


Dietary Polyphenols as Natural Inhibitors of α-Amylase and α-Glucosidase


Abstract
:1. Introduction
2. Dietary Polyphenols as Antidiabetic Agents
3. Digestion of Carbohydrates: Role of α-Amylase and α-Glucosidase
4. Structure and Activity of α-Amylase
5. Structure–Activity Relationship of Polyphenols Inhibiting α-Amylase
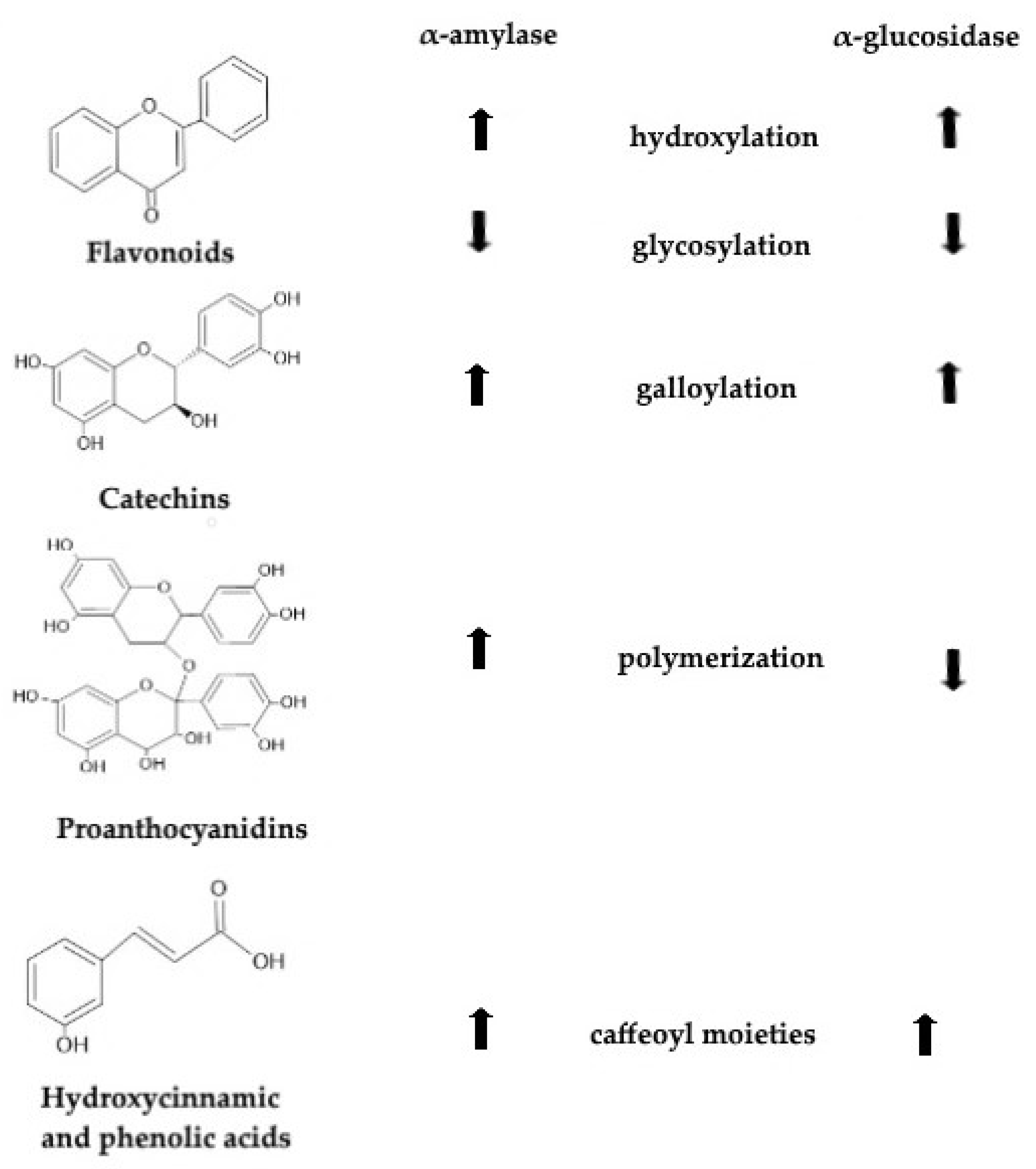
5.1. Flavonoids
5.1.1. Methylation and Methoxylation of Flavonoids
5.1.2. Hydroxylation of Flavonoids
5.1.3. Glycosylation of Flavonoids
5.1.4. Hydrogenation of the C2=C3 Double Bond of Flavonoids
5.2. Catechins
5.3. Proanthocyanidins and Anthocyanidins
5.4. Tannins
5.5. Hydroxycinnamic Acids and Phenolic Acids
6. Structure and Activity of α-Glucosidase
7. Structure–Activity Relationship of Polyphenols Inhibiting α-Glucosidase
7.1. Flavonoids
7.1.1. Hydroxylation of Flavonoids
7.1.2. Methylation and Methoxylation of Flavonoids
7.1.3. Glycosylation of Flavonoids
7.1.4. Hydrogenation of the C2=C3 Double Bond of Flavonoids
7.2. Catechins
7.3. Proanthocyanidins and Anthocyanidins
7.4. Stilbenes
7.5. Hydroxycinnamic Acids
7.6. Tannins
7.7. Chalcones
8. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tinajero, M.G.; Malik, V.S. An Update on the Epidemiology of Type 2 Diabetes: A Global Perspective. Endocrinol. Metab. Clin. N. Am. 2021, 50, 337–355. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.; Bain, S.; Kanamarlapudi, V. A review of Current Trends with Type 2 Diabetes Epidemiology, Aetiology, Pathogenesis, Treatments and Future Perspectives. Diabetes Metab. Syndr. Obes. 2021, 14, 3567–3602. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.; Hasan, M.K.; Neaz, S.; Hussain, N.; Hossain, M.F.; Rahman, T. Diabetes Mellitus: Insights from Epidemiology, Biochemistry, Risk Factors, Diagnosis, Complications and Comprehensive Management. Diabetology 2021, 2, 36–50. [Google Scholar] [CrossRef]
- International Diabetes Federation. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021; Available online: https://www.diabetesatlas.org (accessed on 17 October 2022).
- He, K.; Shi, J.C.; Mao, X.M. Safety and efficacy of acarbose in the treatment of diabetes in Chinese patients. Ther. Clin. Risk Manag. 2014, 10, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Zhao, C.; Capanoglu Guven, E.; Paoli, P.; Simal-Gandara, J.; Ramkumar, K.M.; Wang, S.; Buleu, F.; Pah, A.; Turi, V.; et al. Dietary polyphenols as antidiabetic agents. Front. Nutr. 2020, 1, 18–44. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Miao, M. Dietary polyphenols modulate starch digestion and glycaemic level: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 541–555. [Google Scholar] [CrossRef]
- Modak, M.; Dixit, P.; Londhe, J.; Ghaskadbi, S.; Devasagayam, T.P.A. Indian herbs and herbal drugs used for the treatment of diabetes. J. Clin. Biochem. Nutr. 2007, 40, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, C.; Wang, G.; Cao, J.; Yang, X.; Liu, X.; Sun, L. α-Amylase inhibition of a certain dietary polyphenol is predominantly affected by the concentration of α-1.4-glucosidic bonds in starchy and artificial substrates. Food Res. Int. 2022, 157, 111210. [Google Scholar] [CrossRef]
- Vinson, J. Investigation of an amylase inhibitor on human glucose absorption after starch consumption. Open Nutraceuticals J. 2009, 2, 88–91. [Google Scholar] [CrossRef] [Green Version]
- Bedekar, A.; Shah, K.; Koffas, M. Natural products for type 2 diabetes treatment. Adv. Appl. Microbiol. 2010, 71, 21–73. [Google Scholar]
- Sharma, R. Enzyme Inhibition and Bioapplications; IntechOpen: Rijeka, Croatia, 2012. [Google Scholar]
- Nelson, D.; Cox, M. Enzyme Kinetics as an Approach to Understanding Mechanism. In Lehninger Principles of Biochemistry, 4th ed.; Freeman, W.H., Ed.; Macmillan: New York, NY, USA, 2005. [Google Scholar]
- Copeland, R. Evaluation of Enzyme Inhibitors in Drug Discovery; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013. [Google Scholar]
- Di Stefano, E.; Oliviero, T.; Udenigwe, C.C. Functional significance and structure-activity relationship of food-derived α-glucosidase inhibitors. Curr. Opin. Food Sci. 2018, 20, 7–12. [Google Scholar] [CrossRef]
- Şöhretoğlu, D.; Sari, S.; Barut, B.; Özel, A. Discovery of potent α-glucosidase inhibitor flavonols: Insights into mechanism of action through inhibition kinetics and docking simulations. Bioorg. Chem. 2018, 79, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Hu, M.; Hu, X.; Ding, H.; Gong, D.; Zhang, G. Inhibitory mechanism of epicatechin gallate on α-amylase and α-glucosidase and its combinational effect with acarbose or epigallocatechin gallate. J. Mol. Liq. 2019, 290, 111202. [Google Scholar] [CrossRef]
- Aleixandre, A.; Gil, J.V.; Sineiro, J.; Rosell, C.M. Understanding phenolic acids inhibition of α-amylase and α-glucosidase and influence of reaction conditions. Food Chem. 2022, 372, 131231. [Google Scholar] [CrossRef]
- Fraga, C.G.; Galleano, M.; Verstraeten, S.V.; Oteiza, P.I. Basic biochemical mechanisms behind the health benefits of polyphenols. Mol. Asp. Med. 2010, 31, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Leri, M.; Scuto, M.; Ontario, M.L.; Calabrese, V.; Calabrese, E.J.; Bucciantini, M.; Stefani, M. Healthy Effects of Plant Polyphenols: Molecular Mechanisms. Int. J. Mol. Sci. 2020, 21, 1250. [Google Scholar] [CrossRef] [Green Version]
- Mithul Aravind, S.; Wichienchot, S.; Tsao, R.; Ramakrishnan, S.; Chakkaravarthi, S. Role of dietary polyphenols on gut microbiota, their metabolites and health benefits. Food Res. Int. 2021, 42, 110189. [Google Scholar] [CrossRef]
- Grosso, G.; Godos, J.; Currenti, W.; Micek, A.; Falzone, L.; Libra, M.; Giampieri, F.; Forbes-Hernández, T.Y.; Quiles, J.L.; Battino, M.; et al. The Effect of Dietary Polyphenols on Vascular Health and Hypertension: Current Evidence and Mechanisms of Action. Nutrients 2022, 14, 545. [Google Scholar] [CrossRef]
- Rana, A.; Samtiya, M.; Dhewa, T.; Mishra, V.; Aluko, R.E. Health benefits of polyphenols: A concise review. J. Food Biochem. 2022, 46, 14264. [Google Scholar] [CrossRef]
- Xie, Y.X.; Xiao, J.B.; Kai, G.Y.; Chen, X.Q. Glycation of plasma proteins in type II diabetes lowers the non-covalent interaction affinities for dietary polyphenols. Integr. Biol. 2012, 4, 502–507. [Google Scholar] [CrossRef]
- Wang, Y.L.; Zhao, Y.; Yang, F.; Yuan, Y.M.; Wang, H.; Xiao, J.B. Influences of glucose on the dietary hydroxyflavonoids-plasma proteins interaction. J. Agric. Food Chem. 2012, 60, 12116–12221. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Shao, R.; Xiao, J. Is There Consistency between the Binding Affinity and Inhibitory Potential of Natural Polyphenols as α-amylase Inhibitors? Crit. Rev. Food Sci. Nutr. 2016, 56, 1630–1639. [Google Scholar] [CrossRef]
- Gundala, N.K.V.; Naidu, V.G.M.; Das, U.N. Arachidonic acid and lipoxin A4 attenuate alloxan-induced cytotoxicity to RIN5F cells in vitro and type 1 diabetes mellitus in vivo. BioFactors 2016, 43, 251–271. [Google Scholar] [CrossRef] [PubMed]
- Viswanath, B.; Choi, C.S.; Lee, K.; Kim, S. Recent trends in the development of diagnostic tools for diabetes mellitus using patient saliva. Trends Anal. Chem. 2017, 89, 60–67. [Google Scholar] [CrossRef]
- Sales, P.M.; Souza, P.M.; Simeoni, L.A.; Magalhães, P.O.; Silveira, D. α-Amylase inhibitors: A review of raw material and isolated compounds from plant source. J. Pharm. Pharm. Sci. 2012, 15, 141–183. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Warren, F.J.; Gidley, M.J. Natural products for glycaemic control: Polyphenols as inhibitors of alpha-amylase. Trends Food Sci. Technol. 2019, 91, 262–273. [Google Scholar] [CrossRef]
- Maideen, N.M.P. Pharmacologically relevant drug interactions of α-glucosidase inhibitors. J. Diabetes Metab. Disord. Control 2019, 6, 28–30. [Google Scholar] [CrossRef]
- Wang, L.; Voss, E.A.; Weaver, J.; Hester, L.; Yuan, Z.; DeFalco, F.; Schuemie, M.J.; Ryan, P.B.; Sun, D.; Freedman, A.; et al. Diabetic ketoacidosis in patients with type 2 diabetes treated with sodium glucose co-transporter 2 inhibitors versus other antihyperglycemic agents: An observational study of four US administrative claims databases. Pharmacoepidemiol. Drug Saf. 2019, 28, 1620–1628. [Google Scholar] [CrossRef]
- Rosak, C.; Mertes, G. Critical evaluation of the role of acarbose in the treatment of diabetes: Patient considerations. Diabetes Metab. Syndr. Obes. 2012, 5, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Bahadoran, Z.; Mirmiran, P.; Azizi, F. Dietary polyphenols as potential nutraceuticals in management of diabetes: A review. J. Diabetes Metab. Disord. 2013, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Heidari, R.; Zareae, S.; Heidarizadeh, M. Extraction, purification, and inhibitory effect of alpha-amylase inhibitor from wheat (Triticum aestivum Var. Zarrin). Pak. J. Nutr. 2005, 4, 101–105. [Google Scholar]
- Al Kazaz, M.; Desseaux, V.; Marchis-Mouren, G.; Prodanov, E.; Santimone, M. The mechanism of porcine pancreatic alpha-amylase. Inhibition of maltopentaose hydrolysis by acarbose, maltose and maltotriose. Eur. J. Biochem. 1998, 252, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Lee, S.B.; Lee, H.S.; Lee, S.Y.; Baek, J.S.; Kim, D.; Moon, T.W.; Robyt, J.F.; Park, K.H. Comparative study of the inhibition of alpha-glucosidase, alpha-amylase, and cyclomaltodextrin glucanosyltransferase by acarbose, isoacarbose, and acarviosineglucose. Arch. Biochem. Biophys. 1999, 371, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Junge, B.; Frommer, W.; Schmidt, D.; Truscheit, E. Enzyme Inhibitors; Brodbeck, U., Ed.; Berlag Chemie: Weinheim, Germany, 1980. [Google Scholar]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, F.; Khadija, J.F.; Islam, R.; Shohag, S.; Mitra, S.; Alghamdi, S.; Babalghith, A.O.; Theyab, A.; Rahman, M.T.; Akter, A.; et al. Investigating Polyphenol Nanoformulations for Therapeutic Targets against Diabetes Mellitus. Hindawi 2022, 2022, 5649156. [Google Scholar] [CrossRef] [PubMed]
- Kesharwani, P.; Gorain, B.; Low, S.L.; Tan, S.A.; Ling, E.C.S.; Lim, Y.K.; Chin, C.M.; Lee, P.Y.; Lee, C.M.M.; Ooi, C.H.; et al. Nanotechnology based approaches for anti-diabetic drugs delivery. Diabetes Res. Clin. Pract. 2018, 136, 52–77. [Google Scholar] [CrossRef]
- Yang, B.; Dong, Y.; Wang, F.; Zhang, Y. Nanoformulations to Enhance the Bioavailability and Physiological Functions of Polyphenols. Molecules 2020, 25, 4613. [Google Scholar] [CrossRef]
- Mezzenga, R.; Schurtenberger, P.; Burbidge, A.; Michel, M. Understanding foods as soft materials. Nat. Mater. 2005, 4, 729–740. [Google Scholar] [CrossRef]
- Ubbink, J.; Burbidge, A.; Mezzenga, R. Food structure and functionality: A soft matter perspective. Soft Matter 2008, 4, 1569–1581. [Google Scholar] [CrossRef]
- Des Rieux, A.; Fievez, V.; Garinot, M.; Schneider, Y.J.; Preat, V.J. Nanoparticles as potential oral delivery systems of proteins and vaccines: A mechanistic approach. J. Control. Release 2006, 116, 1–27. [Google Scholar] [CrossRef]
- Hu, B.; Liu, X.; Zhang, C.; Zeng, X. Food macromolecule based nanodelivery systems for enhancing the bioavailability of polyphenols. J. Food Drug Anal. 2017, 25, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurell, R.F.; Finot, P.A. Nutritional consequences of the reactions between proteins and oxidised polyphenolic acids. Adv. Exp. Med. Biol. 1984, 177, 423–435. [Google Scholar]
- Yoon, S.H.; Robyt, J.F. Addition of maltodextrins to the nonreducingend of acarbose by reaction of acarbose with cyclomaltohexaose and cyclomaltodextrin glucanyltransferase. Carbohydr. Res. 2002, 337, 509–516. [Google Scholar] [CrossRef]
- Moran, J.I.; Cyras, V.P.; Vazquez, A.; Foresti, M.L. Characterization of chemically modified potato starch films through enzymatic degradation. J. Polym. Environ. 2011, 19, 217–224. [Google Scholar] [CrossRef]
- Obiro, W. The nutraceutical role of the Phaseolus vulgaris alpha amylase inhibitor. Braz. J. Nutr. 2008, 100, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Warren, F.J.; Zhang, B.; Waltzer, G.; Gidley, M.J.; Dhital, S. The interplay of α-amylase and amyloglucosidase activities on the digestion of starch in in vitro enzymic systems. Carbohydr. Polym. 2015, 117, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Radulian, G.; Rusu, E.; Dragomir, A.; Posea, M. Metabolic effects of low glycaemic index diets. Nutr. J. 2009, 8, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.J.; Gidley, M.J.; Warren, F.J. Tea polyphenols enhance binding of porcine pancreatic alpha-amylase with starch granules but reduce catalytic activity. Food Chem. 2018, 258, 164–173. [Google Scholar] [CrossRef]
- Chai, Y.; Wang, M.; Zhang, G. Interaction between Amylose and Tea Polyphenols Modulates the Postprandial Glycemic Response to High-Amylose Maize Starch. J. Agric. Food Chem. 2013, 61, 8608–8615. [Google Scholar] [CrossRef]
- Obiro, W.C.; Ray, S.S.; Emmambux, M.N. V-amylose Structural Characteristics, Methods of Preparation, Significance, and Potential Applications. Food Rev. Int. 2012, 28, 412–438. [Google Scholar] [CrossRef] [Green Version]
- Tang, P.; Liu, Y.; Gao, Y.; Wang, Y.; Zhang, H.; Liu, Y.; Wu, D. Molecular simulation, characteristics and mechanism of thermal-responsive acetylated amylose V-type helical complexes. J. Mater. Chem. B 2021, 9, 3389. [Google Scholar] [CrossRef] [PubMed]
- Amoako, D.B.; Awika, J.M. Resistant starch formation through intrahelical V-complexes between polymeric proanthocyanidins and amylose. Food Chem. 2019, 285, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Svensson, B. Regional distant sequence homology between amylases, alpha-glucosidases and transglucanosylases. FEBS Lett. 1998, 230, 72–76. [Google Scholar] [CrossRef]
- Chakraborty, R.; Rajagopalan, R. Diabetes and insulin resistance associated disorders: Disease and the therapy. Curr. Sci. 2002, 83, 1533–1538. [Google Scholar]
- Mahmood, N. A review of α-amylase inhibitors on weight loss and glycemic control in pathological state such as obesity and diabetes. Comp. Clin. Path. 2016, 25, 1253–1264. [Google Scholar] [CrossRef]
- Gumucio, D.L.; Wiebauer, K.; Caldwell, R.M.; Samuelson, L.C.; Meisler, M.H. Concerted evolution of human amylase genes. Mol. Cell. Biol. 1988, 8, 1197–1205. [Google Scholar] [PubMed] [Green Version]
- Wattanathorn, J.; Kawvised, S.; Thukhammee, W. Encapsulated Mulberry Fruit Extract Alleviates Changes in an Animal Model of Menopause with Metabolic Syndrome. Oxid. Med. Cell. Longev. 2019, 2019, 5360560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varghese, L.N.; Mehrotra, N. α-amylase inhibitory activity of microencapsulated Nigella sativa L. and herb-drug interaction: An in vitro analysis. Ann. Phytomed. 2020, 9, 107–112. [Google Scholar] [CrossRef]
- Safitri, A.; Roosdiana, A.; Kurnianingsih, N.; Fatchiyah, F.; Mayasari, E.; Rachamawati, R. Microencapsulation of Ruellia tuberosa L. Aqueous Root Extracts Using Chitosan-Sodium Tripolyphosphate and Their In Vitro Biological Activities. Scientifica 2022, 2022, 9522463. [Google Scholar] [CrossRef]
- Daza, L.D.; Fujita, A.; Granato, D.; Fávaro-Trindade, C.S.; Genovese, M.I. Functional properties of encapsulated Cagaita (Eugenia dysenterica DC.) fruit extract. Food Biosci. 2017, 18, 15–21. [Google Scholar] [CrossRef]
- De Souza, V.B.; Thomazini, M.; Echalar Barrientos, M.A.; Nalin, C.M.; Ferro-Furtado, R.; Genovese, M.I.; Favaro-Trindade, C.S. Functional properties and encapsulation of a proanthocyanidin-rich cinnamon extract (Cinnamomum zeylanicum) by complex coacervation using gelatin and different polysaccharides. Food Hydrocoll. 2018, 77, 297–306. [Google Scholar] [CrossRef]
- Safitri, A.; Roosdiana, A.; Hitdatania, E.; Damayanti, S.A. In Vitro Alpha-Amylase Inhibitory Activtiy of Microencapsulated Cosmos caudatus Kunth Extracts. Indones. J. Chem. 2022, 22, 212–222. [Google Scholar] [CrossRef]
- Zorzenon, M.R.T.; Formigoni, M.; da Silva, S.B.; Hodas, F.; Piovan, S.; Ciotta, S.R.; Jansen, C.A.; Dacome, A.S.; Pilau, E.J.; Mareze-Costa, C.E.; et al. Spray drying encapsulation of stevia extract with maltodextrin and evaluation of the physicochemical and functional properties of produced powders. J. Food Sci. 2020, 85, 3590–3600. [Google Scholar] [CrossRef] [PubMed]
- Lavelli, V.; Pedapati, S.C.; Harsha, S.; Laureati, M.; Pagliarini, E. Degradation kinetics of encapsulated grape skin phenolics and micronized grape skins in various water activity environments to improve wide-ranging and tailormade food applications. Innov. Food Sci. Emerg. Technol. 2016, 39, 156–164. [Google Scholar] [CrossRef]
- Buljeta, I.; Nosić, M.; Pichler, A.; Ivić, I.; Šimunović, J.; Kopjar, M. Apple Fibers as Carriers of Blackberry Juice Polyphenols: Development of Natural Functional Food Additives. Molecules 2022, 27, 3029. [Google Scholar] [CrossRef]
- Pasukamonset, P.; Kwon, O.; Adisakwattana, S. Alginate-based encapsulation of polyphenols from Clitoria ternatea petal flower extract enhances stability and biological activity under simulated gastrointestinal conditions. Food Hydrocoll. 2016, 61, 772–779. [Google Scholar] [CrossRef]
- Vukoja, J.; Buljeta, I.; Pichler, A.; Šimunović, J.; Kopjar, M. Formulation and Stability of Cellulose-Based Delivery Systems of Raspberry Phenolics. Processes 2021, 9, 90. [Google Scholar] [CrossRef]
- Vasile, M.A.; Milea, S.A.; Enachi, E.; Barbu, V.; Circiumaru, A.; Bahrim, G.E.; Rapeanu, G.; Stanciuc, N. Functional Enhancement of Bioactives from Black Beans and Lactic Acid Bacteria into an Innovative Food Ingredient by Comicroencapsulation. Food Bioprocess Technol. 2020, 13, 978–987. [Google Scholar] [CrossRef]
- Sylla, N.; Bouyahya, A.; Taha, D.; Dakka, N.; Elhajji, H. Study of the antioxidant and antidiabetic activity in vitro of free and encapsulated phenolic compounds of olive pomace. Biocatal. Agric. Biotechnol. 2021, 36, 102126. [Google Scholar] [CrossRef]
- Wang, L.; Clardy, A.; Hui, D.; Wu, Y. Physicochemical properties of encapsulated bitter melon juice using spray drying. Bioact. Carbohydr. Diet. Fibre 2021, 26, 100278. [Google Scholar] [CrossRef]
- Wu, G.; Hui, X.; Mu, J.; Brennan, M.; Brennan, C.S. Functionalization of whey protein isolate fortified with blackcurrant concentrate by spray-drying and freeze-drying strategies. Food Res. Int. 2021, 141, 110025. [Google Scholar] [CrossRef] [PubMed]
- Keskin, M. Chemical characterization of arabic gum-chitosan-propolis beads and determination of α-amylase inhibition effect. Prog. Nutr. 2020, 22, 562–567. [Google Scholar] [CrossRef]
- Ćorković, I.; Pichler, A.; Buljeta, I.; Šimunović, J.; Kopjar, M. Carboxymethylcellulose hydrogels: Effect of its different amount on preservation of tart cherry anthocyanins and polyphenols. Curr. Plant Biol. 2021, 28, 100222. [Google Scholar] [CrossRef]
- Akolade, J.O.; Oloyede, H.O.B.; Onyenekwe, P.C. Encapsulation in chitosan-based polyelectrolyte complexes enhances antidiabetic activity of curcumin. J. Funct. Foods 2017, 35, 584–594. [Google Scholar] [CrossRef]
- Mel, M.M.R.D.; Gunathilake, K.D.P.P.; Fernando, C.A.N. Formulation of microencapsulated rutin and evaluation of bioactivity and stability upon in vitro digestive and dialysis conditions. Int. J. Biol. Macromol. 2020, 159, 316–323. [Google Scholar] [CrossRef]
- Lo Piparo, E.; Scheib, H.; Frei, N.; Williamson, G.; Grigorov, M.; Chou, C.J. Flavonoids for Controlling Starch Digestion: Structural Requirements for Inhibiting Human α-Amylase. J. Med. Chem. 2008, 51, 3555–3561. [Google Scholar] [CrossRef]
- Xiao, J.; Ni, X.; Kai, G.; Chen, X. A Review on Structure-Activity Relationship of Dietary Polyphenols Inhibiting α-amylase. Crit. Rev. Food Sci. Nutr. 2013, 53, 497–506. [Google Scholar] [CrossRef]
- Gong, L.; Feng, D.; Wang, T.; Ren, Y.; Liu, Y.; Wang, J. Inhibitors of α-amylase and α-glucosidase: Potential linkage for whole cereal foods on prevention of hyperglycemia. Food Sci. Nutr. 2020, 8, 6320–6337. [Google Scholar] [CrossRef]
- Xiao, J.; Kai, G.; Ni, X.; Yang, F.; Chen, X. Interaction of natural polyphenols with α-amylase in vitro: Molecular property-affinity relationship aspect. Mol. BioSyst. 2011, 7, 1883–1890. [Google Scholar] [CrossRef]
- Rohn, S.; Rawel, H.M.; Kroll, J. Inhibitory Effects of Plant Phenols on the Activity of Selected Enzymes. J. Agric. Food Chem. 2002, 50, 3566–3571. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, Y.; Zhou, W. In vitro and in silico studies of the inhibition activity of anthocyanins against porcine pancreatic α-amylase. J. Funct. Foods 2016, 21, 50–57. [Google Scholar] [CrossRef]
- Boath, A.S.; Grussu, D.; Stewart, D.; McDougall, G.J. Berry Polyphenols Inhibit Digestive Enzymes: A Source of Potential Health Benefits? Food Dig. 2012, 3, 1–7. [Google Scholar] [CrossRef]
- Kaeswurm, J.A.H.; Claasen, B.; Fischer, M.; Buchweitz, M. Interaction of Structurally Diverse Phenolic Compounds with Porcine Pancreatic α-amylase. J. Agric. Food Chem. 2019, 67, 11108–11118. [Google Scholar] [CrossRef] [PubMed]
- Moremen, K.W.; Trimble, R.B.; Herscovics, A. Glycosidases of the asparagine-linked oligosaccharide processing pathway. Glycobiology 1994, 4, 113–125. [Google Scholar] [CrossRef]
- Terra, W.R.; Ferreira, C. Biochemistry of digestion. In Comprehensive Molecular Insect Science; Lawrence, I.G., Ed.; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Auiewiriyanukul, W.; Saburi, W.; Kato, K.; Yao, M.; Mori, H. Function and structure of GH13_31 α-glucosidase with high α-(1→4)-glucosidic linkage specificity and transglucosylation activity. FEBS Lett. 2018, 592, 2268–2281. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S. Molecular Mechanism in α-Glucosidase and Glucoamylase. Biosci. Biotechnol. Biochem. 1997, 61, 1233–1239. [Google Scholar] [CrossRef]
- Martinez-Gonzalez, A.I.; Díaz-Sánchez, A.G.; de la Rosa, L.A.; Vargas-Requena, C.L.; Bustos-Jaimes, I.; Alvarez-Parrilla, E. Polyphenolic Compounds and Digestive Enzymes: In Vitro Non-Covalent Interactions. Molecules 2017, 22, 669. [Google Scholar] [CrossRef] [Green Version]
- Zhai, X.; Wu, K.; Ji, R.; Zhao, Y.; Lu, J.; Yu, Z.; Xu, X.; Huang, J. Structure and Function Insight of the α-Glucosidase QsGH13 from Qipengyuania seohaensis sp. SW-135. Front. Microbiol. 2022, 13, 849585. [Google Scholar] [CrossRef]
- Vulić, J.; Šeregelj, V.; Kalušević, A.; Lević, S.; Nedović, V.; Tumbas Šaponjac, V.; Čanadanović-Brunet, J.; Ćetković, G. Bioavailability and Bioactivity of Encapsulated Phenolics and Carotenoids Isolated from Red Pepper Waste. Molecules 2019, 24, 2837. [Google Scholar] [CrossRef] [Green Version]
- Kopjar, M.; Ćorković, I.; Buljeta, I.; Šimunović, J.; Pichler, A. Fortification of Pectin/Blackberry Hydrogels with Apple Fibers: Effect on Phenolics, Antioxidant Activity and Inhibition of α-Glucosidase. Antioxidants 2022, 11, 1459. [Google Scholar] [CrossRef]
- Ahmad, M.; Mudgil, P.; Gani, A.; Hamed, F.; Masoodi, F.A.; Maqsood, S. Nanoencapsulation of catechin in starch nanoparticles: Characterization; release behavior and bioactivity retention during in-vitro digestion. Food Chem. 2018, 270, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chena, C.; Zhang, B.; Huang, Q. The inhibitory effects of flavonoids on α-amylase and α-glucosidase. Crit. Rev. Food Sci. Nutr. 2019, 60, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Kai, G.; Yamamoto, K.; Chen, X. Advance in Dietary Polyphenols as α-Glucosidases Inhibitors: A Review on Structure-Activity Relationship Aspect. Crit. Rev. Food Sci. Nutr. 2013, 53, 818–836. [Google Scholar] [CrossRef] [PubMed]
- McDougall, G.J.; Stewart, D. The inhibitory effects of berry polyphenols on digestive enzymes. BioFactors 2005, 23, 189–195. [Google Scholar] [CrossRef]
- McDougall, G.J.; Shpiro, F.; Dobson, P.; Smith, P.; Blake, A.; Stewart, D. Different Polyphenolic Components of Soft Fruits Inhibit α-amylase and α-glucosidase. J. Agric. Food Chem. 2005, 53, 2760–2766. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, H.Y.; Jin, C.H. Mechanistic investigation of anthocyanidin derivates as α-glucosidase inhibitors. Bioorg. Chem. 2019, 87, 803–809. [Google Scholar] [CrossRef]


{kind=link}
| Polyphenols from Plant Material | ||||
|---|---|---|---|---|
| Source | Individual Polyphenols | Delivery System | Inhibitory Activity | Ref. |
| Mulberry fruit (Morus alba) extract | Cyanidin-3-glucoside, rutin and gallic acid | Freeze-dried particles prepared from maltodextrin | IC50 value for mulberry fruit extract was 0.30 mg/mL and for encapsulated mulberry fruit extract it was 0.28 mg/mL | [62] |
| Black Caraway (Nigella sativa) seeds | n.d. | Extract microencapsulated in β-cyclodextrin | For concentrations of extracts from 10 µg/mL to 160 µg/mL, inhibitory activity (%) ranged from 16.9% to 32.31% | [63] |
| Minnieroot (Ruellia tuberosa) root | n.d. | Chitosan microcapsules crosslinked using sodium tripolyphosphate by spray drying | IC50 value of acarbose was 4.81 μg/mL; R. tuberosa extract had IC50 value of 47.15 μg/mL; microcapsules of R. tuberosa extract prepared in pH 4, 0.1% (w/v) chitosan and 90 min stirring time had IC50 value of 50.65 μg/mL | [64] |
| Cagaita (Eugenia dysenterica) | Quercetin and its derivates | Spray-dried and freeze-dried powders produced with gum arabic or inulin | The IC50 values were in the range from 10.6 mg/mL to 107 mg/mL for powders prepared with gum arabic and from 9.8 mg/mL to 99.5 mg/mL for powders prepared with inulin | [65] |
| Cinnamon (Cinnamomum zeylanicum) extract | Proanthocyanidins | Spray-dried and freeze-dried microparticles formed using gelatin and five different polysaccharides—pectin, gum arabic, cashew gum, κ-carrageenan and carboxymethylcellulose | Inhibition of α-amylase, expressed as IC50 values ranged from 2.6 μg/mL (spray-dried samples) to 4.1 μg/mL (cinnamon extract) | [66] |
| King’s salad (Cosmos caudatus) extracts | n.d. | Spray-dried microcapsules | Microcapsules prepared in pH 4, 0.05% of chitosan and 90 min stirring time had optimum efficiency, with the IC50 value of 92.85 μg/mL, pure extract had IC50 value of 73.07 μg/mL, while for acarbose it was 4.83 μg/mL | [67] |
| Stevia (Stevia rebaudiana) leaves extract | n.d. | Maltodextrin microcapsules prepared by spray-drying process | For prepared samples, enzyme assay showed lack of inhibition at concentrations from 0.1 to 0.5 mg/mL | [68] |
| Winemaking byproducts | Malvidin-3-glucoside, quercetin-3-O-glucoside, catechin and epicatechin | Spray-dried grape skin powder and maltodextrin-encapsulated grape skin phenolics | IC50 values for grape skin and encapsulated grape skin phenolics were 0.44 mg/mL dry weight and 0.20 mg/mL dry weight | [69] |
| Blackberry (Rubus subg. Rubus) juice and apple fiber polyphenols | Cyanidin-3-glucoside, cyanidin 3-dioxalylglucoside, quercetin, ellagic acid, chlorogenic acid, phloretin and phlorizin | Freeze-dried apple fiber/blackberry juice complexes | Inhibition ranged from 22.10% (10% apple fiber addition) to 37.53% (4% apple fiber addition) | [70] |
| Blue pea (Clitoria ternatea) petal flower extract | n.d. | Beads prepared by microencapsulation process | Inhibition (%) of α-amylase was higher for microparticles (22.01%) than for phenolic extract (15.12%) | [71] |
| Raspberry (Rubus idaeus) juice polyphenols | n.d. | Cellulose-based complexes prepared by freeze-drying | Complexes formulated with lower amounts of cellulose had higher inhibition activities (%), for complexes prepared by 15 min of complexation the highest inhibition activity (56.07%) was observed for sample prepared with 2.5% of cellulose | [72] |
| Black beans (Glycine max) | n.d. | Freeze-dried powder prepared using combination of inulin, chitosan, whey protein isolate | Inhibition activity against α-amylase was around 68% | [73] |
| Olive pomace | Hydroxytyrosol, hydroxytyrosol glucoside, tyrosol and dihydrocaffeic acid | The lyophilisates were prepared using tyrosol in the gelatinized potato starch by mixing conventional and microwave heating during the freeze-drying | Tyrosol lyophilisate under microwave heating had the highest inhibition activity (IC50 was 559.57 µg/mL) | [74] |
| Bitter melon (Momordica charantia) juice polyphenols | n.d. | Fresh bitter juice was encapsulated using spray drying, as wall materials maltodextrin, gum arabic, citrus pectin and soy protein isolate were used | The lowest inhibitory activity was observed for encapsulates prepared with pectin and soy protein isolate | [75] |
| Blackcurrant (Ribes nigrum) concentrate | n.d. | Whey protein isolate was used as wall material in spray-drying and freeze-drying techniques | Freeze-dried samples (IC50 = 73.46 µg/mL) showed higher inhibitory activity than spray-dried samples (IC50 = 81.46 µg/mL) | [76] |
| Propolis extract | Vanillin, eugenol, ferulic acid, vanillic acid, caffeic acid and p-coumaric acid | Microencapsulation of propolis was conducted using gum arabic and chitosan as wall materials | Inhibition activity (IC50) was 0.55 mg/mL | [77] |
| Tart cherry (Prunus cerasus) polyphenols | Cyanidin-3-glucosyl-rutinoside, cyanidin-3- rutinoside, chlorogenic acid, coumaric acid, rutin, quercetin and (-)-epicatechin | Carboxymethylcellulose hydrogels | Inhibition ranged from 21.97% (for hydrogel prepared with 5% carboxymethylcellulose) to around 26% (for hydrogels prepared with 3% and 4% carboxymethylcellulose) | [78] |
| Individual Polyphenols | ||||
| Polyphenol | Delivery System | Inhibitory Activity | Ref. | |
| Curcumin | Chitosan–alginate polyelectrolyte complex | α-amylase activity of curcumin encapsulated in chitosan–tripolyphosphate was significantly higher after 30 min of incubation, when compared to curcumin nanoencapsulated in chitosan-alginate complex | [79] | |
| Rutin | Rutin was encapsulated with three types of carrier materials (starch, egg albumin, lipid) using three different techniques to investigate the impact of gastrointestinal digestion | Dialysable fractions showed lower anti-diabetic activity than undigested samples (except for the rutin encapsulated in egg albumin which showed the higher inhibition activity than undigested sample) | [80] | |
| Polyphenols from Plant Material | ||||
|---|---|---|---|---|
| Source | Polyphenols | Delivery System | Inhibitory Activity | Ref. |
| Mulberry fruit (Morus alba) extract | Cyanidin-3-glucoside, rutin, gallic acid | Freeze-dried particles prepared from maltodextrin | IC50 for mulberry fruit extract was 0.62 mg/mL and for encapsulated mulberry fruit extract it was 0.57 mg/mL | [62] |
| Cinnamon (Cinnamomum zeylanicum) extract | Proanthocyanidins | Spray-dried and freeze-dried microparticles formed using gelatin and five different polysaccharides—pectin, gum arabic, cashew gum, κ-carrageenan and carboxymethylcellulose | Inhibition of α-glucosidase (IC50 values) ranged from 0.7 μg/mL (freeze-dried samples) to 6.3 μg/mL (cinnamon extract) | [66] |
| Winemaking byproducts | Malvidin-3-glucoside, quercetin-3-O-glucoside, catechin and epicatechin | Spray-dried grape skin powder and maltodextrin-encapsulated grape skin phenolics | IC50 value of grape skin was 0.28 mg/mL dry weight and for encapsulated grape skin phenolics it was 0.15 mg/mL dry weight | [69] |
| Red pepper (Capsicum annuum) waste | Gallic acid, protocatechuic acid, epicatechin, chlorogenic acid, vanillic acid, caffeic acid, myricetin, quercetin and rutin | Freeze-dried and spray-dried encapsulates prepared to investigate inhibition before and during simulated gastric and intestinal fluids | Freeze-dried samples had higher potential to inhibit α-glucosidase than spray-dried samples, the simulated intestinal fluid showed the highest inhibition activity (%) for freeze-dried (31.6%) and spray-dried (30.42%) samples | [95] |
| Black beans (Glycine max) | n.d. | Freeze-dried powder prepared using combination of inulin, chitosan and whey-protein isolate | Inhibition activity against α-glucosidase was around 68% | [73] |
| Blackberry juice (Rubus subg. Rubus) and apple fiber polyphenols | Cyanidin-3-O-glucoside, cyanidin-3-O-dioxalylglucoside, ellagic acid, chlorogenic acid, rutin and phloridzin | Pectin/blackberry hydrogels enriched with apple fiber | IC50 value for acarbose was 105.61 µg GAE/mL; for prepared samples IC50 values ranged from 1.98 µg GAE/mL (hydrogel with low-methoxylated pectin) to 6.55 µg GAE/mL (hydrogel with high-methoxylated pectin enriched with apple fiber) | [96] |
| Tart cherry (Prunus cerasus) polyphenols | Cyanidin-3-glucosyl-rutinoside, cyanidin-3- rutinoside, chlorogenic acid, coumaric acid, rutin, quercetin and (-)-epicatechin | Carboxymethylcellulose hydrogels | Inhibition ranged from 38.39% (for hydrogel prepared with 5% carboxymethylcellulose) to 56.64% (for hydrogel prepared with 3% carboxymethylcellulose) | [78] |
| Individual Polyphenols | ||||
| Polyphenol | Delivery System | Inhibitory Activity | Ref. | |
| Catechin hydrate | Starch-based nanoparticles from three sources: horse chestnut, water chestnut and lotus stem | Before the process of simulated gastrointestinal conditions, catechin had inhibitory activity of 92.4%, while free catechin after the process of simulated gastrointestinal conditions had inhibitory activity of 88.79% | [97] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ćorković, I.; Gašo-Sokač, D.; Pichler, A.; Šimunović, J.; Kopjar, M. Dietary Polyphenols as Natural Inhibitors of α-Amylase and α-Glucosidase. Life 2022, 12, 1692. https://doi.org/10.3390/life12111692
Ćorković I, Gašo-Sokač D, Pichler A, Šimunović J, Kopjar M. Dietary Polyphenols as Natural Inhibitors of α-Amylase and α-Glucosidase. Life. 2022; 12(11):1692. https://doi.org/10.3390/life12111692
Chicago/Turabian StyleĆorković, Ina, Dajana Gašo-Sokač, Anita Pichler, Josip Šimunović, and Mirela Kopjar. 2022. "Dietary Polyphenols as Natural Inhibitors of α-Amylase and α-Glucosidase" Life 12, no. 11: 1692. https://doi.org/10.3390/life12111692
APA StyleĆorković, I., Gašo-Sokač, D., Pichler, A., Šimunović, J., & Kopjar, M. (2022). Dietary Polyphenols as Natural Inhibitors of α-Amylase and α-Glucosidase. Life, 12(11), 1692. https://doi.org/10.3390/life12111692