
Essential Roles of Exocyst Complex Component 3-like 2 on Cardiovascular Development in Mice
 , , ,
, , ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Reverse Transcription (RT)-Quantitative (q) PCR
2.3. Single Cell (sc) RNA-Seq Analysis
2.4. Immunohistochemistry
2.5. Immunostaining of Whole-Mount Retinas
2.6. Image Acquisition
2.7. Flowcytometry Analysis
2.8. CT imaging Analyses
2.9. Statistical Analysis
3. Results
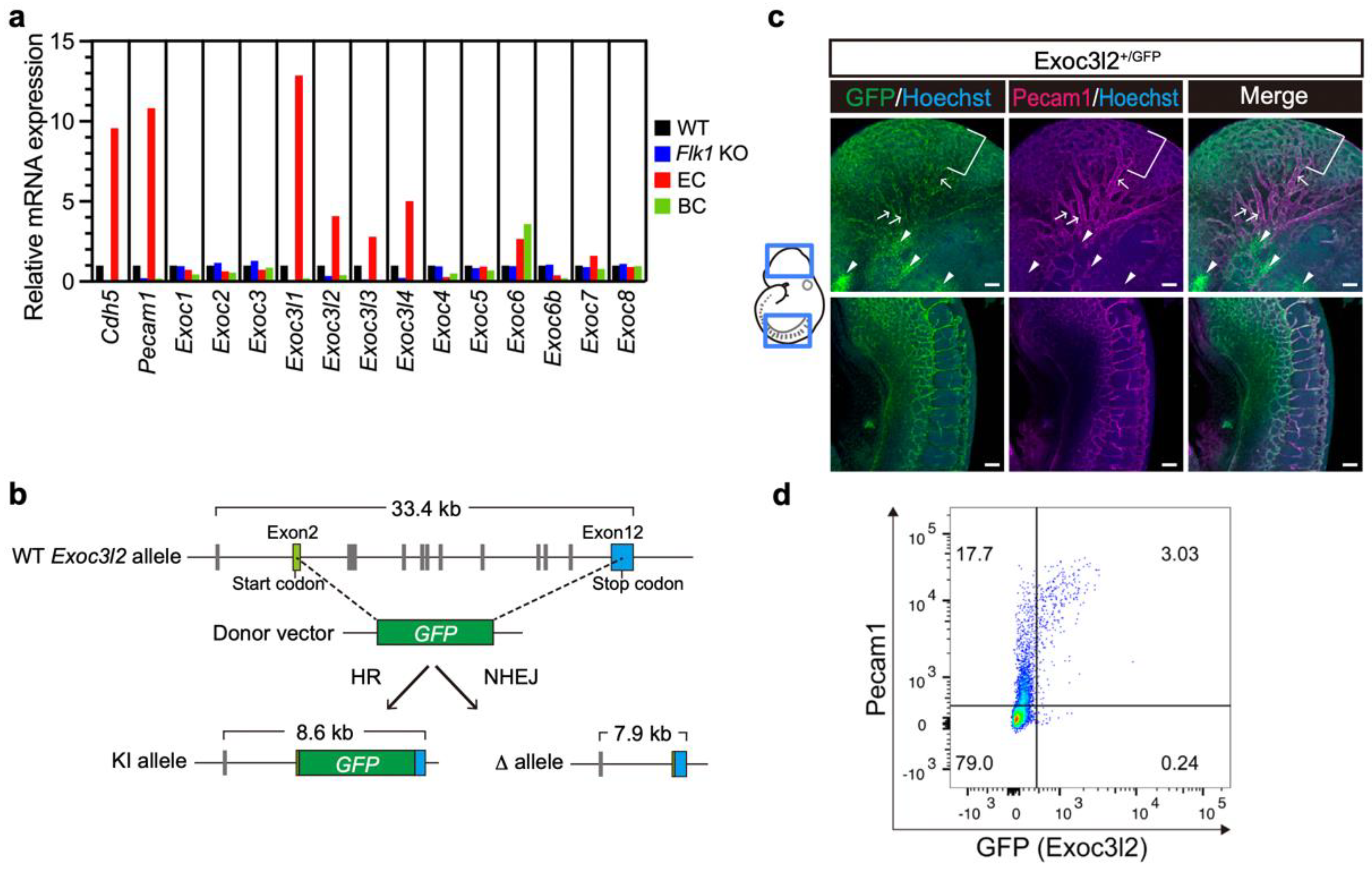
3.1. Expression of Exoc3l Members and Generation of Exoc3l2 KO Mice
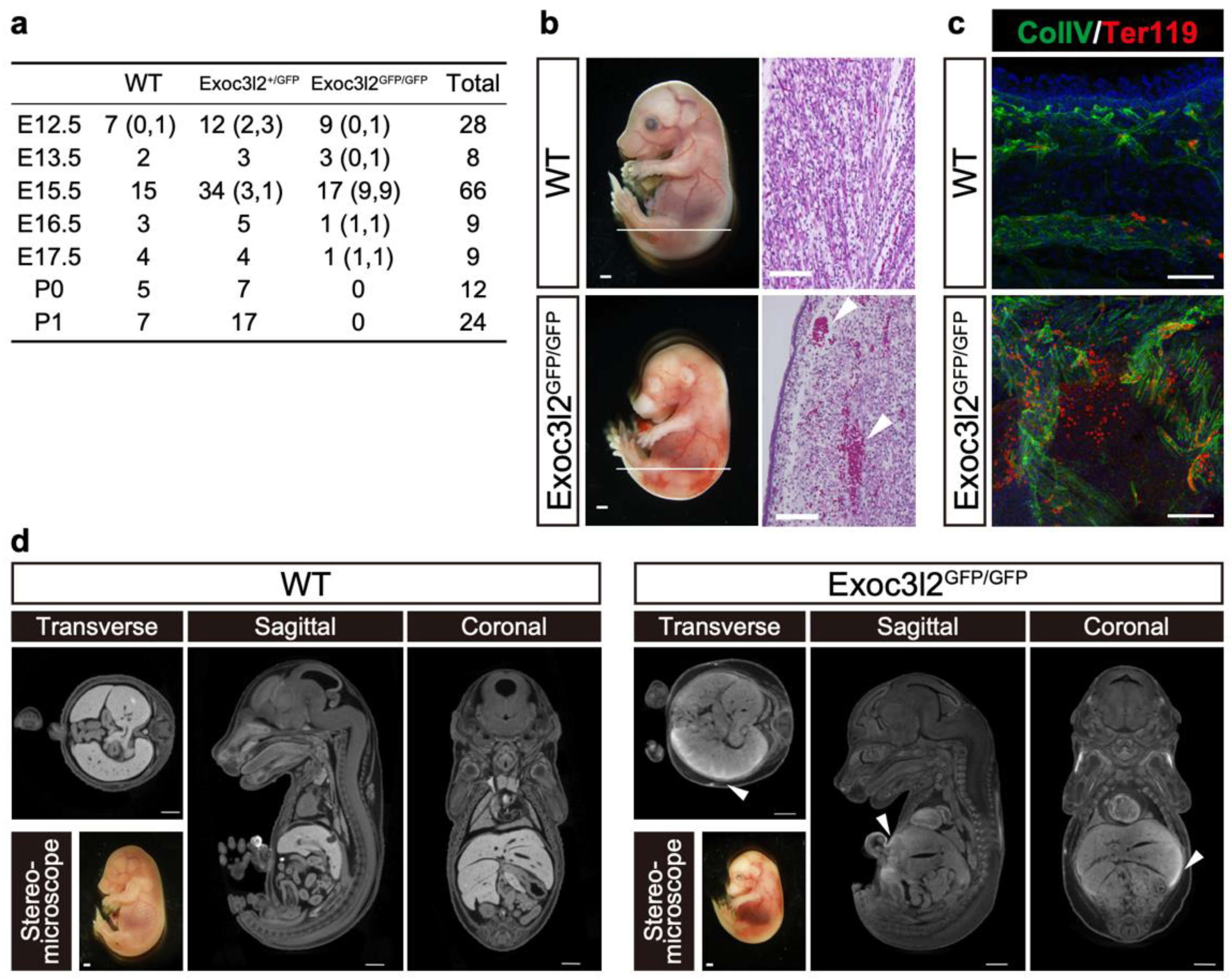
3.2. Phenotypes of Exoc3l2 KO Embryo
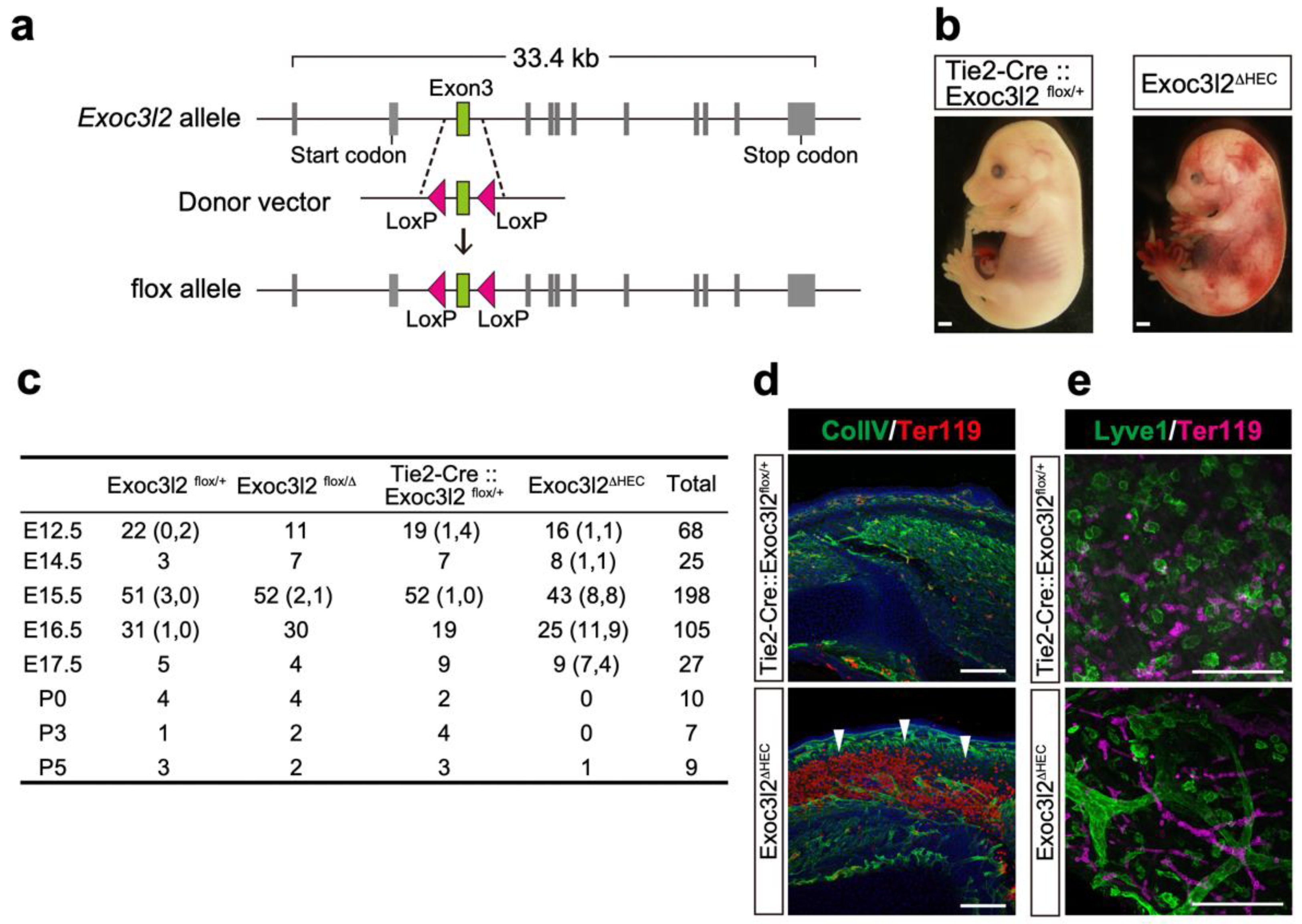
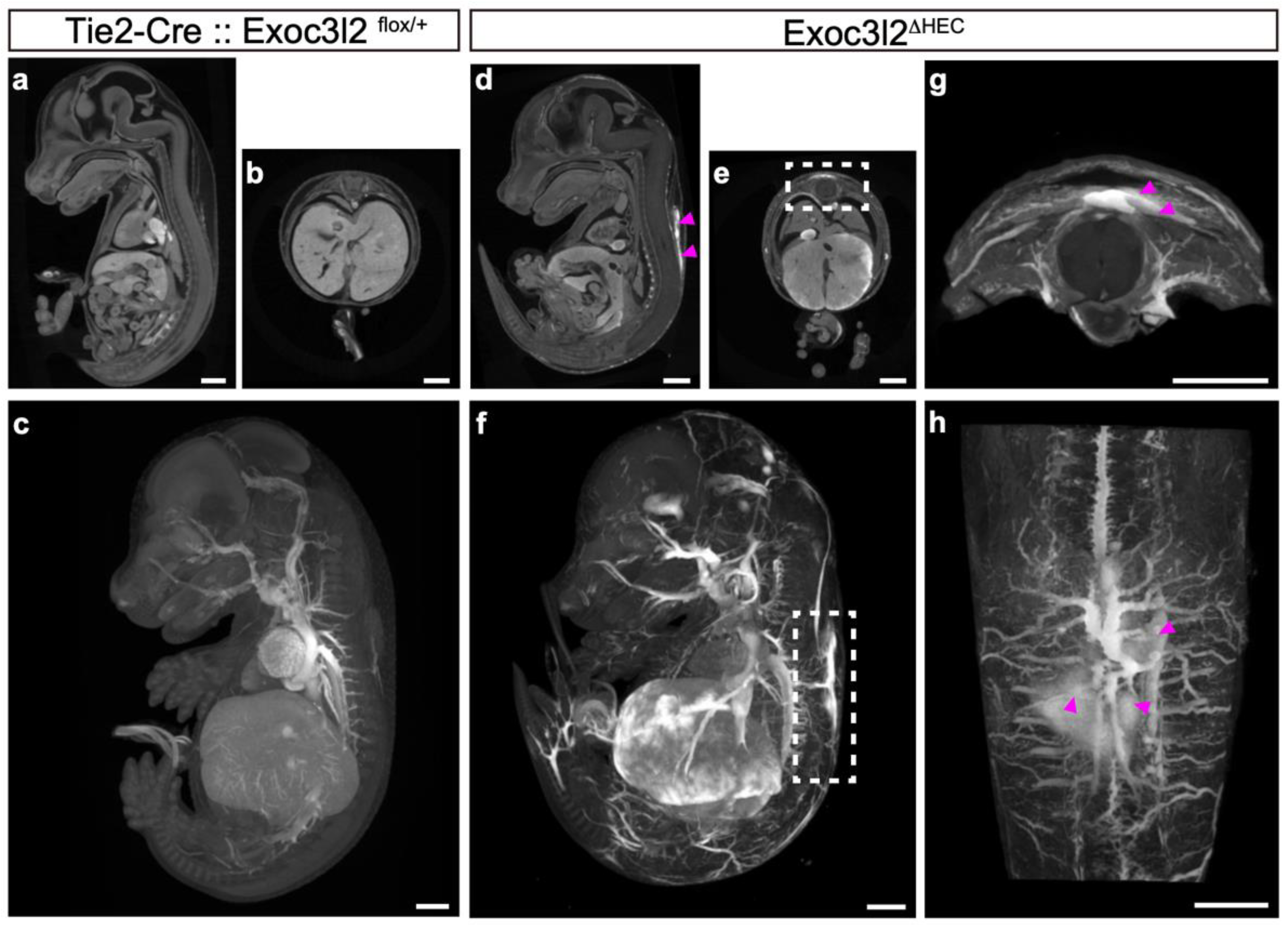
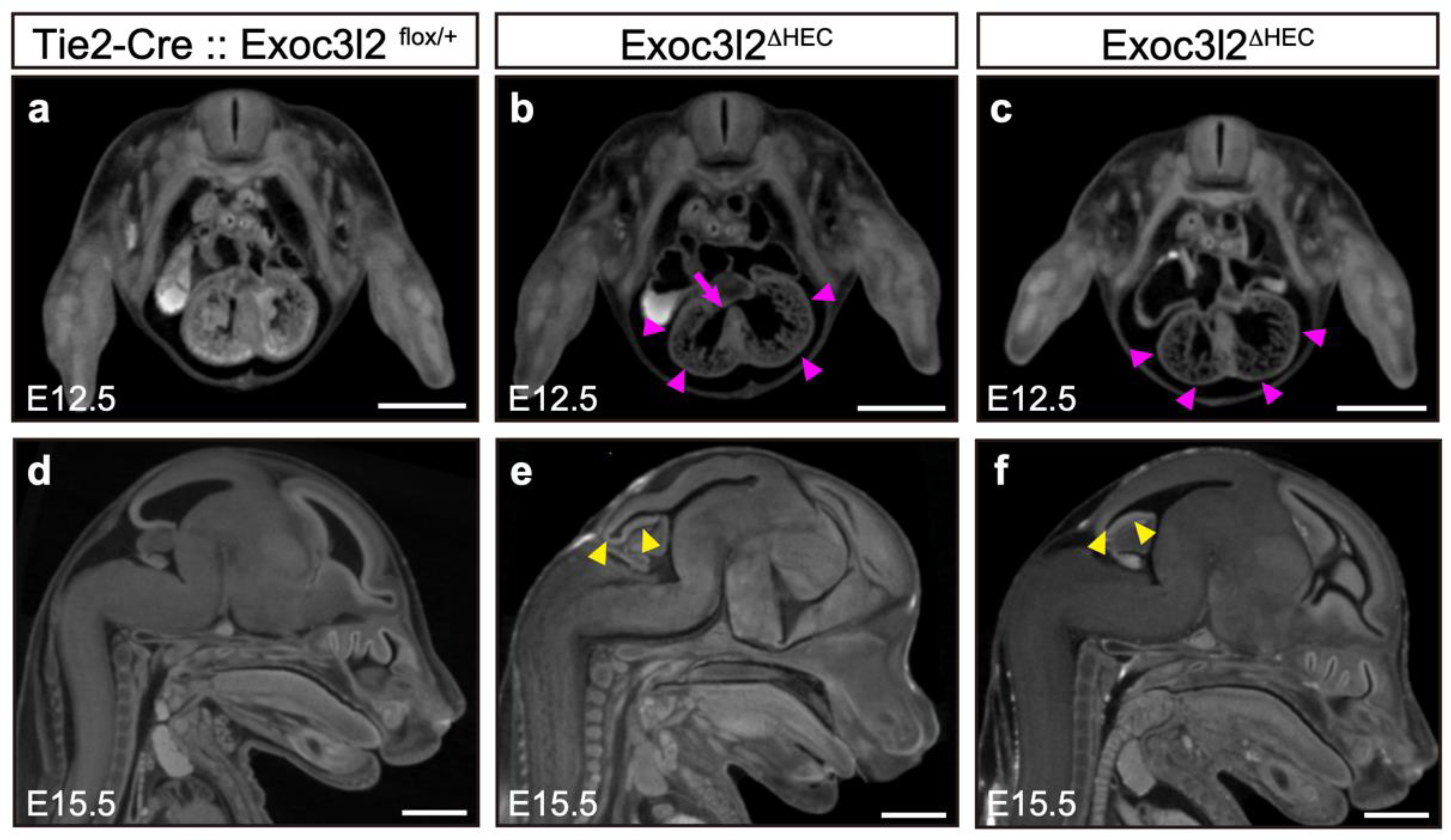
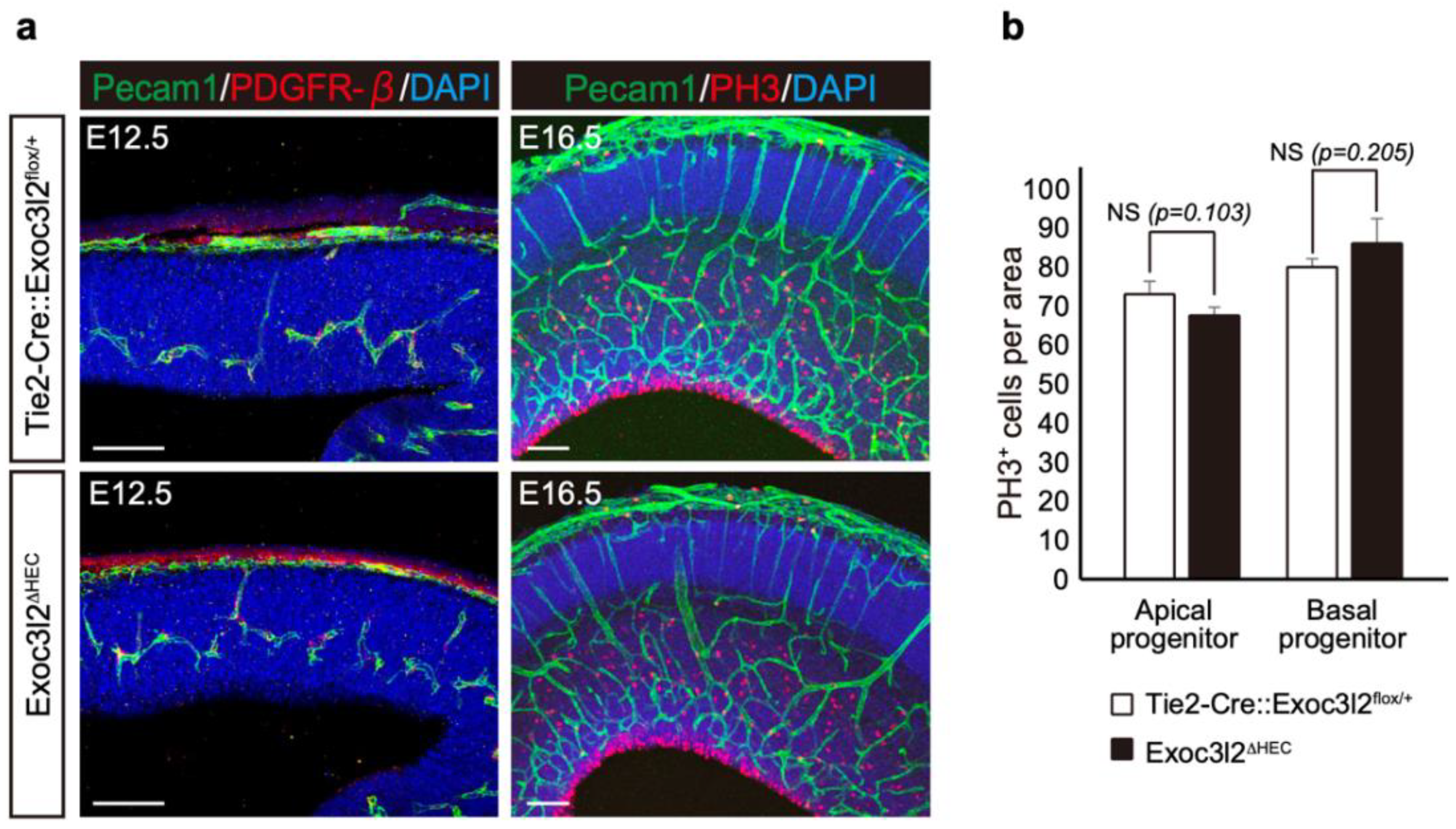
3.3. Phenotypes of Tie2-Cre: Exoc3l2 Conditional Knock-Out Embryos
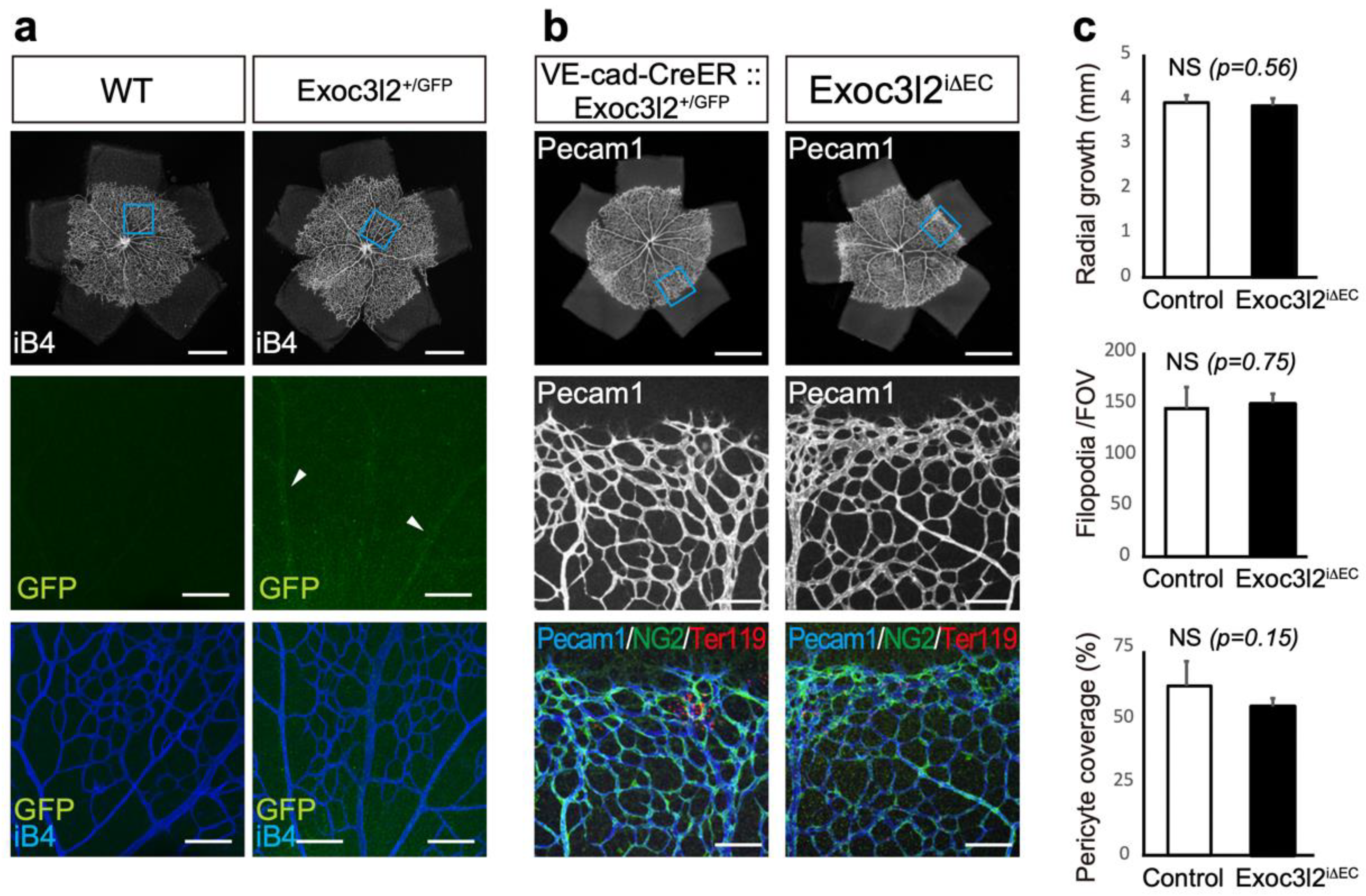
3.4. Postnatal Roles of Exoc3l2 in Retinal Vascular Development
3.5. Roles of Exoc3l2 in Heart and Brain Development
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Adams, R.H.; Alitalo, K. Molecular Regulation of Angiogenesis and Lymphangiogenesis. Nat. Rev. Mol. Cell Biol. 2007, 8, 464–478. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Jain, R.K. Molecular Mechanisms and Clinical Applications of Angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.; Guo, W. The Exocyst at a Glance. J. Cell Sci. 2015, 128, 2957–2964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.; Feng, S.; Wu, B.; Guo, W. Polarized Exocytosis. Cold Spring Harb. Perspect. Biol. 2017, 9, a027870. [Google Scholar] [CrossRef] [Green Version]
- Polgar, N.; Fogelgren, B. Regulation of Cell Polarity by Exocyst-Mediated Trafficking. Cold Spring Harb. Perspect. Biol. 2018, 10, a031401. [Google Scholar] [CrossRef]
- Novick, P.; Schekman, R. Secretion and Cell-Surface Growth Are Blocked in a Temperature-Sensitive Mutant of Saccharomyces Cerevisiae. Proc. Natl. Acad. Sci. USA 1979, 76, 1858–1862. [Google Scholar] [CrossRef] [Green Version]
- Shaheen, R.; Szymanska, K.; Basu, B.; Patel, N.; Ewida, N.; Faqeih, E.; Al Hashem, A.; Derar, N.; Alsharif, H.; Aldahmesh, M.A.; et al. Characterizing the Morbid Genome of Ciliopathies. Genome Biol. 2016, 17, 242. [Google Scholar] [CrossRef] [Green Version]
- Shalata, A.; Lauhasurayotin, S.; Leibovitz, Z.; Li, H.; Hebert, D.; Dhanraj, S.; Hadid, Y.; Mahroum, M.; Bajar, J.; Egenburg, S.; et al. Biallelic Mutations in EXOC3L2 Cause a Novel Syndrome That Affects the Brain, Kidney and Blood. J. Med. Genet. 2019, 56, 340–346. [Google Scholar] [CrossRef]
- Smogavec, M.; Gerykova Bujalkova, M.; Lehner, R.; Neesen, J.; Behunova, J.; Yerlikaya-Schatten, G.; Reischer, T.; Altmann, R.; Weis, D.; Duba, H.-C.; et al. Singleton Exome Sequencing of 90 Fetuses with Ultrasound Anomalies Revealing Novel Disease-Causing Variants and Genotype-Phenotype Correlations. Eur. J. Hum. Genet. 2022, 30, 428–438. [Google Scholar] [CrossRef]
- Takase, H.; Matsumoto, K.; Yamadera, R.; Kubota, Y.; Otsu, A.; Suzuki, R.; Ishitobi, H.; Mochizuki, H.; Kojima, T.; Takano, S.; et al. Genome-Wide Identification of Endothelial Cell-Enriched Genes in the Mouse Embryo. Blood 2012, 120, 914–923. [Google Scholar] [CrossRef]
- Mizuno-Iijima, S.; Ayabe, S.; Kato, K.; Matoba, S.; Ikeda, Y.; Dinh, T.T.H.; Le, H.T.; Suzuki, H.; Nakashima, K.; Hasegawa, Y.; et al. Efficient Production of Large Deletion and Gene Fragment Knock-in Mice Mediated by Genome Editing with Cas9-Mouse Cdt1 in Mouse Zygotes. Methods 2021, 191, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Cao, J.; Martin, B.K.; Li, T.; Welsh, I.C.; Srivatsan, S.; Huang, X.; Calderon, D.; Noble, W.S.; Disteche, C.M.; et al. Systematic Reconstruction of Cellular Trajectories across Mouse Embryogenesis. Nat. Genet. 2022, 54, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, C.; Matsushita, J.; Azami, T.; Tsukiyama-Fujii, S.; Tsukiyama, T.; Mizuno, S.; Takahashi, S.; Ema, M. Generating Vegfr3 Reporter Transgenic Mouse Expressing Membrane-Tagged Venus for Visualization of VEGFR3 Expression in Vascular and Lymphatic Endothelial Cells. PLoS ONE 2019, 14, e0210060. [Google Scholar] [CrossRef] [PubMed]
- Komabayashi-Suzuki, M.; Yamanishi, E.; Watanabe, C.; Okamura, M.; Tabata, H.; Iwai, R.; Ajioka, I.; Matsushita, J.; Kidoya, H.; Takakura, N.; et al. Spatiotemporally Dependent Vascularization Is Differently Utilized among Neural Progenitor Subtypes during Neocortical Development. Cell Rep. 2019, 29, 1113–1129.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishitobi, H.; Wakamatsu, A.; Liu, F.; Azami, T.; Hamada, M.; Matsumoto, K.; Kataoka, H.; Kobayashi, M.; Choi, K.; Nishikawa, S.; et al. Molecular Basis for Flk1 Expression in Hemato-Cardiovascular Progenitors in the Mouse. Development 2011, 138, 5357–5368. [Google Scholar] [CrossRef] [Green Version]
- Okabe, K.; Kobayashi, S.; Yamada, T.; Kurihara, T.; Tai-Nagara, I.; Miyamoto, T.; Mukouyama, Y.S.; Sato, T.N.; Suda, T.; Ema, M.; et al. Neurons Limit Angiogenesis by Titrating VEGF in Retina. Cell 2014, 159, 584–596. [Google Scholar] [CrossRef] [Green Version]
- Ichiyama, Y.; Obata, S.; Saishin, Y.; Sawada, O.; Kakinoki, M.; Sawada, T.; Kubota, Y.; Ohji, M. The Systemic Antiangiogenic Effect of Intravitreal Aflibercept Injection in a Mouse Model of Retinopathy of Prematurity. FASEB J. 2021, 35, e21390. [Google Scholar] [CrossRef]
- Dickinson, M.E.; Flenniken, A.M.; Ji, X.; Teboul, L.; Wong, M.D.; White, J.K.; Meehan, T.F.; Weninger, W.J.; Westerberg, H.; Adissu, H.; et al. High-Throughput Discovery of Novel Developmental Phenotypes. Nature 2016, 537, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Tamura, M.; Hosoya, M.; Fujita, M.; Iida, T.; Amano, T.; Maeno, A.; Kataoka, T.; Otsuka, T.; Tanaka, S.; Tomizawa, S.; et al. Overdosage of Hand2 Causes Limb and Heart Defects in the Human Chromosomal Disorder Partial Trisomy Distal 4q. Hum. Mol. Genet. 2013, 22, 2471–2481. [Google Scholar] [CrossRef] [Green Version]
- Escobedo, N.; Oliver, G. Lymphangiogenesis: Origin, Specification, and Cell Fate Determination. Annu. Rev. Cell Dev. Biol. 2016, 32, 677–691. [Google Scholar] [CrossRef]
- Oliver, G.; Kipnis, J.; Randolph, G.J.; Harvey, N.L. The Lymphatic Vasculature in the 21(St) Century: Novel Functional Roles in Homeostasis and Disease. Cell 2020, 182, 270–296. [Google Scholar] [CrossRef] [PubMed]
- Grimsley-Myers, C.M.; Isaacson, R.H.; Cadwell, C.M.; Campos, J.; Hernandes, M.S.; Myers, K.R.; Seo, T.; Giang, W.; Griendling, K.K.; Kowalczyk, A.P. VE-Cadherin Endocytosis Controls Vascular Integrity and Patterning during Development. J. Cell Biol. 2020, 219, e201909081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langevin, J.; Morgan, M.J.; Sibarita, J.B.; Aresta, S.; Murthy, M.; Schwarz, T.; Camonis, J.; Bellaïche, Y. Drosophila Exocyst Components Sec5, Sec6, and Sec15 Regulate DE-Cadherin Trafficking from Recycling Endosomes to the Plasma Membrane. Dev. Cell 2005, 9, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Xiong, X.; Xu, Q.; Huang, Y.; Singh, R.D.; Anderson, R.; Leof, E.; Hu, J.; Ling, K. An Association between Type Iγ PI4P 5-Kinase and Exo70 Directs E-Cadherin Clustering and Epithelial Polarization. Mol. Biol. Cell 2012, 23, 87–98. [Google Scholar] [CrossRef]
- Jafar-Nejad, H.; Andrews, H.K.; Acar, M.; Bayat, V.; Wirtz-Peitz, F.; Mehta, S.Q.; Knoblich, J.A.; Bellen, H.J. Sec15, a Component of the Exocyst, Promotes Notch Signaling during the Asymmetric Division of Drosophila Sensory Organ Precursors. Dev. Cell 2005, 9, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Harmelink, C.; Baldwin, H.S. Tie2 Regulates Endocardial Sprouting and Myocardial Trabeculation. JCI Insight 2019, 5, e96002. [Google Scholar] [CrossRef] [Green Version]
- Barkefors, I.; Fuchs, P.F.; Heldin, J.; Bergström, T.; Forsberg-Nilsson, K.; Kreuger, J. Exocyst Complex Component 3-like 2 (EXOC3L2) Associates with the Exocyst Complex and Mediates Directional Migration of Endothelial Cells. J. Biol. Chem. 2011, 286, 24189–24199. [Google Scholar] [CrossRef] [Green Version]
- Seshadri, S.; Fitzpatrick, A.L.; Ikram, M.A.; DeStefano, A.L.; Gudnason, V.; Boada, M.; Bis, J.C.; Smith, A.V.; Carassquillo, M.M.; Lambert, J.C.; et al. Genome-Wide Analysis of Genetic Loci Associated with Alzheimer Disease. JAMA 2010, 303, 1832–1840. [Google Scholar] [CrossRef] [Green Version]
- Shang, H.; Fu, J.; Zhang, X.-M.; Song, R.-R.; Wang, W.-Z. Association between EXOC3L2 Rs597668 Polymorphism and Alzheimer’s Disease. CNS Neurosci. Ther. 2013, 19, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.-J.; Sun, S.-Y.; Yan, C.-J.; Cheng, Z.-C.; Yang, M.-F.; Li, Z.-F.; Cheng, H.-W.; Fang, T.-K. EXOC3L2 Rs597668 Variant Contributes to Alzheimer’s Disease Susceptibility in Asian Population. Oncotarget 2017, 8, 20086–20091. [Google Scholar] [CrossRef]
- Wijsman, E.M.; Pankratz, N.D.; Choi, Y.; Rothstein, J.H.; Faber, K.M.; Cheng, R.; Lee, J.H.; Bird, T.D.; Bennett, D.A.; Diaz-Arrastia, R.; et al. Genome-Wide Association of Familial Late-Onset Alzheimer’s Disease Replicates BIN1 and CLU and Nominates CUGBP2 in Interaction with APOE. PLoS Genet. 2011, 7, e1001308. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, C.; Shibuya, H.; Ichiyama, Y.; Okamura, E.; Tsukiyama-Fujii, S.; Tsukiyama, T.; Matsumoto, S.; Matsushita, J.; Azami, T.; Kubota, Y.; et al. Essential Roles of Exocyst Complex Component 3-like 2 on Cardiovascular Development in Mice. Life 2022, 12, 1730. https://doi.org/10.3390/life12111730
Watanabe C, Shibuya H, Ichiyama Y, Okamura E, Tsukiyama-Fujii S, Tsukiyama T, Matsumoto S, Matsushita J, Azami T, Kubota Y, et al. Essential Roles of Exocyst Complex Component 3-like 2 on Cardiovascular Development in Mice. Life. 2022; 12(11):1730. https://doi.org/10.3390/life12111730
Chicago/Turabian StyleWatanabe, Chisato, Hirotoshi Shibuya, Yusuke Ichiyama, Eiichi Okamura, Setsuko Tsukiyama-Fujii, Tomoyuki Tsukiyama, Shoma Matsumoto, Jun Matsushita, Takuya Azami, Yoshiaki Kubota, and et al. 2022. "Essential Roles of Exocyst Complex Component 3-like 2 on Cardiovascular Development in Mice" Life 12, no. 11: 1730. https://doi.org/10.3390/life12111730
APA StyleWatanabe, C., Shibuya, H., Ichiyama, Y., Okamura, E., Tsukiyama-Fujii, S., Tsukiyama, T., Matsumoto, S., Matsushita, J., Azami, T., Kubota, Y., Ohji, M., Sugiyama, F., Takahashi, S., Mizuno, S., Tamura, M., Mizutani, K. -i., & Ema, M. (2022). Essential Roles of Exocyst Complex Component 3-like 2 on Cardiovascular Development in Mice. Life, 12(11), 1730. https://doi.org/10.3390/life12111730