
The Possible Role of the Type I Chaperonins in Human Insulin Self-Association

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Insulin Preparation and Amyloid Aggregation
2.3. Static and Dynamic Light Scattering
2.4. Circular Dichroism Spectroscopy
2.5. Non-Denaturing Gel
2.6. Thioflavin T Spectrofluorometric Measurements
2.7. Atomic Force Microscope (AFM)
3. Results
3.1. Zn2+ Insulin Self-Association in the Refolding Buffer
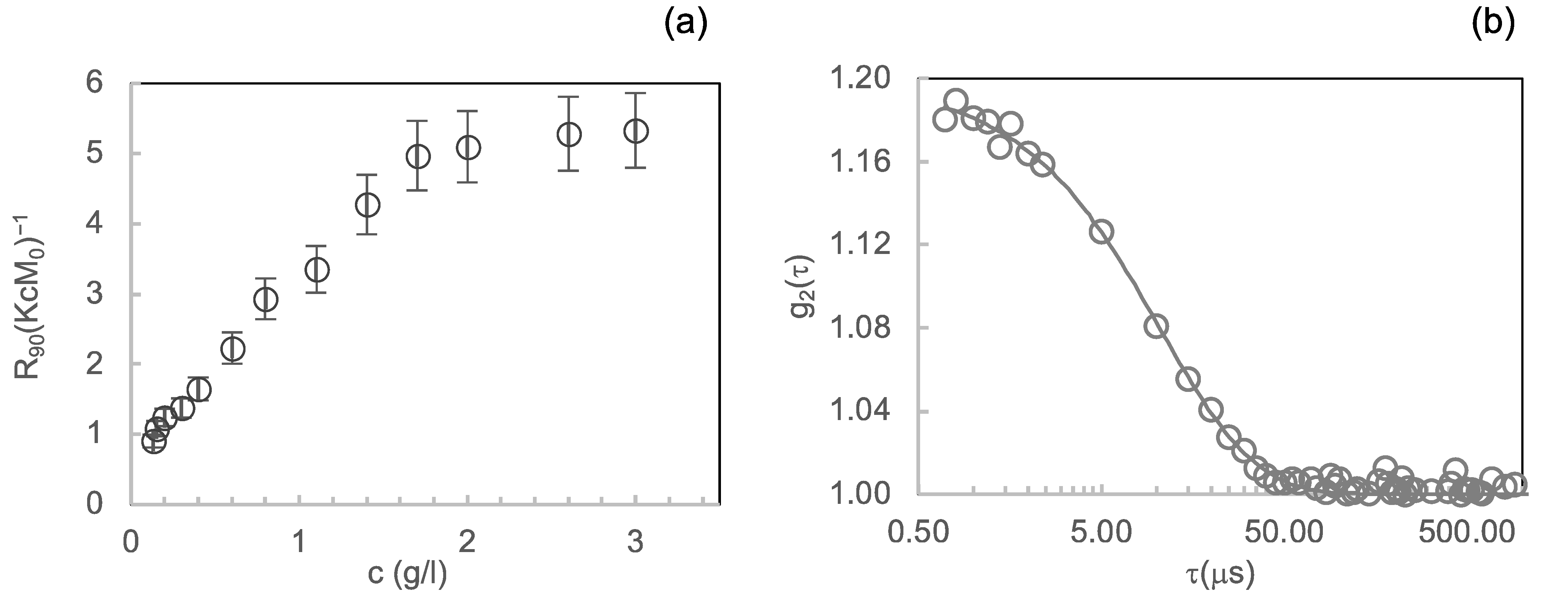
3.1.1. Light Scattering
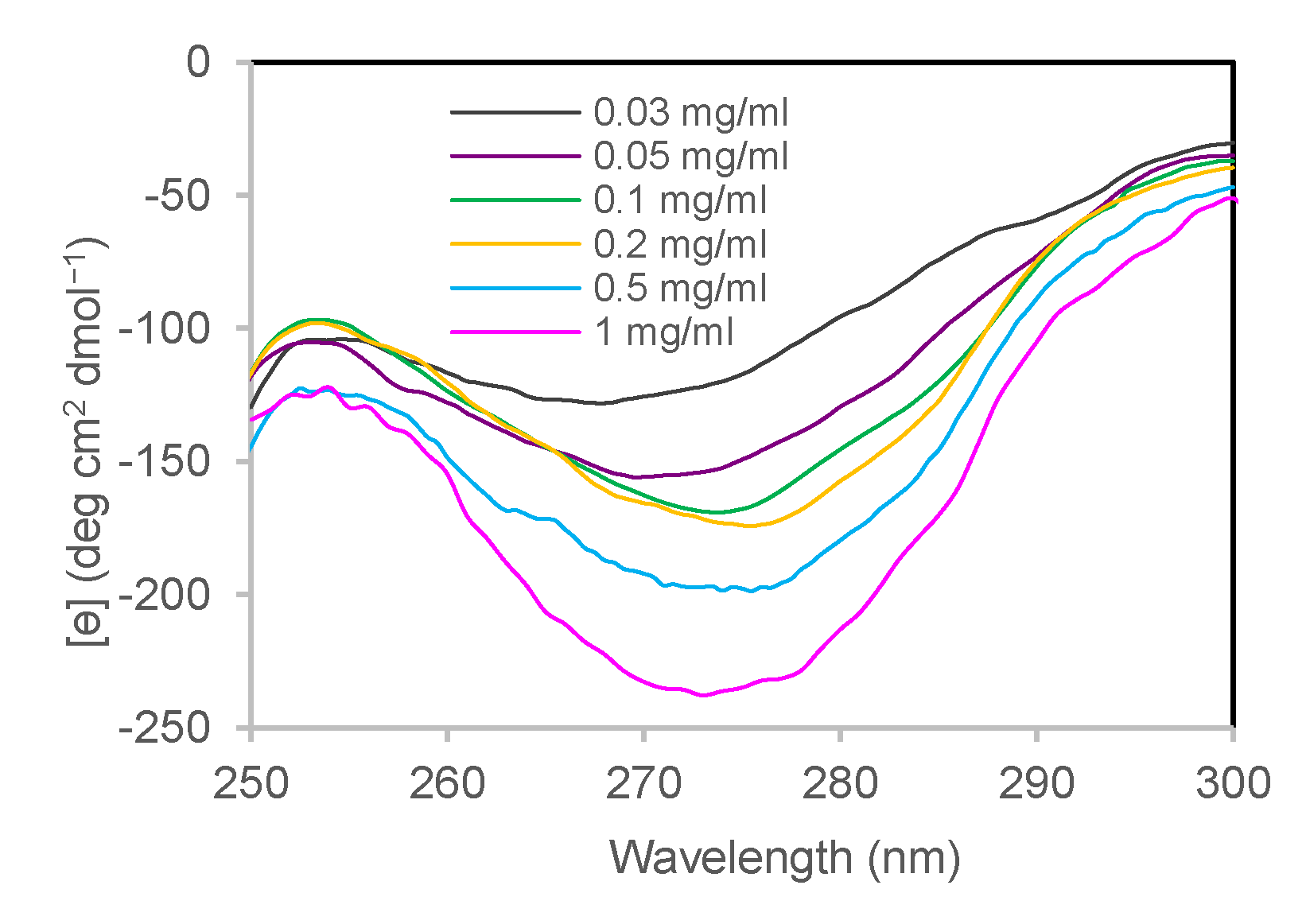
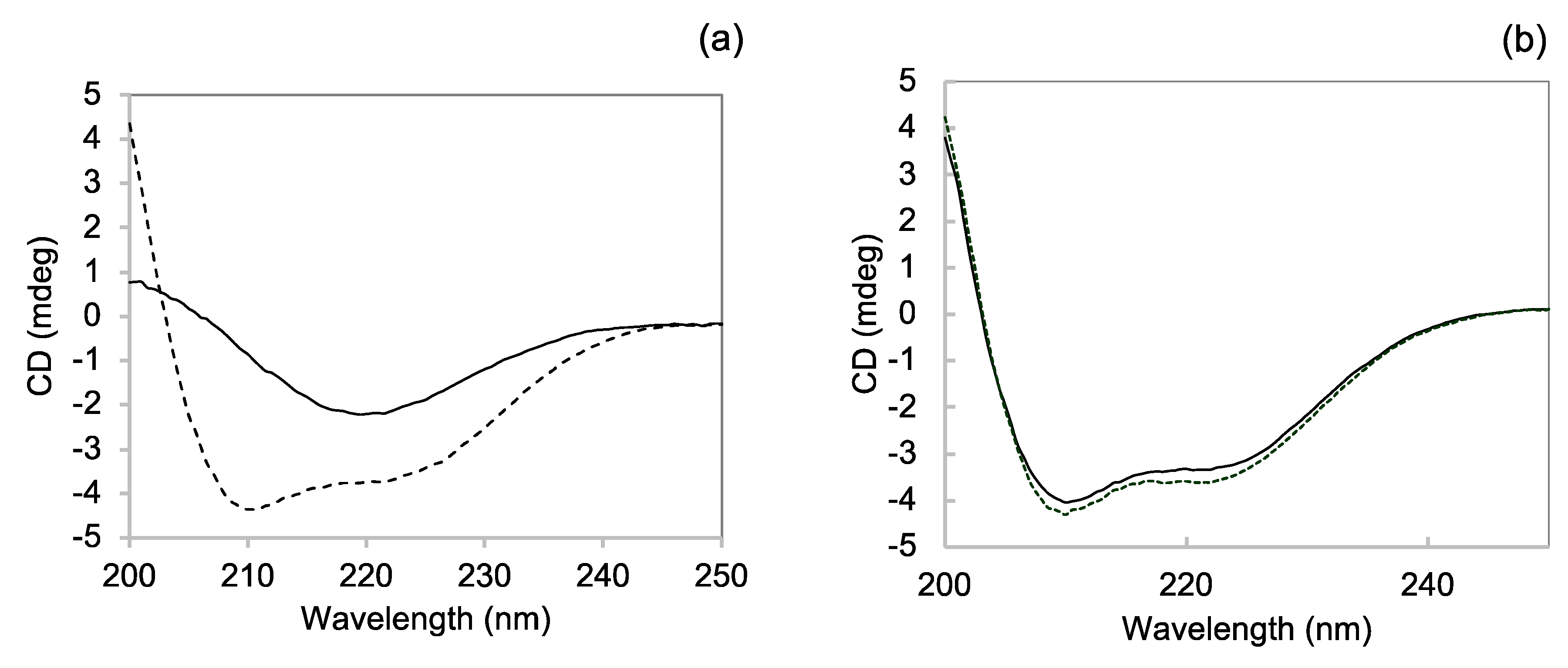
3.1.2. Near-UV CD
3.2. GroEL Effect on Zn2+ Insulin Self-Association
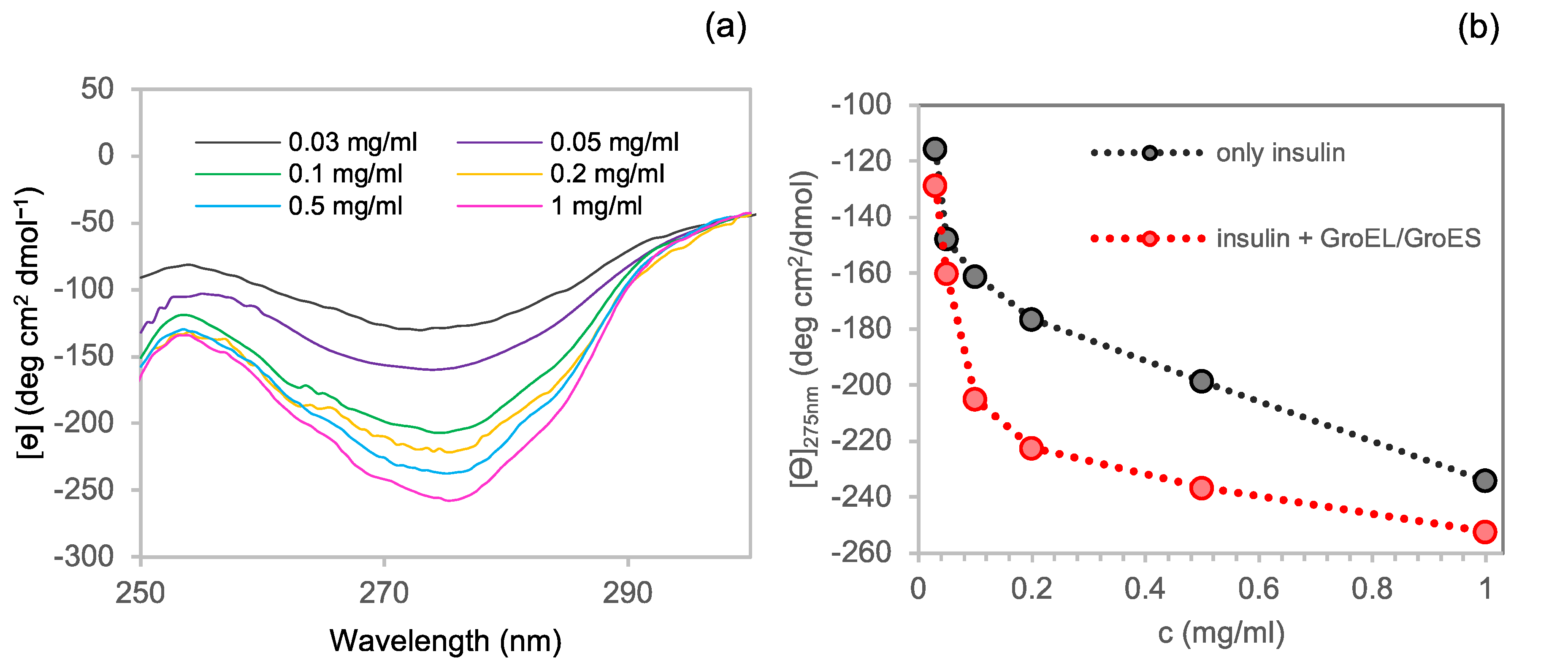
3.2.1. Near-UV CD
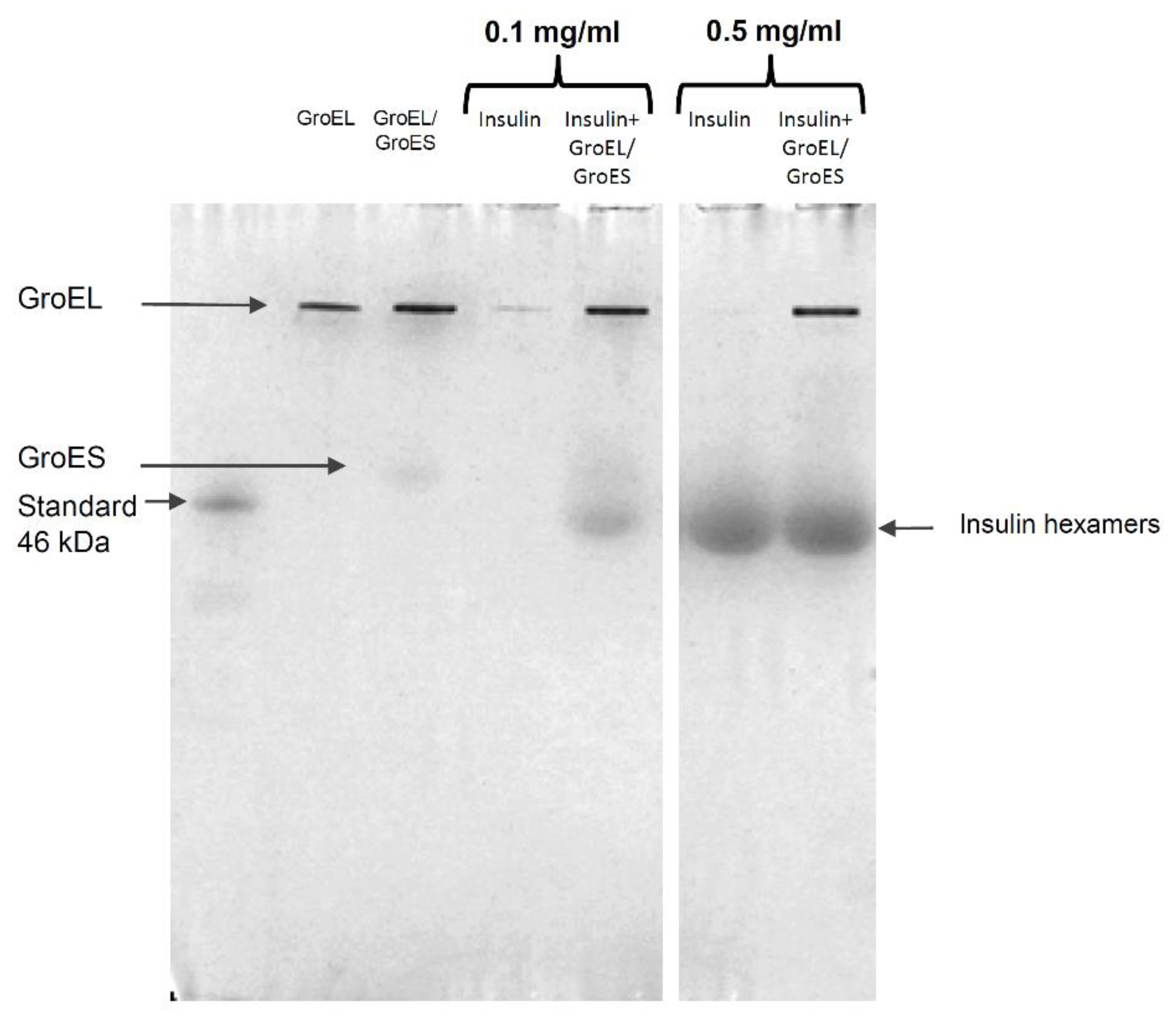
3.2.2. Native Electrophoresis
3.3. Influence of Chaperonins on Zn2+ Insulin Amyloid Formation at pH 7.4
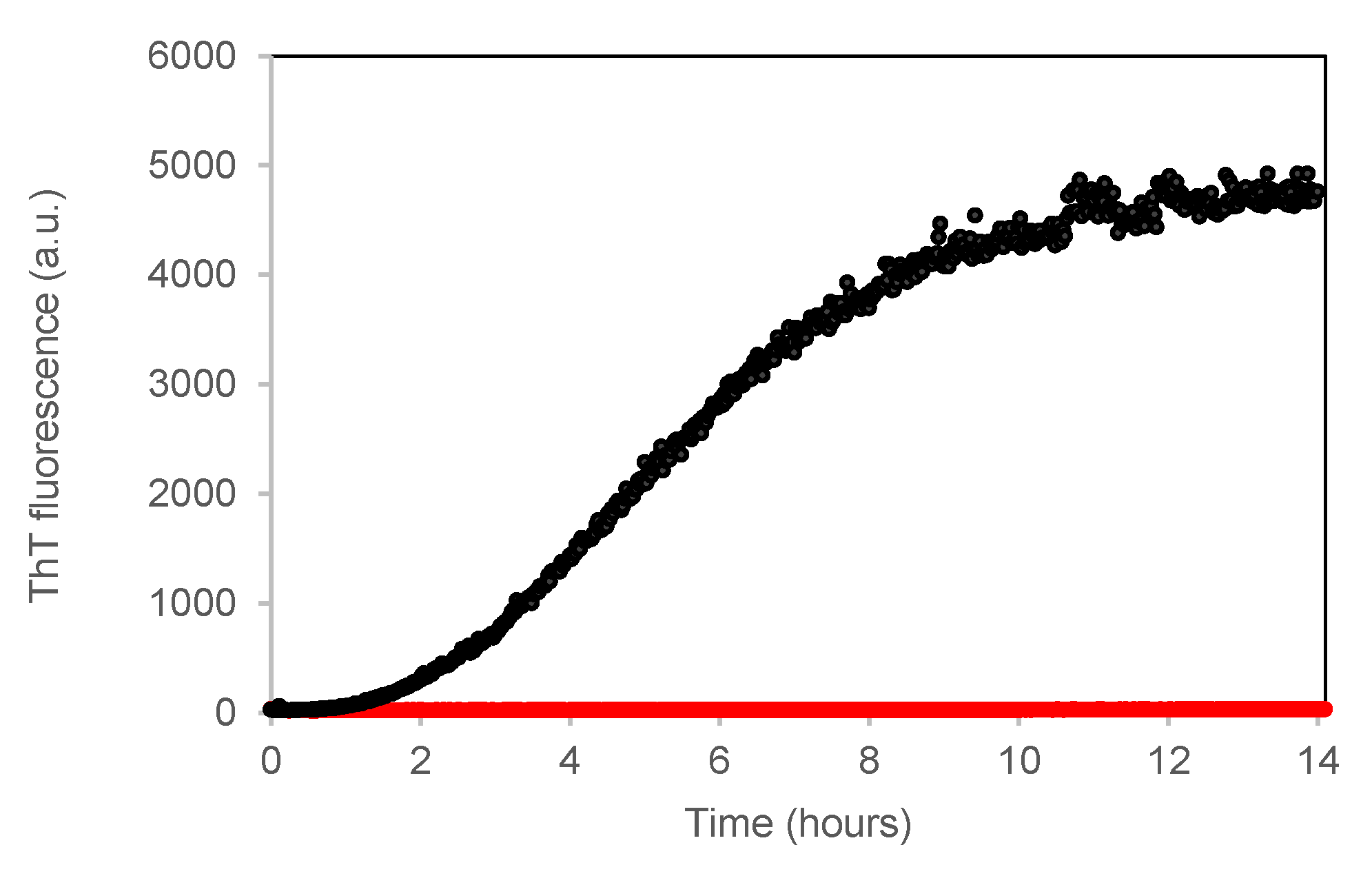
3.3.1. ThT Fluorescence Assay
3.3.2. Far-UV CD
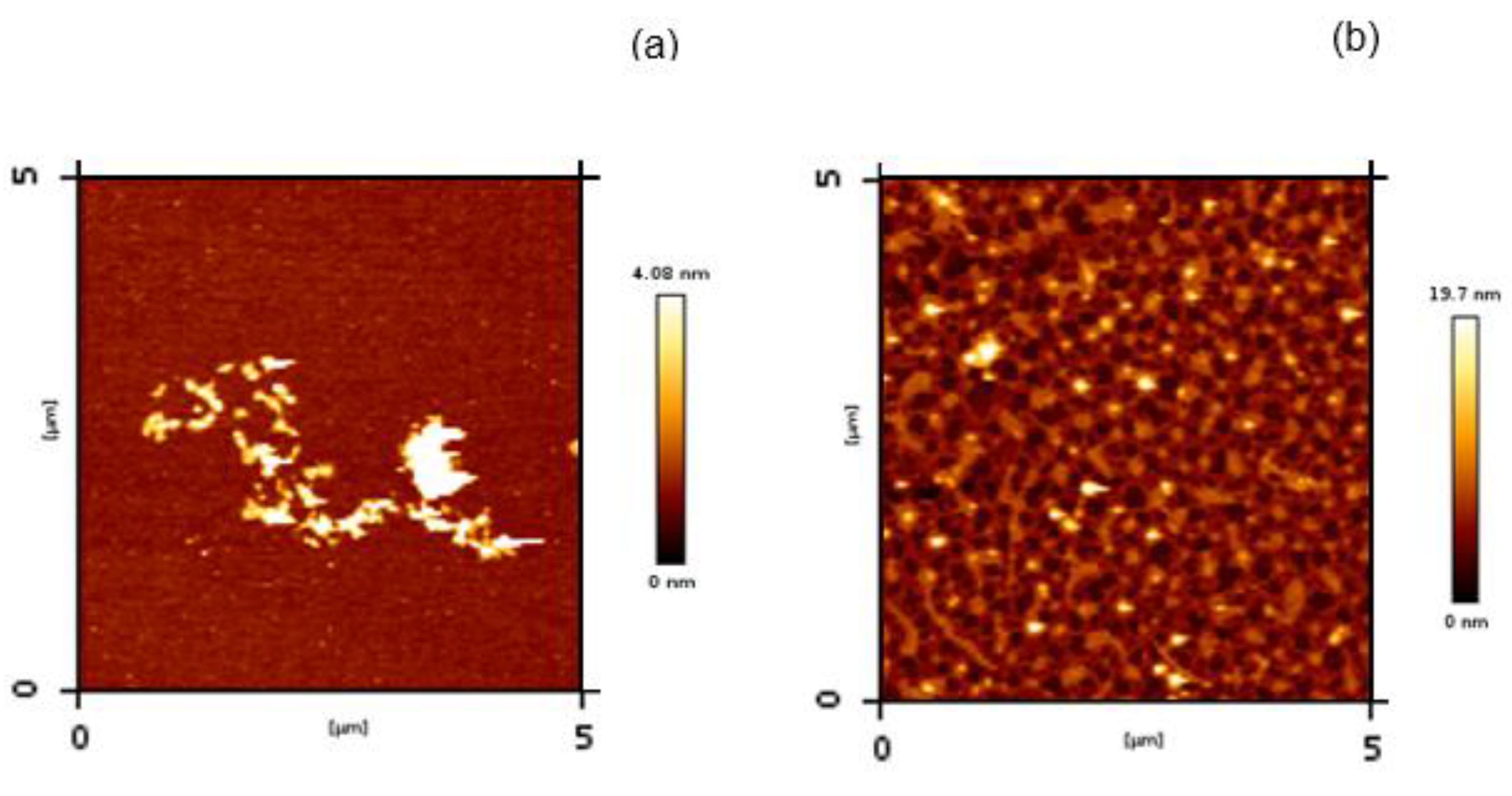
3.3.3. Atomic Force Microscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dodson, G.; Steiner, D. The role of assembly in insulin’s biosynthesis. Curr. Opin. Struct. Biol. 1998, 8, 189–194. [Google Scholar] [CrossRef]
- Adams, M.J.; Blundell, T.L.; Dodson, E.J.; Dodson, G.G.; Vijayan, M.; Baker, E.N.; Harding, M.M.; Hodgkin, D.C.; Rimmer, B.; Sheat, S. Structure of Rhombohedral 2 Zinc Insulin Crystals. Nature 1969, 224, 491–495. [Google Scholar] [CrossRef]
- Kiselar, J.G.; Datt, M.; Chance, M.R.; Weiss, M.A. Structural Analysis of Proinsulin Hexamer Assembly by Hydroxyl Radical Footprinting and Computational Modeling. J. Biol. Chem. 2011, 286, 43710–43716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, J.; Carroll, R.; Swift, H.H.; Steiner, D.F. Studies on the molecular organization of rat insulin secretory granules. J. Biol. Chem. 1987, 262, 16531–16535. [Google Scholar] [CrossRef]
- Hellman, U.; Wernstedt, C.; Westermark, P.; Obrien, T.D.; Rathbun, W.B.; Johnson, K.H. Amino-Acid-Sequence from Degu Islet Amyloid-Derived Insulin Shows Unique Sequence Characteristics. Biochem. Biophys. Res. Commun. 1990, 169, 571–577. [Google Scholar] [CrossRef]
- Brange, J. Galenics of Insulin: The Physico-Chemical and Pharmaceutical Aspects of Insulin and Insulin Preparations; Springer: Berlin/Heidelberg, Germany, 1987. [Google Scholar]
- Waugh, D.F.; Wilhelmson, D.F.; Commerford, S.L.; Sackler, M.L. Studies of the Nucleation and Growth Reactions of Selected Types of Insulin Fibrils. J. Am. Chem. Soc. 1953, 75, 2592–2600. [Google Scholar] [CrossRef]
- Attri, A.K.; Fernandez, C.; Minton, A.P. pH-dependent self-association of zinc-free insulin characterized by concentration-gradient static light scattering. Biophys. Chem. 2010, 148, 28–33. [Google Scholar] [CrossRef] [Green Version]
- Attri, A.K.; Fernandez, C.; Minton, A.P. Self-association of Zn-insulin at neutral pH: Investigation by concentration gradient-static and dynamic light scattering. Biophys. Chem. 2010, 148, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, M.R. Insulin amyloid at injection sites of patients with diabetes. Amyloid Int. J. Exp. Clin. Investig. 2016, 23, 139–147. [Google Scholar] [CrossRef]
- Manno, M.; Craparo, E.F.; Martorana, V.; Bulone, D.; San Biagio, P.L. Kinetics of insulin aggregation: Disentanglement of amyloid fibrillation from large-size cluster formation. Biophys. J. 2006, 90, 4585–4591. [Google Scholar] [CrossRef] [Green Version]
- Librizzi, F.; Rischel, C. The kinetic behavior of insulin fibrillation is determined by heterogeneous nucleation pathways. Protein Sci. 2005, 14, 3129–3134. [Google Scholar] [CrossRef] [PubMed]
- Fodera, V.; Librizzi, F.; Groenning, M.; van de Weert, M.; Leone, M. Secondary nucleation and accessible surface in insulin amyloid fibril formation. J. Phys. Chem. B 2008, 112, 3853–3858. [Google Scholar] [CrossRef]
- Fodera, V.; Cataldo, S.; Librizzi, F.; Pignataro, B.; Spiccia, P.; Leone, M. Self-organization pathways and spatial heterogeneity in insulin amyloid fibril formation. J. Phys. Chem. B 2009, 113, 10830–10837. [Google Scholar] [CrossRef]
- Nielsen, L.; Khurana, R.; Coats, A.; Frokjaer, S.; Brange, J.; Vyas, S.; Uversky, V.N.; Fink, A.L. Effect of environmental factors on the kinetics of insulin fibril formation: Elucidation of the molecular mechanism. Biochemistry 2001, 40, 6036–6046. [Google Scholar] [CrossRef]
- Podesta, A.; Tiana, G.; Milani, P.; Manno, M. Early events in insulin fibrillization studied by time-lapse atomic force microscopy. Biophys. J. 2006, 90, 589–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manno, M.; Craparo, E.F.; Podesta, A.; Bulone, D.; Carrotta, R.; Martorana, V.; Tiana, G.; San Biagio, P.L. Kinetics of Different Processes in Human Insulin Amyloid Formation (vol 366, pg 258, 2007). J. Mol. Biol. 2011, 406, 354. [Google Scholar] [CrossRef]
- Manno, M.; Giacomazza, D.; Newman, J.; Martorana, V.; San Biagio, P.L. Amyloid Gels: Precocious Appearance of Elastic Properties during the Formation of an Insulin Fibrillar Network. Langmuir 2010, 26, 1424–1426. [Google Scholar] [CrossRef]
- Martorana, V.; Raccosta, S.; Giacomazza, D.; Ditta, L.A.; Noto, R.; San Biagio, P.L.; Manno, M. Amyloid jams: Mechanical and dynamical properties of an amyloid fibrillar network. Biophys. Chem. 2019, 253, 106231. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.; Frokjaer, S.; Brange, J.; Uversky, V.N.; Fink, A.L. Probing the mechanism of insulin fibril formation with insulin mutants. Biochemistry 2001, 40, 8397–8409. [Google Scholar] [CrossRef]
- Bernson, D.; Mecinovic, A.; Abed, M.T.; Lime, F.; Jageland, P.; Palmlof, M.; Esbjorner, E.K. Amyloid formation of bovine insulin is retarded in moderately acidic pH and by addition of short-chain alcohols. Eur. Biophys. J. 2020, 49, 145–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias, A.E.; Velez-Granell, C.S.; Mayer, G.; Bendayan, M. Colocalization of chaperone Cpn60, proinsulin and convertase PC1 within immature secretory granules of insulin-secreting cells suggests a role for Cpn60 in insulin processing. J. Cell Sci. 2000, 113 Pt 11, 2075–2083. [Google Scholar] [CrossRef] [PubMed]
- Soltys, B.J.; Gupta, R.S. Immunoelectron microscopic localization of the 60-kDa heat shock chaperonin protein (Hsp60) in mammalian cells. Exp. Cell Res. 1996, 222, 16–27. [Google Scholar] [CrossRef]
- Soltys, B.J.; Gupta, R.S. Mitochondrial molecular chaperones hsp60 and mhsp70: Are their role restricted to mitochondria. In Stress Proteins; Latchman, D.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1998; pp. 69–100. [Google Scholar]
- Kleinridders, A.; Lauritzen, H.P.M.M.; Ussar, S.; Christensen, J.H.; Mori, M.A.; Bross, P.; Kahn, C.R. Leptin regulation of Hsp60 impacts hypothalamic insulin signaling. J. Clin. Investig. 2013, 123, 4667–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juwono, J.; Martinus, R.D. Does Hsp60 Provide a Link between Mitochondrial Stress and Inflammation in Diabetes Mellitus? J. Diabetes Res. 2016, 2016, 8017571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.Y.; Hartl, F.U.; Horwich, A.L. The mitochondrial chaperonin hsp60 is required for its own assembly. Nature 1990, 348, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.Y.; Hartl, F.U.; Martin, J.; Pollock, R.A.; Kalousek, F.; Neupert, W.; Hallberg, E.M.; Hallberg, R.L.; Horwich, A.L. Mitochondrial Heat-Shock Protein Hsp60 Is Essential for Assembly of Proteins Imported into Yeast Mitochondria. Nature 1989, 337, 620–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilasi, S.; Bulone, D.; Caruso Bavisotto, C.; Campanella, C.; Marino Gammazza, A.; San Biagio, P.L.; Cappello, F.; Conway de Macario, E.; Macario, A.J.L. Chaperonin of Group I: Oligomeric Spectrum and Biochemical and Biological Implications. Front. Mol. Biosci. 2017, 4, 99. [Google Scholar] [CrossRef] [PubMed]
- Horwich, A.L. Protein folding in the cell: An inside story. Nat. Med. 2011, 17, 1211–1216. [Google Scholar] [CrossRef]
- Vilasi, S.; Carrotta, R.; Ricci, C.; Rappa, G.C.; Librizzi, F.; Martorana, V.; Ortore, M.G.; Mangione, M.R. Inhibition of A beta(1-42) Fibrillation by Chaperonins: Human Hsp60 Is a Stronger Inhibitor than Its Bacterial Homologue GroEL. ACS Chem. Neurosci. 2019, 10, 3565–3574. [Google Scholar] [CrossRef]
- Mangione, M.R.; Vilasi, S.; Marino, C.; Librizzi, F.; Canale, C.; Spigolon, D.; Bucchieri, F.; Fucarino, A.; Passantino, R.; Cappello, F.; et al. Hsp60, amateur chaperone in amyloid-beta fibrillogenesis. BBA-Gen. Subj. 2016, 1860, 2474–2483. [Google Scholar] [CrossRef]
- Walti, M.A.; Steiner, J.; Meng, F.J.; Chung, H.S.; Louis, J.M.; Ghirlando, R.; Tugarinov, V.; Nath, A.; Clore, G.M. Probing the mechanism of inhibition of amyloid-beta(1-42)-induced neurotoxicity by the chaperonin GroEL. Proc. Natl. Acad. Sci. USA 2018, 115, E11924–E11932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricci, C.; Maccarini, M.; Falus, P.; Librizzi, F.; Mangione, M.R.; Moran, O.; Ortore, M.G.; Schweins, R.; Vilasi, S.; Carrotta, R. Amyloid beta-Peptides Interaction with Membranes: Can Chaperones Change the Fate? J. Phys. Chem. B 2019, 123, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.S.; Chuang, D.T. Mechanisms for GroEL/GroES-mediated folding of a large 86-kDa fusion polypeptide in vitro. J. Biol. Chem. 1999, 274, 10405–10412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusey, P.N. Introduction to scattering experiments. In Neutrons, X-rays, and Light: Scattering Methods Applied to Soft Condensed Matter; Linder, P., Zemb, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2002; pp. 3–22. [Google Scholar]
- Berne, B.J.; Pecora, R. Dynamic Light Scattering; John Wiley: New York, NY, USA, 1976; p. 376. [Google Scholar]
- Frisken, B.J. Revisiting the method of cumulants for the analysis of dynamic light-scattering data. Appl. Opt. 2001, 40, 4087–4091. [Google Scholar] [CrossRef] [Green Version]
- Noto, R.; Santangelo, M.G.; Ricagno, S.; Mangione, M.R.; Levantino, M.; Pezzullo, M.; Martorana, V.; Cupane, A.; Bolognesi, M.; Manno, M. The Tempered Polymerization of Human Neuroserpin. PLoS ONE 2012, 7, e32444. [Google Scholar] [CrossRef] [PubMed]
- Mawhinney, M.T.; Williams, T.L.; Hart, J.L.; Taheri, M.L.; Urbanc, B. Elucidation of insulin assembly at acidic and neutral pH: Characterization of low molecular weight oligomers. Proteins-Struct. Funct. Bioinform. 2017, 85, 2096–2110. [Google Scholar] [CrossRef]
- Frankaer, C.G.; Sonderby, P.; Bang, M.B.; Mateiu, R.V.; Groenning, M.; Bukrinski, J.; Harris, P. Insulin fibrillation: The influence and coordination of Zn2+. J. Struct. Biol. 2017, 199, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Sirangelo, I.; Borriello, M.; Vilasi, S.; Iannuzzi, C. Hydroxytyrosol Inhibits Protein Oligomerization and Amyloid Aggregation in Human Insulin. Int. J. Mol. Sci. 2020, 21, 4636. [Google Scholar] [CrossRef]
- Kadima, W.; Ogendal, L.; Bauer, R.; Kaarsholm, N.; Brodersen, K.; Hansen, J.F.; Porting, P. The Influence of Ionic-Strength and Ph on the Aggregation Properties of Zinc-Free Insulin Studied by Static and Dynamic Laser-Light Scattering. Biopolymers 1993, 33, 1643–1657. [Google Scholar] [CrossRef]
- Hvidt, S. Insulin Association in Neutral Solutions Studied by Light-Scattering. Biophys. Chem. 1991, 39, 205–213. [Google Scholar] [CrossRef]
- Uversky, V.N.; Garriques, L.N.; Millett, I.S.; Frokjaer, S.; Brange, J.; Doniach, S.; Fink, A.L. Prediction of the association state of insulin using spectral parameters. J. Pharm. Sci. 2003, 92, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Strickland, E.H.; Mercola, D. Near-Ultraviolet Tyrosyl Circular-Dichroism of Pig Insulin Monomers, Dimers, and Hexamers—Dipole-Dipole Coupling Calculations in Monopole Approximation. Biochemistry 1976, 15, 3875–3884. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, M.J.; Timasheff, S.N. Optical Activity of Insulin. 1. Nature of Circular Dichroism Bands. Biochemistry 1971, 10, 824–831. [Google Scholar] [PubMed]
- Librizzi, F.; Carrotta, R.; Spigolon, D.; Bulone, D.; San Biagio, P.L. alpha-Casein Inhibits Insulin Amyloid Formation by Preventing the Onset of Secondary Nucleation Processes. J. Phys. Chem. Lett. 2014, 5, 3043–3048. [Google Scholar] [CrossRef] [PubMed]
- Vilasi, S.; Carrotta, R.; Mangione, M.R.; Campanella, C.; Librizzi, F.; Randazzo, L.; Martorana, V.; Gammazza, A.M.; Ortore, M.G.; Vilasi, A.; et al. Human Hsp60 with Its Mitochondrial Import Signal Occurs in Solution as Heptamers and Tetradecamers Remarkably Stable over a Wide Range of Concentrations. PLoS ONE 2014, 9, e97657. [Google Scholar] [CrossRef] [Green Version]
- Ricci, C.; Carrotta, R.; Rappa, G.C.; Mangione, M.R.; Librizzi, F.; San Biagio, P.L.; Amenitsch, H.; Ortore, M.G.; Vilasi, S. Investigation Investigation on different chemical stability of mitochondrial Hsp60 and its precursor. Biophys. Chem. 2017, 229, 31–38. [Google Scholar] [CrossRef]
- Ricci, C.; Ortore, M.G.; Vilasi, S.; Carrotta, R.; Mangione, M.R.; Bulone, D.; Librizzi, F.; Spinozzi, F.; Burgio, G.; Amenitsch, H.; et al. Stability and disassembly properties of human naive Hsp60 and bacterial GroEL chaperonins. Biophys. Chem. 2016, 208, 68–75. [Google Scholar] [CrossRef]
- Iannuzzi, C.; Borriello, M.; Portaccio, M.; Irace, G.; Sirangelo, I. Insights into Insulin Fibril Assembly at Physiological and Acidic pH and Related Amyloid Intrinsic Fluorescence. Int. J. Mol. Sci. 2017, 18, 2551. [Google Scholar] [CrossRef] [Green Version]
- Jayamani, J.; Shanmugam, G. Gallic acid, one of the components in many plant tissues, is a potential inhibitor for insulin amyloid fibril formation. Eur. J. Med. Chem. 2014, 85, 352–358. [Google Scholar] [CrossRef]
- Grudzielanek, S.; Jansen, R.; Winter, R. Solvational tuning of the unfolding, aggregation and amyloidogenesis of insulin. J. Mol. Biol. 2005, 351, 879–894. [Google Scholar] [CrossRef]
- Clare, D.K.; Vasishtan, D.; Stagg, S.; Quispe, J.; Farr, G.W.; Topf, M.; Horwich, A.L.; Saibil, H.R. ATP-Triggered conformational changes delineate substrate-binding and -folding mechanics of the GroELchaperonin. Cell 2012, 149, 113–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, J.A.; Demeler, B.; Horowitz, P.M. Alteration of the quaternary structure of cpn60 modulates chaperonin- assisted folding. Implications for the mechanism of chaperonin action. J. Biol. Chem. 1994, 269, 2447–2451. [Google Scholar] [CrossRef]
- Soltys, B.J.; Gupta, R.S. Mitrochondrial proteins at unexpected cellular locations: Export of proteins from mitochondria from an evolutionary perspective. Int. Rev. Cytol. 1999, 194, 133–196. [Google Scholar]
- Ziaunys, M.; Sneideris, T.; Smirnovas, V. Self-inhibition of insulin amyloid-like aggregation. Phys. Chem. Chem. Phys. 2018, 20, 27638–27645. [Google Scholar] [CrossRef]
- Yagi-Utsumi, M.; Kunihara, T.; Nakamura, T.; Uekusa, Y.; Makabe, K.; Kuwajima, K.; Kato, K. NMR characterization of the interaction of GroEL with amyloid β as a model ligand. FEBS Lett. 2013, 587, 1605–1609. [Google Scholar] [CrossRef]
- Ansari, M.Y.; Mande, S.C. A Glimpse Into the Structure and Function of Atypical Type I Chaperonins. Front. Mol. Biosci. 2018, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Weiss, M.A. The Structure and Function of Insulin: Decoding the Tr Transition. Vitam. Horm. 2009, 80, 33–49. [Google Scholar]
- Palivec, V.; Viola, C.M.; Kozak, M.; Ganderton, T.R.; Krizkova, K.; Turkenburg, J.P.; Haluskova, P.; Zakova, L.; Jiracek, J.; Jungwirth, P.; et al. Computational and structural evidence for neurotransmitter-mediated modulation of the oligomeric states of human insulin in storage granules. J. Biol. Chem. 2017, 292, 8342–8355. [Google Scholar] [CrossRef] [Green Version]
- Kosinova, L.; Veverka, V.; Novotna, P.; Collinsova, M.; Urbanova, M.; Moody, N.R.; Turkenburg, J.P.; Jiracek, J.; Brzozowski, A.M.; Zakova, L. Insight into the Structural and Biological Relevance of the T/R Transition of the N-Terminus of the B-Chain in Human Insulin. Biochemistry 2014, 53, 3392–3402. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pizzo, F.; Mangione, M.R.; Librizzi, F.; Manno, M.; Martorana, V.; Noto, R.; Vilasi, S. The Possible Role of the Type I Chaperonins in Human Insulin Self-Association. Life 2022, 12, 448. https://doi.org/10.3390/life12030448
Pizzo F, Mangione MR, Librizzi F, Manno M, Martorana V, Noto R, Vilasi S. The Possible Role of the Type I Chaperonins in Human Insulin Self-Association. Life. 2022; 12(3):448. https://doi.org/10.3390/life12030448
Chicago/Turabian StylePizzo, Federica, Maria Rosalia Mangione, Fabio Librizzi, Mauro Manno, Vincenzo Martorana, Rosina Noto, and Silvia Vilasi. 2022. "The Possible Role of the Type I Chaperonins in Human Insulin Self-Association" Life 12, no. 3: 448. https://doi.org/10.3390/life12030448
APA StylePizzo, F., Mangione, M. R., Librizzi, F., Manno, M., Martorana, V., Noto, R., & Vilasi, S. (2022). The Possible Role of the Type I Chaperonins in Human Insulin Self-Association. Life, 12(3), 448. https://doi.org/10.3390/life12030448