
Chemical Profiling and Molecular Docking Study of Agathophora alopecuroides

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Metabolites Profiling of the Methanolic Extract of A. alopecuroides
2.3. Molecular Docking Study
3. Results
3.1. In Vitro Testing of the Antidibetic Activity of A. alopecuroides (AAE)
3.2. Metabolomic Profiling of the Methanolic Extract of A. alopecuroides (AAE)
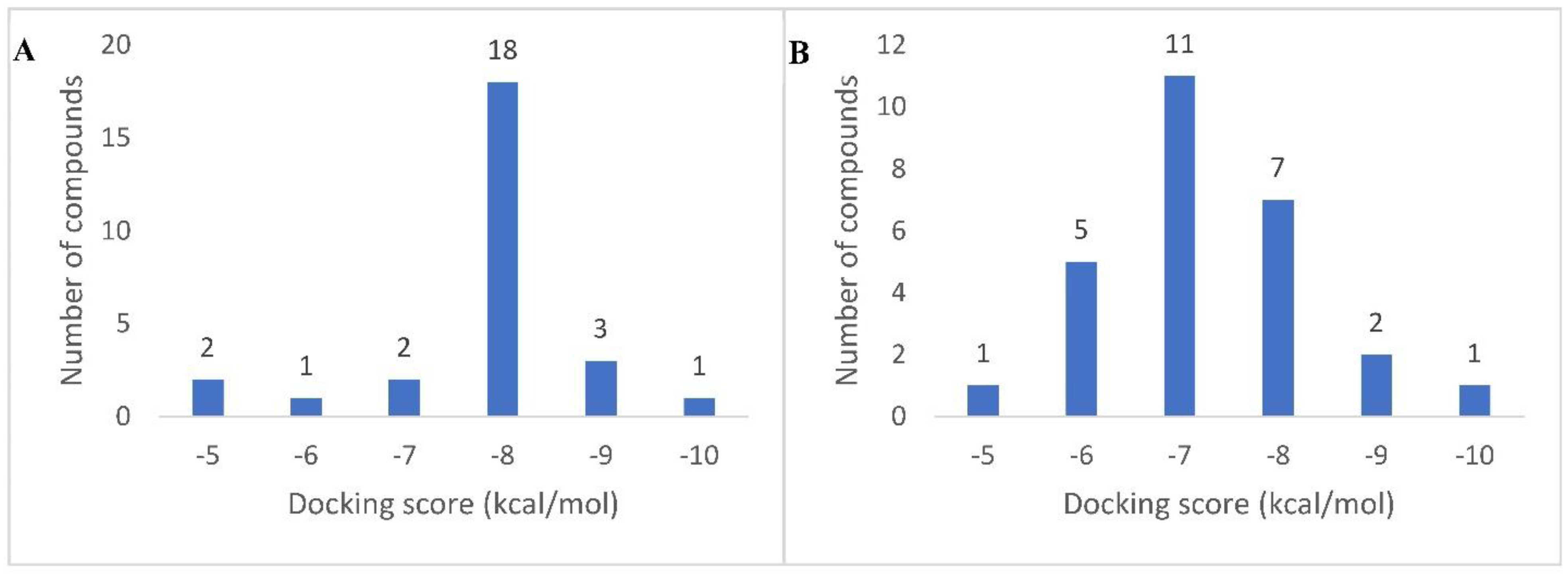
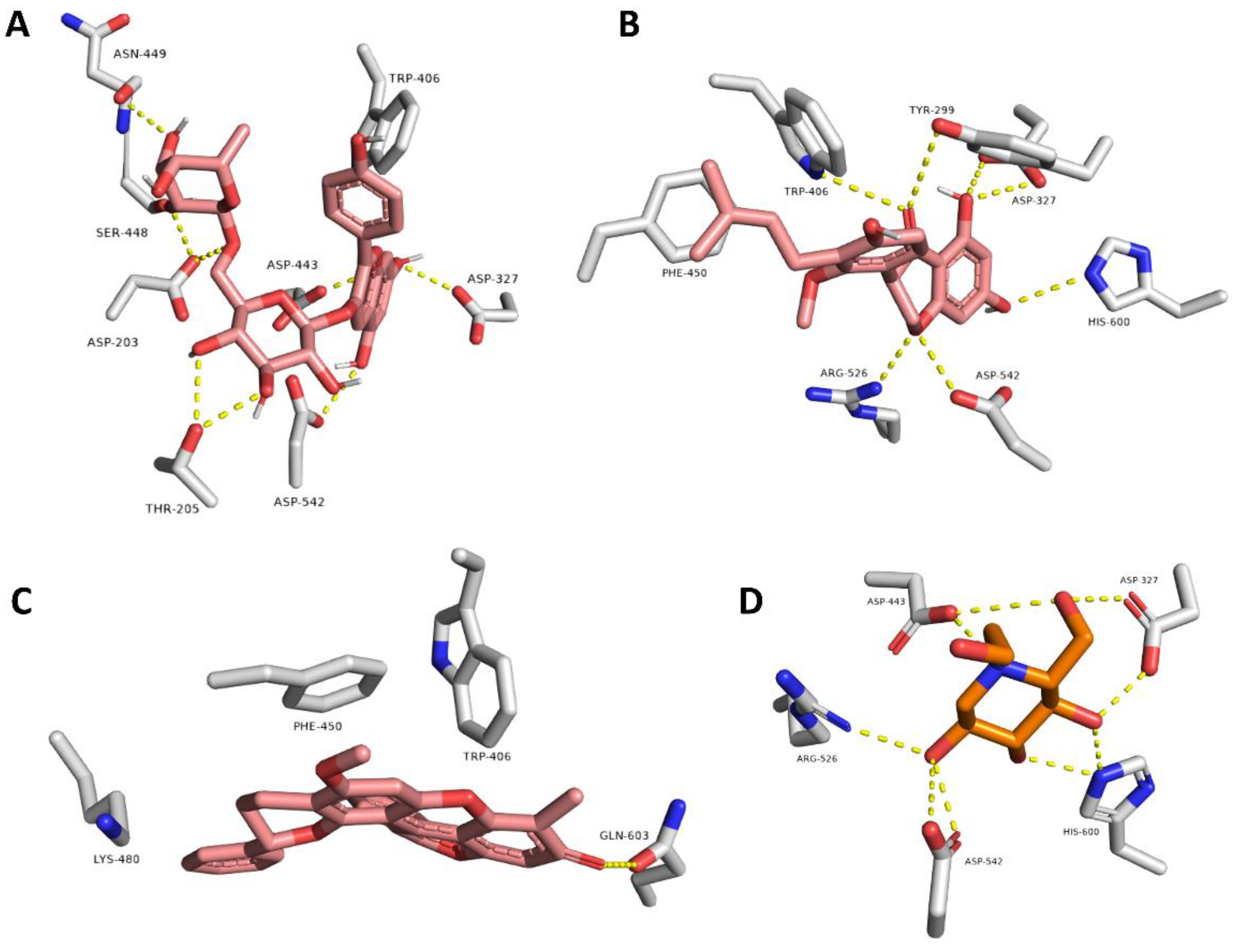
3.3. Molecular Docking Analysis
4. Discussion
- Metabolomic profiling of the methanolic extract of A. alopecuroides
- Molecular docking study of A. alopecuroides metabolites for inhibition of α-amylase and α-glucosidase enzymes
- Bridging the metabolomic profiling of A. alopecuroides with its biological activity.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Belayneh, Y.M.; Birru, E.M. Antidiabetic activities of hydromethanolic leaf extract of Calpurnia aurea (Ait.) Benth. Subspecies aurea (Fabaceae) in mice. Evid. Based Complement. Altern. Med. 2018, 2018, 3509073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alqahtani, A.S.; Hidayathulla, S.; Rehman, T.; ElGamal, A.A.; Al-Massarani, S.; Razmovski-Naumovski, V.; Alqahtani, M.S.; El Dib, R.A.; AlAjmi, M.F. Alpha-Amylase and Alpha-Glucosidase Enzyme Inhibition and Antioxidant Potential of 3-Oxolupenal and Katononic Acid Isolated from Nuxia oppositifolia. Biomolecules 2019, 10, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajaria, D.; Tiwari, S.; Tripathi, J.; Tripathi, Y.; Ranjana. In-vitro α amylase and glycosidase inhibitory effect of ethanolic extract of antiasthmatic drug—Shirishadi. J. Am. Pharm. Technol. Res. 2013, 4, 206. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Zhu, L.; Chen, Y.; Tian, J.; Wang, Y. In vivo and in vitro antioxidant activity and α-glucosidase, α-amylase inhibitory effects of flavonoids from Cichorium glandulosum seeds. Food Chem. 2013, 139, 59–66. [Google Scholar] [CrossRef]
- Nair, S.S.; Kavrekar, V.; Mishra, A. In vitro studies on alpha amylase and alpha glucosidase inhibitory activities of selected plant extracts. Eur. J. Exp. Biol. 2013, 3, 128–132. [Google Scholar]
- Mumtaz, M.W.; Al-Zuaidy, M.H.; Hamid, A.A.; Danish, M.; Akhtar, M.T.; Mukhtar, H. Metabolite profiling and inhibitory properties of leaf extracts of Ficus benjamina towards α-glucosidase and α-amylase. Int. J. Food Prop. 2018, 21, 1560–1574. [Google Scholar] [CrossRef] [Green Version]
- Amin, E.; Abdel-Bakky, M.S.; Darwish, M.A.; Mohammed, H.A.; Chigurupati, S.; Qureshi, K.A.; Hassan, M.H. The Glycemic Control Potential of Some Amaranthaceae Plants, with Particular Reference to In Vivo Antidiabetic Potential of Agathophora alopecuroides. Molecules 2022, 27, 973. [Google Scholar] [CrossRef]
- Amin, E.; Abdel-Bakky, M.S.; Mohammed, H.A.; Chigrupati, S.; Qureshi, K.A.; Hassan, M.H.A. Phytochemical Analysis and Evaluation of the Antioxidant and Antimicrobial Activities of Five Halophytes from Qassim Flora. Pol. J. Environ. Stud. 2022, 31, 3005–3012. [Google Scholar] [CrossRef]
- Chikhi, I.; Allali, H.; Dib, M.E.A.; Medjdoub, H.; Tabti, B. Antidiabetic activity of aqueous leaf extract of Atriplex halimus L. (Chenopodiaceae) in streptozotocin–induced diabetic rats. Asian Pac. J. Trop. Dis. 2014, 4, 181–184. [Google Scholar] [CrossRef]
- Othman, A.; Sayed, A.M.; Amen, Y.; Shimizu, K. Possible neuroprotective effects of amide alkaloids from Bassia indica and Agathophora alopecuroides: In vitro and in silico investigations. RSC Adv. 2022, 12, 18746–18758. [Google Scholar] [CrossRef]
- Sashidhara, K.V.; Rosaiah, J.N. Various Dereplication Strategies Using LC-MS for Rapid Natural Product Lead Identification and Drug Discovery. Nat. Prod. Commun. 2007, 2, 193–202. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular Docking and Structure-Based Drug Design Strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput. Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, L.; Jayakanthan, K.; Mohan, S.; Nasi, R.; Johnston, B.D.; Pinto, B.M.; Rose, D.R. New Glucosidase Inhibitors from an Ayurvedic Herbal Treatment for Type 2 Diabetes: Structures and Inhibition of Human Intestinal Maltase-Glucoamylase with Compounds from Salacia reticulata. Biochemistry 2009, 49, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.K.; Zhang, X.; Caner, S.; Tysoe, C.; Nguyen, N.T.; Wicki, J.; Williams, D.E.; Coleman, J.; McNeill, J.H.; Yuen, V.; et al. The amylase inhibitor montbretin A reveals a new glycosidase inhibition motif. Nat. Chem. Biol. 2015, 11, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Bastida, J.; Viladomat, F.; Bergoñon, S.; Fernandez, J.M.; Codina, C.; Rubiralta, M.; Quirion, J.-C. Alkaloids from Narcissus leonensis. Phytochemistry 1993, 34, 1656–1658. [Google Scholar] [CrossRef]
- Kam, T.-S.; Tan, P.-S.; Chuah, C.-H. Alkaloids from leaves of Kopsia larutensis. Phytochemistry 1992, 31, 2936–2938. [Google Scholar] [CrossRef]
- Cahlíková, L.; Vaněčková, N.; Šafratová, M.; Breiterová, K.; Blunden, G.; Hulcová, D.; Opletal, L. The Genus Nerine Herb. (Amaryllidaceae): Ethnobotany, Phytochemistry, and Biological Activity. Molecules 2019, 24, 4238. [Google Scholar] [CrossRef] [Green Version]
- Cahlíková, L.; Zavadil, S.; Macáková, K.; Valterova, I.; Kulhánková, A.; Hostalkova, A.; Kuneš, J.; Opletal, L. Isolation and Cholinesterase Activity of Amaryllidaceae Alkaloids from Nerine bowdenii. Nat. Prod. Commun. 2011, 6, 1827–1830. [Google Scholar] [CrossRef] [Green Version]
- Piboonprai, K.; Khumkhrong, P.; Khongkow, M.; Yata, T.; Ruangrungsi, N.; Chansriniyom, C.; Iempridee, T. Anticancer activity of arborinine from Glycosmis parva leaf extract in human cervical cancer cells. Biochem. Biophys. Res. Commun. 2018, 500, 866–872. [Google Scholar] [CrossRef]
- Hagel, J.M.; Facchini, P.J. Benzylisoquinoline Alkaloid Metabolism: A Century of Discovery and a Brave New World. Plant Cell Physiol. 2013, 54, 647–672. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, Y.; Moriyasu, M.; Ichimaru, M.; Iwasa, K.; Kato, A.; Mathenge, S.G.; Mutiso, P.B.C.; Juma, F.D. Quaternary isoquinoline alkaloids from Xylopia parviflora. Phytochemistry 2004, 65, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Steyn, P.S.; Vleggaar, R.; Wessels, P.L.; Cole, R.J.; Scott, D.B. Structure and carbon-13 nuclear magnetic resonance assignments of versiconal acetate, versiconol acetate, and versiconol, metabolites from cultures of Aspergillus parasiticus treated with dichlorvos. J. Chem. Soc. Perkin Trans. 1 1979, 451–459. [Google Scholar] [CrossRef]
- Mitscher, L.A.; Gollapudi, S.R.; Oburn, D.S.; Drake, S. Antimicrobial agents from higher plants: Two dimethylbenzisochromans from Karwinskia humboldtiana. Phytochemistry 1985, 24, 1681–1683. [Google Scholar] [CrossRef]
- Li, Y.; Xie, S.; Ying, J.; Wei, W.; Gao, K. Chemical Structures of Lignans and Neolignans Isolated from Lauraceae. Molecules 2018, 23, 3164. [Google Scholar] [CrossRef] [Green Version]
- Macías-Villamizar, V.; Cuca-Suárez, L.; González, F.V.; Rodríguez, S. Lignoids Isolated from Nectandra turbacensis (Kunth) Nees (Lauraceae). Rec. Nat. Prod. 2016, 10, 654. [Google Scholar]
- Qurtam, A.A.; Mechchate, H.; Es-Safi, I.; Al-Zharani, M.; Nasr, F.A.; Noman, O.M.; Aleissa, M.; Imtara, H.; Aleissa, A.M.; Bouhrim, M.; et al. Citrus Flavanone Narirutin, In Vitro and In Silico Mechanistic Antidiabetic Potential. Pharmaceutics 2021, 13, 1818. [Google Scholar] [CrossRef]
- Xu, Y.; Xie, L.; Xie, J.; Liu, Y.; Chen, W. Pelargonidin-3-O-rutinoside as a novel α-glucosidase inhibitor for improving postprandial hyperglycemia. Chem. Commun. 2018, 55, 39–42. [Google Scholar] [CrossRef]
- Viljoen, A.; Mncwangi, N.; Vermaak, I. Anti-Inflammatory Iridoids of Botanical Origin. Curr. Med. Chem. 2012, 19, 2104–2127. [Google Scholar] [CrossRef] [Green Version]
- Ersöz, T.; Berkman, M.; Taşdemir, D.; Çaliş, İ.; Ireland, C.M. Iridoid and phenylethanoid glycosides from Euphrasia pectinata. Turk. J. Chem. 2002, 26, 178–188. [Google Scholar]
- Toebes, A.H.W.; de Boer, V.; Verkleij, J.A.C.; Lingeman, H.; Ernst, W.H.O. Extraction of Isoflavone Malonylglucosides from Trifolium pratense L. J. Agric. Food Chem. 2005, 53, 4660–4666. [Google Scholar] [CrossRef] [PubMed]
- Youssef, F.S.; Ashour, M.L.; El-Beshbishy, H.A.; Ahmed Hamza, A.; Singab, A.N.B.; Wink, M. Pinoresinol-4-O-β-D-glucopyranoside: A lignan from prunes (Prunus domestica) attenuates oxidative stress, hyperglycaemia and hepatic toxicity in vitro and in vivo. J. Pharm. Pharmacol. 2020, 72, 1830–1839. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Ichinose, K.; Takahashi, C.; Ho, F.-C.; Wu, J.-B.; Sankawa, U. Chemical studies on Sophora tomentosa: The isolation of a new class of isoflavonoid. Chem. Pharm. Bull. 1990, 38, 2756–2759. [Google Scholar] [CrossRef]
- Liang, Z.; Yang, M.; Xu, X.; Xie, Z.; Huang, J.; Li, X.; Yang, D. Isolation and purification of geniposide, crocin-1, and geniposidic acid from the fruit of Gardenia jasminoides Ellis by high-speed counter-current chromatography. Sep. Sci. Technol. 2014, 49, 1427–1433. [Google Scholar] [CrossRef]
- Forino, M.; Tartaglione, L.; Dell’Aversano, C.; Ciminiello, P. NMR-based identification of the phenolic profile of fruits of Lycium barbarum (goji berries). Isolation and structural determination of a novel N-feruloyl tyramine dimer as the most abundant antioxidant polyphenol of goji berries. Food Chem. 2016, 194, 1254–1259. [Google Scholar] [CrossRef]
- Pei, K.; Ou, J.; Huang, J.; Ou, S. p-Coumaric acid and its conjugates: Dietary sources, pharmacokinetic properties and biological activities. J. Sci. Food Agric. 2016, 96, 2952–2962. [Google Scholar] [CrossRef]
- Johnsson, P.; Peerlkamp, N.; Kamal-Eldin, A.; Andersson, R.E.; Andersson, R.; Lundgren, L.N.; Åman, P. Polymeric fractions containing phenol glucosides in flaxseed. Food Chem. 2002, 76, 207–212. [Google Scholar] [CrossRef]
- Terazawa, M.; Okuyama, H.; Miyake, M. Isolation of coniferin and syringin from the cambial tissue and inner-bark of some angiospermous woods. J. Jpn. Wood Res. Soc. 1984, 30, 409–412. [Google Scholar]
- Sticher, O.; Lahloub, M.F. Phenolic Glycosides of Paulownia tomentosa Bark. Planta Med. 1982, 46, 145–148. [Google Scholar] [CrossRef]
- Tram, N.T.N.; Mitova, M.; Bankova, V.; Handjieva, N.; Popov, S.S. GC-MS of Crinum latifolium L. alkaloids. Z. Nat. C 2002, 57, 239–242. [Google Scholar] [CrossRef] [Green Version]
- Zalkow, L.; Ekpo, B.; Gelbaum, L.; Harris III, R.; Keinan, E.; Novak Jr, J.; Ramming, C.; Van Derveer, D. The benzofurans of Isocoma wrightii. Structure and stereochemistry. J. Nat. Prod. 1979, 42, 203–219. [Google Scholar] [CrossRef] [PubMed]
- De Moura, N.F.; Simionatto, E.; Porto, C.; Hoelzel, S.C.S.; Dessoy, E.C.S.; Zanatta, N.; Morel, A.F. Quinoline Alkaloids, Coumarins and Volatile Constituents of Helietta longifoliata. Planta Med. 2002, 68, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.E.; de Arias, A.R.; Yaluff, G.; de Bilbao, N.V.; Nakayama, H.; Torres, S.; Schinini, A.; Guy, I.; Heinzen, H.; Fournet, A. Antileishmanial activity of furoquinolines and coumarins from Helietta apiculata. Phytomedicine 2010, 17, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.N.H.; Ho, W.S. Antiproliferative activity of gambogic acid isolated from Garcinia hanburyi in Hep3B and Huh7 cancer cells. Oncol. Rep. 2013, 29, 1744–1750. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.J.; Sousa, M.; Parola, A.J.; de Melo, J.S.S.; Catarino, F.; Marçalo, J.; Pina, F. Identification of 7, 4′-Dihydroxy-5-methoxyflavylium in “Dragon’s Blood”: To Be or Not to Be an Anthocyanin. Chem. Eur. J. 2007, 13, 1417–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Li, F.; Yan, X.; Cheng, Q.; Xue, R.; Yu, H.; Li, Z.; Wang, C. Phytochemical Constituents and Biological Activities of Plants from the Genus cissampelos. Chem. Biodivers. 2021, 18, e2100358. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, A.; Wójcicki, K.; Klensporf-Pawlik, D.; Marzec, M.; Lucarini, M.; Durazzo, A.; Fonseca, J.; Santini, A.; Nowak, I.; Souto, E.B. Cold-Pressed Pomegranate Seed Oil: Study of Punicic Acid Properties by Coupling of GC/FID and FTIR. Molecules 2022, 27, 5863. [Google Scholar] [CrossRef]
- Lannang, A.M.; Anjum, S.; Tangmouo, J.G.; Krohn, K.; Choudhary, M.I. Conessine isolated from Holarrhena floribunda. Acta Crystallogr. Sect. E Struct. Rep. Online 2007, 63, o4398. [Google Scholar] [CrossRef]
- Ikeda, T.; Udayama, M.; Okawa, M.; Arao, T.; Kinjo, J.; Nohara, T. Partial Hydrolysis of Soyasaponin I and the Hepatoprotective Effects of the Hydrolytic Products. Studies on the Hepatoprotective Drugs. Part IV. (Studies on the Constituents of the Leguminous Plants. Part LVII.) Study of the Structure-Hepatoprotective Relationship of Soyasapogenol B Analogs. Chem. Pharm. Bull. 1998, 46, 359–361. [Google Scholar] [CrossRef] [Green Version]
- Udayama, M.; Ohkawa, M.; Yoshida, N.; Kinjo, J.; Nohara, T. Structures of Three New Oleanene Glucuronides Isolated from Lathyrus palustris var. pilosus and Hepatoprotective Activity. Chem. Pharm. Bull. 1998, 46, 1412–1415. [Google Scholar] [CrossRef] [Green Version]
- Dwuma-Badu, D.; Ayim, J.S.; Mingle, C.; Tackie, A.; Slatkin, D.; Knapp, J.; Schiff, P., Jr. Alkaloids of Cissampelos pareira. Phytochemistry 1975, 14, 2520–2521. [Google Scholar] [CrossRef]
- Steyn, P.S.; Vleggaar, R.; Wessels, P.L.; Scott, D.B. Biosynthesis of versiconal acetate, versiconol acetate, and versiconol, metabolites from cultures of Aspergillus parasiticus treated with dichlorvos. The role of versiconal acetate in aflatoxin biosynthesis. J. Chem. Soc. Perkin Trans. 1 1979, 460–463. [Google Scholar] [CrossRef]
- Yao, R.C.; Hsieh, D.P. Step of dichlorvos inhibition in the pathway of aflatoxin biosynthesis. Appl. Microbiol. 1974, 28, 52–57. [Google Scholar] [CrossRef]
- Jadalla, B.M.I.S.; Moser, J.J.; Sharma, R.; Etsassala, N.G.E.R.; Egieyeh, S.A.; Badmus, J.A.; Marnewick, J.L.; Beukes, D.; Cupido, C.N.; Hussein, A.A. In Vitro Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities and Antioxidant Capacity of Helichrysum cymosum and Helichrysum pandurifolium Schrank Constituents. Separations 2022, 9, 190. [Google Scholar] [CrossRef]
- Ahmed, S.; Ali, C.; Ruma, R.A.; Mahmud, S.; Paul, G.K.; Saleh, A.; Alshahrani, M.M.; Obaidullah, A.J.; Biswas, S.K.; Rahman, M.; et al. Molecular Docking and Dynamics Simulation of Natural Compounds from Betel Leaves (Piper betle L.) for Investigating the Potential Inhibition of Alpha-Amylase and Alpha-Glucosidase of Type 2 Diabetes. Molecules 2022, 27, 4526. [Google Scholar] [CrossRef]
- Chikezie, P.C.; Ojiako, O.A.; Nwufo, K.C. Overview of anti-diabetic medicinal plants: The Nigerian research experience. J. Diabetes Metab. 2015, 6, 546. [Google Scholar] [CrossRef]
- Pereira, A.S.; Haan, H.D.; Peña-García, J.; Moreno, M.M.; Pérez-Sánchez, H.; Apostolides, Z. Exploring African Medicinal Plants for Potential Anti-Diabetic Compounds with the DIA-DB Inverse Virtual Screening Web Server. Molecules 2019, 24, 2002. [Google Scholar] [CrossRef] [Green Version]
- Akshatha, J.V.; SantoshKumar, H.S.; Prakash, H.S.; Nalini, M.S. In silico docking studies of α-amylase inhibitors from the anti-diabetic plant Leucas ciliata Benth. and an endophyte, Streptomyces longisporoflavus. 3 Biotech 2021, 11, 51. [Google Scholar] [CrossRef]
- Oliveira, H.; Fernandes, A.; Brás, N.F.; Mateus, N.; de Freitas, V.; Fernandes, I. Anthocyanins as antidiabetic agents—In vitro and in silico approaches of preventive and therapeutic effects. Molecules 2020, 25, 3813. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. Cheminform. J. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastman, P.; Friedrichs, M.S.; Chodera, J.D.; Radmer, R.J.; Bruns, C.M.; Ku, J.P.; Beauchamp, K.A.; Lane, T.J.; Wang, L.; Shukla, D.; et al. OpenMM 4: A Reusable, Extensible, Hardware Independent Library for High Performance Molecular Simulation. J. Chem. Theory Comput. 2013, 9, 461. [Google Scholar] [CrossRef] [PubMed]
- Dallakyan, S.; Olson, A.J. Chemical Biology; Hempel, J.E., Williams, C.H., Hong, C.C., Eds.; Springer: New York, NY, USA, 2015; pp. 243–250. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Tentative Identification | Rt | m/z | Formula | Ms/Ms-Fragmnets | Biological Source | Nature of Compounds | References |
|---|---|---|---|---|---|---|---|---|
| 1 | Epinorlycoramine | 1.2 | 275.152 | C16H21NO3 | 112.0874-124.0875-154.0978-190.098-202.0982-220.1087 | Narcissus leonensis plant | Alkaloid | [16] |
| 2 | Eburnamonine | 1.6 | 294.173 | C19H22N2O | 114.1028-145.0519 | Leaves of Kopsia zarutensi | Alkaloid | [17] |
| 3 | 3-Acetylnerbowdine | 2.0 | 361.153 | C19H23NO6 | 151.0774 | Bulbs of Nerine bowdenii | Alkaloid | [18,19] |
| 4 | Arborinine | 3.3 | 285.290 | C16H15NO4 | 110.0375-135.0299-153.0546 | Leaves of Glycosmis parva | Alkaloid | [20] |
| 5 | 1,2-Dehydroreticuline | 4.1 | 328.155 | C19H22NO4 | 120.0817-132.0821-166.0880-178.0875 | Bark and the root of Xylopia parviflora | Alkaloid | [21,22] |
| 6 | Versiconol acetate | 6.7 | 402.095 | C20H18O9 | 143.0711 | Culture of Aspergillus parasiticus | Anthraquinone | [23] |
| 7 | Karwinaphthol B | 7.0 | 288.136 | C17H20O4 | 112.0879-117.0333-138.0664-145.0289 | Roots of Karwinskia humboldtiana | Benzisochromans | [24] |
| 8 | Veraguensin | 7.4 | 372.194 | C22H28O5 | 177.0550-196.1444-222.1249 | Leaves and root bark of Nectandra turbacensis (Kunth) Nees | Lignan | [25,26] |
| 9 | Narirutin | 8.6 | 580.179 | C27H32O14 | 313.0713-415.1029-433.1137 | Citrus fruits | Flavanone glycoside | [27] |
| 10 | Pelargonidin 3-O-rutinoside | 8.6 | 579.171 | C27H31O14 | 313.0713-397.0918 415.1029-433.1137 | Strawberries | Anthocyanin | [28] |
| 11 | Boschnaloside | 8.8 | 344.147 | C16H24O8 | 151.0742-177.0546-186.0527 | Boschniakia rossica plant | Iridoid glycoside | [29,30] |
| 12 | Biochanin A-β-d-glucoside | 9.0 | 446.121 | C22H22O10 | 145.0288-175.0619-177.0551 | Trifolium pratense L. plant | Isoflavone glycoside | [31] |
| 13 | Pinoresinol glucoside | 9.1 | 520.194 | C26H32O11 | 177.0552-184.0717-186.0523-191.0710 | Prunes of Prunus domestica L. | Lignan | [32] |
| 14 | Sophora isoflavanone A | 9.2 | 370.142 | C21H22O6 | 145.0285-177.0553-284.0695 | Sophora tomentosa L. plant | Isoflavone | [33] |
| 15 | Geniposide | 9.2 | 388.137 | C17H24O10 | 149.0614-151.0390-177.0552-186.0541 | Fruit of Gardenia jasminoides Ellis | Iridoid glycoside | [34] |
| 16 | N-Feruloyltyramine | 9.3 | 313.131 | C18H19NO4 | 117.0339-145.0293-149.0607-162.0539 | Fruits of Lycium barbarum (goji berries) Bassia indica and A. alopecuroides plants | Alkaloid | [10,35] |
| 17 | 4-Hydroxycinnamyl alcohol 4-d-glucoside | 9.3 | 312.121 | C15H20O7 | 117.0339-145.0293-149.0607- | Linum usitatissimum, Linn. plant | Lignan | [36,37] |
| 18 | Coniferin | 9.4 | 342.131 | C16H22O8 | 137.0600-175.0763-177.0546-218.0794 | Paulownia tomentosa bark | Lignan | [38,39] |
| 19 | Powelline | 9.7 | 301.131 | C17H19NO4 | 121.0652-135.0448-163.0395-180.0660 | Leaves from Crinum latifolium L. | Alkaloid | [40] |
| 20 | Toxyl angelate | 9.7 | 300.136 | C18H20O4 | 121.0652-135.0448-145.0286-163.0395- | Isocoma wrightii plant | Banzofuran | [41] |
| 21 | Heliettin | 10.1 | 314.152 | C19H22O4 | 121.0657-145.0289-177.0553 | Stem bark of Helietta longifoliata Britt | Furanochomarine | [42,43] |
| 22 | Gambogic acid | 10.2 | 628.304 | C38H44O8 | 121.0655-177.0549-201.0549-297.1123 | Garcinia hanburyi plant | Phenolic acid (Xanthoid derivative) | [44] |
| 23 | Dracorubin | 10.9 | 488.162 | C32H24O5 | 121.0654-177.0565-201.0545-323.0921 | Resin extracted from the tree Dracaena draco | Proanthocyanidine | [45] |
| 24 | Isochondrodendrine | 12.1 | 594.273 | C36H38N2O6 | 565.2679 | Cissampelos mucronate and Cissampelos pareira plants | Alkaloids | [46] |
| 25 | Punicic acid | 13.0 | 278.225 | C18H30O2 | 107.0864-121.1017-133.1021-135.1160-149.1334 | Pomegranate Seed Oil | Unsaturated fatty acid | [47] |
| 26 | Conessine | 13.3 | 356.319 | C24H40N2 | 121.1015-123.1154-135.1162-149.1330 | Holarrhena floribunda G. Don. plant | Alkaloid | [48] |
| 27 | Soyasapogenol B 3-O-d-glucuronide | 13.5 | 634.408 | C36H58O9 | 133.0863-177.1134-247.2054-291.2317 | Aerial parts of Lathylus palustris L. | Triterpenoid saponin | [49,50] |
| No. | Tentative Identification | Binding Energy (kcal/mol) | |
|---|---|---|---|
| α-Amylase | α-Glucosidase | ||
| 1 | Epinorlycoramine | −7.6 | −6.2 |
| 2 | Eburnamonine | −8.7 | −6.4 |
| 3 | 3-Acetylnerbowdine | −7.2 | −6.6 |
| 4 | Arborinine | −7.5 | −6.0 |
| 5 | 1,2-Dehydroreticuline | −7.4 | −7.4 |
| 6 | Versiconol acetate | −7.5 | −7.1 |
| 7 | Karwinaphthol B | −7.4 | −5.9 |
| 8 | Veraguensin | −7.5 | −6.7 |
| 9 | Narirutin | −8.5 | −7.9 |
| 10 | Pelargonidin 3-O-rutinoside | −8.5 | −8.4 |
| 11 | Boschnaloside | −7.1 | −5.8 |
| 12 | Biochanin A-β-D-glucoside | −7.2 | −6.3 |
| 13 | Pinoresinol glucoside | −7.9 | −6.0 |
| 14 | Sophora isoflavanone A | −7.4 | −9.1 |
| 15 | Geniposide | −7.2 | −6.5 |
| 16 | N-Feruloyltyramine | −7.3 | −7.6 |
| 17 | 4-Hydroxycinnamyl alcohol 4-D-glucoside | −7.3 | −6.8 |
| 18 | Coniferin | −6.9 | −5.7 |
| 19 | Powelline | −6.7 | −6.5 |
| 20 | Toxyl angelate | −7.2 | −6.4 |
| 21 | Heliettin | −7.2 | −7.1 |
| 22 | Gambogic acid | −7.2 | −7.1 |
| 23 | Dracorubin | −7.8 | −8.3 |
| 24 | Isochondrodendrine | −9.1 | −7.0 |
| 25 | Punicic acid | −5.6 | −5.7 |
| 26 | Conessine | −4.8 | −5.6 |
| 27 | Soyasapogenol B 3-O-D-glucuronide | −4.5 | −4.7 |
| STD | Montbretin A | −8.1 | …. |
| Miglitol | …. | −8.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amin, E.; Abdel-Bakky, M.S.; Mohammed, H.A.; Hassan, M.H.A. Chemical Profiling and Molecular Docking Study of Agathophora alopecuroides. Life 2022, 12, 1852. https://doi.org/10.3390/life12111852
Amin E, Abdel-Bakky MS, Mohammed HA, Hassan MHA. Chemical Profiling and Molecular Docking Study of Agathophora alopecuroides. Life. 2022; 12(11):1852. https://doi.org/10.3390/life12111852
Chicago/Turabian StyleAmin, Elham, Mohamed Sadek Abdel-Bakky, Hamdoon A. Mohammed, and Marwa H. A. Hassan. 2022. "Chemical Profiling and Molecular Docking Study of Agathophora alopecuroides" Life 12, no. 11: 1852. https://doi.org/10.3390/life12111852
APA StyleAmin, E., Abdel-Bakky, M. S., Mohammed, H. A., & Hassan, M. H. A. (2022). Chemical Profiling and Molecular Docking Study of Agathophora alopecuroides. Life, 12(11), 1852. https://doi.org/10.3390/life12111852