytoid from the late Silurian of the Welsh Borderland. Neues Jahrb. Geol. Paläontol. Abh. 1991, 183, 37–47. [Google Scholar]
- Fanning, U.; Richardson, J.B.; Edwards, D. A review of in situ spores in Silurian land plants. In Systematics Association, Pollen and Spores, Special Volume; Blackmore, S., Barnes, S.H., Eds.; Clarendon Press: Oxford, UK, 1991; Volume 44, pp. 25–47. [Google Scholar]
- Edwards, D. A late Silurian flora from the Lower Old Red Sandstone of South-West Dyfed. Palaeontology 1979, 22, 23–52. [Google Scholar]
- Bodzioch, A.; Kozlowski, W.; Poplawska, A. A Cooksonia−type flora from the Upper Silurian of the Holy Cross Mountains, Poland. Acta Palaeontol. Pol. 2003, 48, 653–656. [Google Scholar]
- Ishchenko, T.A. The Late Silurian Flora of Podolia; Institute of Geological Science, Academy of Science of the Ukrainian SSR: Kiev, Ukrainian, 1975; pp. 1–80. [Google Scholar]
- Senkevich, M.A. Fossil plants in the Tokrau horizon of the Upper Silurian. In The Tokrau Horizon of the Upper Silurian Series: Balkhash Segment; Nikitin, I.F., Bandaletoc, S.M., Eds.; Nauka, Alma-Ata: Almaty, Kazakhstan, 1986. (In Russian) [Google Scholar]
- Senkevich, M.A. New Devonian psilophytes from Kazakhstan. Esheg Vses Paleontol Obs. 1975, 21, 288–298. (In Russian) [Google Scholar]
- Cai, C.Y.; Dou, Y.W.; Edwards, D. New observations on a Přídolí plant assemblage from north Xinjing, northwest China, with comments on its evolutionary and palaeogeographical significance. Geol. Mag. 1993, 130, 155–170. [Google Scholar]
- Gonez, P.; Huu, H.N.; Hoa, P.T.; Clément, G.; Janvire, P. The oldest flora of the South China Block, and the stratigraphic bearings of the plant remains from the Ngoc Vung Series, northern Vietnam. J. Asian Earth Sci. 2012, 43, 51–63. [Google Scholar] [CrossRef]
- Edwards, D.; Feehan, J.; Smith, D.G. A late Wenlock flora from Co. Tipperary, Ireland. Bot. J. Linn. Soc. 1983, 86, 19–36. [Google Scholar] [CrossRef]
- Rogerson, E.C.W.; Edwards, D.; Davies, K.L.; Richardson, J.B. Identification of in situ spores in a Silurian Cooksonia from the Welsh Borderland. In Studies in Palaeobotany and Palynology in Honour of Professor W. G. Chaloner, F.R.S.; Collinson, M.E., Scott, A.C., Eds.; Special Papers in Palaeontology; Palaeontological Association: Durham, UK, 1993; Volume 49, pp. 17–30. [Google Scholar]
- Daber, R. Cooksonia—One of the most ancient psilophytes—Widely distributed, but rare. Botanique 1971, 2, 35–40. [Google Scholar]
- Morel, E.; Edwards, D.; Inigez Rodriguez, M. The first record of Cooksonia from South America in Silurian rocks of Bolivia. Geol. Mag. 1995, 132, 449–452. [Google Scholar] [CrossRef]
- Lang, W.H. On plant-remains from the Downtonian of England and Wales. Philos. Trans. R. Soc. Lond. 1937, B227, 245–291. [Google Scholar]
- Tims, J.D.J. The Early Land Flora of Victoria. Ph.D. Thesis, University of Melbourne, Melbourne, Australia, 1980. [Google Scholar]
- Dou, Y.W.; Sun, Z.H. Devonian Plants. In Palaeontological Atlas of Xinjiang, Vol II, Late Palaeozoic Section; Geological Publishing House: Beijing, China, 1983. (In Chinese) [Google Scholar]
- Steemans, P.; Petus, E.; Breuer, P.; Mauller-Mendlowicz, P.; Gerrienne, P. Palaeozoic innovations in the micro and megafossil plant record from the earliest plant spores to the earliest seeds. In Earth and life: Global Biodiversity, Extinction Intervals and Biogeographic Perturbations through Time; Talent, J.A., Ed.; International Year of Planet Eart; Springer Science: Dordrecht, The Netherlands, 2012; pp. 437–477. [Google Scholar]
- Mehlqvist, K.; Vajda, V.; Steemans, P. Early land plant spore assemblages from the Late Silurian of Skåne, Sweden. GFF 2012, 134, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Gray, J. The microfossil record of early land plants: Advances in understanding early terrestrialization, 1870–1984. Philos. Trans. Royal. Soc. Lond. Ser. B 1985, 309, 167–195. [Google Scholar]
- Morris, J.L.; Wright, V.P.; Edwards, D. Siluro-Devonian landscapes of southern Britain: The stability and nature of early vascular plant habitats. J. Geol. Soc. 2012, 169, 173–190. [Google Scholar] [CrossRef]
- Gray, J. Major Paleozoic land plant evolutionary bio-events. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1993, 104, 153–169. [Google Scholar] [CrossRef]
- Wellman, C.H. Origin, function and development of the spore wall in early land plants. In The Evolution of Plant Physiology; Hemsleyx, A.R., Poole, I., Eds.; Academic Press: Cambridge, MA, USA, 2004; pp. 43–63. [Google Scholar]
- Mogensen, G.S. The biological significance of morphological characters in bryophytes: The spore. Bryologist 1981, 84, 187–207. [Google Scholar] [CrossRef]
- Allen, K.C. A review of in situ late Silurian and Devonian spores. Rev. Palaeobot. Palynol. 1980, 29, 253–270. [Google Scholar] [CrossRef]
- Balme, B.A. Fossil in situ spores and pollen grains: An annotated catalogue. Rev. Palaeobot. Palynol. 1995, 87, 81–323. [Google Scholar] [CrossRef]
- Stemans, P. Miospore evolution from the Ordovician to the Silurian. Rev. Palaeobot. Palynol. 2000, 113, 189–196. [Google Scholar] [CrossRef]
- Wellman, C.H.; Steemans, P.; Vecoli, M. Palaeophytogeography of Ordovician-Silurian Land Plants. Geol. Soc. Lond. Mem. 2014, 38, 461–476. [Google Scholar] [CrossRef]
- Brown, R.C.; Lemmon, B.E.; Shimamura, M.; Villarreal, J.C.; Renzaglia, K.S. Spores of relictual bryophytes: Diverse adaptations to life on land. Rev. Palaeobot. Palynol. 2015, 216, 1–17. [Google Scholar] [CrossRef]
- Mehlqvist, K.; Steemans, P.; Vajda, V. First evidence of Devonian strata in Sweden—A palynological investigation of Övedskloster drillcores 1 and 2, Skåne, Sweden. Rev. Palaeobot. Palynol. 2015, 221, 144–159. [Google Scholar] [CrossRef] [Green Version]
- Hao, S.; Xue, J. The early Devonian Posongchong flora of Yunnan: A Contribution to an Understanding of the Evolution and Early Diversification of Vascular Plants; Science Press: Beijing, China, 2013; pp. 1–366. [Google Scholar]
- Harris, B.J.; Harrison, C.J.; Hetherington, A.M.; Williams, T.A. Phylogenomic Evidence for the Monophyly of Bryophytes and the Reductive Evolution of Stomata. Curr. Biol. 2020, 30, P2001-20012E2. [Google Scholar] [CrossRef]
- Lehnert, O.; Frýda, J.; Joachimski, M.; Meinhold, G.; Čáp, P. A latest Silurian Supergreenhouse: The trigger for the Pridoli Transgrediens Extinction Event. In Proceedings of the 34th International Geological Congress, Brisbane, Australia, 5–10 August 2012; p. 3255. [Google Scholar]
- Qu, Q.M.; Zhu, M.; Zhao, W.J. Silurian atmospheric O2 changes and the early radiation of gnathostomes. Palaeoworld 2010, 19, 146–159. [Google Scholar] [CrossRef]
- Kerp, H.; Hass, H. De Onder-Devonische Rhynie Chert—Het oudste en meest compleet bewaard gebleven terrestriche. Grondboor Hamer 2004, 58, 33–50. [Google Scholar]
- Taylor, T.N.; Kerp, H.; Hass, H. Life history biology of early land plants: Deciphering the gametophyte phase. Proc. Natl. Acad. Sci. USA 2005, 102, 5892–5897. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Pri-Tal, O.; Michaeli, D.; Mosquna, A. Evolution of Abscisic Acid Signaling Module and Its Perception. Front. Plant Sci. 2020, 11, 934. [Google Scholar] [CrossRef] [PubMed]
- Terada, S.; Kubo, M.; Akiyoshi, N.; Sano, R.; Nomura, T.; Sawa, S.; Ohtani, M.; Demura, T. Expression of peat moss VASCULAR RELATED NAC-DOMAIN homologs in Nicotiana benthamiana leaf cells induces ectopic secondary wall formation. Plant Mol. Biol. 2021, 106, 309–317. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | Envelope Enclosed Monads | Envelope Enclosed Dyads | Envelope Enclosed Tetrad | Naked Monads | Naked Dyads | Naked Tetrads | Hilate Spores | Trilete Spores |
|---|---|---|---|---|---|---|---|---|
| Přídolí | █ █ | █ █ | █ █ | ██ ██ | █ █ | █ █ | ███ ███ | ███ ███ |
| Ludlow | █ █ | █ █ | █ █ | ██ ██ | █ █ | █ █ | ██ ██ | ██ ██ |
| Wenlock | █ █ | █ █ | █ █ | █ █ | ██ ██ | ██ ██ | ██ ██ | ██ ██ |
| Llandovery | █ █ | █ █ | █ █ | █ █ | ██ ██ | ██ ██ | █ █ | █ █ |
| ██ ██ | ███ ███ | ███ ███ | █ █ | ███ ███ | ███ ███ | █ █ | █ █ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pšenička, J.; Bek, J.; Frýda, J.; Žárský, V.; Uhlířová, M.; Štorch, P. Dynamics of Silurian Plants as Response to Climate Changes. Life 2021, 11, 906. https://doi.org/10.3390/life11090906
Pšenička J, Bek J, Frýda J, Žárský V, Uhlířová M, Štorch P. Dynamics of Silurian Plants as Response to Climate Changes. Life. 2021; 11(9):906. https://doi.org/10.3390/life11090906
Chicago/Turabian StylePšenička, Josef, Jiří Bek, Jiří Frýda, Viktor Žárský, Monika Uhlířová, and Petr Štorch. 2021. "Dynamics of Silurian Plants as Response to Climate Changes" Life 11, no. 9: 906. https://doi.org/10.3390/life11090906
APA StylePšenička, J., Bek, J., Frýda, J., Žárský, V., Uhlířová, M., & Štorch, P. (2021). Dynamics of Silurian Plants as Response to Climate Changes. Life, 11(9), 906. https://doi.org/10.3390/life11090906