
Weight Loss and Fat Metabolism during Multi-Day High-Altitude Sojourns: A Hypothesis Based on Adipocyte Signaling





Abstract
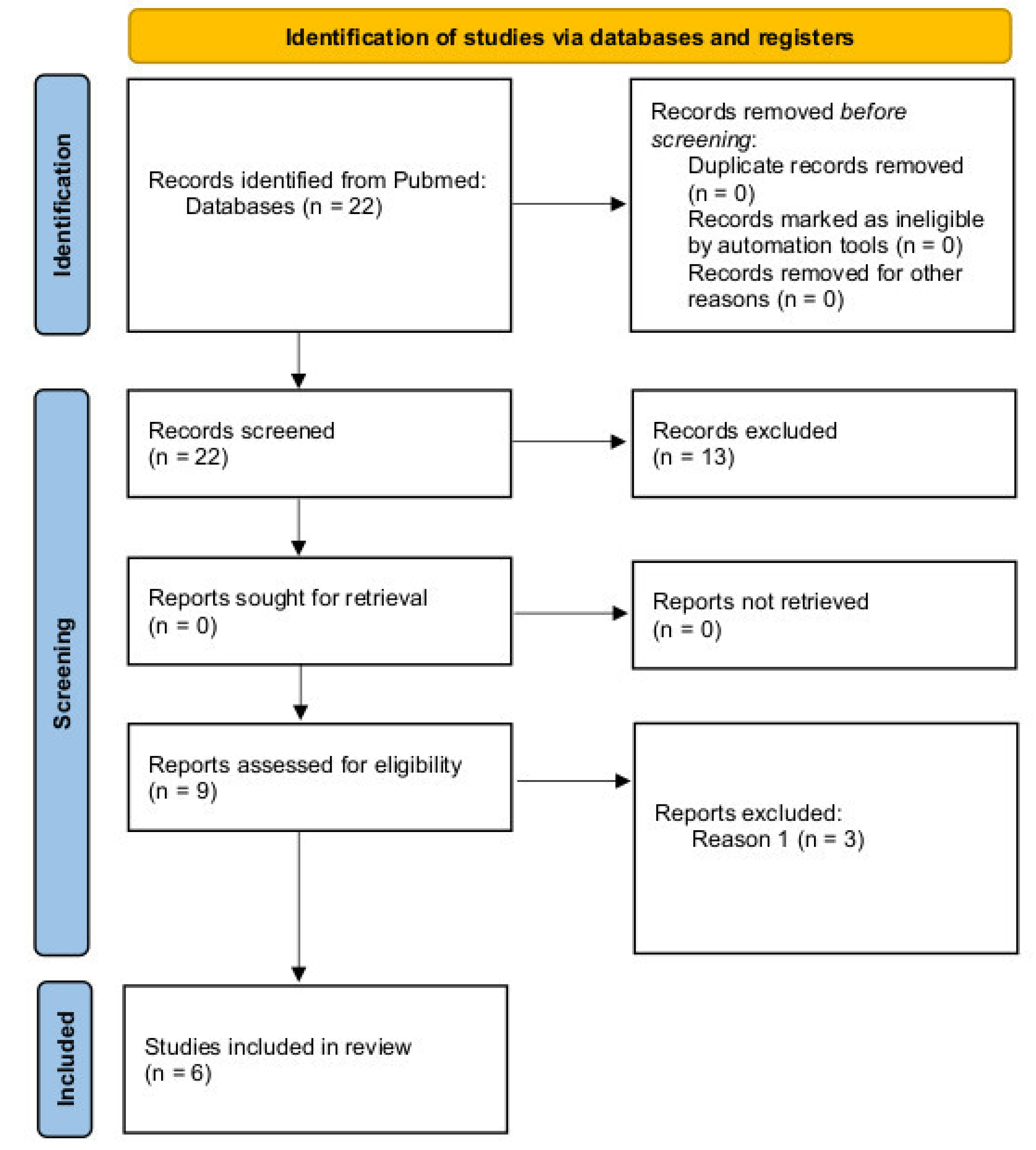
:1. Introduction
2. Materials and Methods
3. Results
3.1. Hormonal Appetite Reduction
3.2. Exercise-Induced Appetite Reduction
3.3. Energy Expenditure and Related Fuel Utilization
3.4. Genetic Adaptions to Hypoxia and Long-Term Sojourns
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kayser, B. Nutrition and energetics of exercise at altitude. Theory and possible practical implications. Sports Med. 1994, 17, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Westerterp, K.R.; Kayser, B. Body mass regulation at altitude. Eur. J. Gastroenterol. Hepatol. 2006, 18, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Westerterp-Plantenga, M.S.; Westerterp, K.R.; Rubbens, M.; Verwegen, C.R.; Richelet, J.P.; Gardette, B. Appetite at “high altitude” Operation Everest III (Comex-’97): A simulated ascent of Mount Everest. J. Appl. Physiol. 1999, 87, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Tschöp, M.; Strasburger, C.J.; Hartmann, G.; Biollaz, J.; Bärtsch, P. Raised leptin concentrations at high altitude associated with loss of appetite. Lancet 1998, 352, 1119–1120. [Google Scholar] [CrossRef]
- Tschöp, M.; Morrison, K.M. Weight loss at high altitude. In Hypoxia; Springer: Boston, MA, USA, 2001; pp. 237–247. [Google Scholar]
- Nair, C.S.; Malhotra, M.S.; Gopinath, P.M. Effect of altitude and cold acclimatisation on the basal metabolism in man. Aerosp. Med. 1971, 42, 1056–1059. [Google Scholar] [PubMed]
- Mekjavic, I.B.; Amon, M.; Kölegård, R.; Kounalakis, S.N.; Simpson, L.; Eiken, O.; Keramidas, M.E.; Macdonald, I.A. The Effect of Normobaric Hypoxic Confinement on Metabolism, Gut Hormones, and Body Composition. Front. Physiol. 2016, 7, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Z.; Zang, Y.; Hu, Y. Normobaric hypoxia training causes more weight loss than normoxia training after a 4-week residential camp for obese young adults. Sleep Breath. 2014, 18, 591–597. [Google Scholar] [CrossRef]
- Braun, B. Effects of high altitude on substrate use and metabolic economy: Cause and effect? Med. Sci. Sports Exerc. 2008, 40, 1495–1500. [Google Scholar] [CrossRef]
- McClelland, G.B.; Hochachka, P.W.; Weber, J.M. Carbohydrate utilization during exercise after high-altitude acclimation: A new perspective. Proc. Natl. Acad. Sci. USA 1998, 95, 10288–10293. [Google Scholar] [CrossRef] [Green Version]
- Sawka, M.N.; Cheuvront, S.N.; Kenefick, R.W. Hypohydration and Human Performance: Impact of Environment and Physiological Mechanisms. Sports Med. 2015, 45, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Dünnwald, T.; Gatterer, H.; Faulhaber, M.; Arvandi, M.; Schobersberger, W. Body Composition and Body Weight Changes at Different Altitude Levels: A Systematic Review and Meta-Analysis. Front. Physiol. 2019, 10, 430. [Google Scholar] [CrossRef] [PubMed]
- Palmer, B.F.; Clegg, D.J. Ascent to altitude as a weight loss method: The good and bad of hypoxia inducible factor activation. Obesity 2014, 22, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Murray, A.J.; Montgomery, H.E. How wasting is saving: Weight loss at altitude might result from an evolutionary adaptation. Bioessays 2014, 36, 721–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, S.J.; Blume, F.D. Weight loss and changes in body composition at high altitude. J. Appl. Physiol. 1984, 57, 1580–1585. [Google Scholar] [CrossRef]
- Wing-Gaia, S.L. Nutritional strategies for the preservation of fat free mass at high altitude. Nutrients 2014, 6, 665–681. [Google Scholar] [CrossRef] [Green Version]
- Rausch, L.K.; Hofer, M.; Pramsohler, S.; Kaser, S.; Ebenbichler, C.; Haacke, S.; Gatterer, H.; Netzer, N.C. Adiponectin, Leptin and Visfatin in Hypoxia and its Effect for Weight Loss in Obesity. Front. Endocrinol. 2018, 9, 615. [Google Scholar] [CrossRef]
- Netzer, N.C.; Chytra, R.; Küpper, T. Low intense physical exercise in normobaric hypoxia leads to more weight loss in obese people than low intense physical exercise in normobaric sham hypoxia. Sleep Breath. 2008, 12, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Hamad, N.; Travis, S.P.L. Weight loss at high altitude: Pathophysiology and practical implications. Eur. J. Gastroenterol. Hepatol. 2006, 18, 5–10. [Google Scholar] [CrossRef]
- Anvari, G.; Bellas, E. Hypoxia induces stress fiber formation in adipocytes in the early stage of obesity. Sci. Rep. 2021, 11, 21473. [Google Scholar] [CrossRef]
- McLaughlin, T.; Schnittger, I.; Nagy, A.; Zanley, E.; Xu, Y.; Song, Y.; Nieman, K.; Tremmel, J.A.; Dey, D.; Boyd, J.; et al. Relationship Between Coronary Atheroma, Epicardial Adipose Tissue Inflammation, and Adipocyte Differentiation Across the Human Myocardial Bridge. J. Am. Heart Assoc. 2021, 10, e021003. [Google Scholar] [CrossRef]
- Pescador, N.; Francisco, V.; Vázquez, P.; Esquinas, E.M.; González-Páramos, C.; Valdecantos, M.P.; García-Martínez, I.; Urrutia, A.A.; Ruiz, L.; Escalona-Garrido, C.; et al. Metformin reduces macrophage HIF1α-dependent proinflammatory signaling to restore brown adipocyte function in vitro. Redox Biol. 2021, 48, 102171. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Fernández, C.; Martín-Reyes, F.; Tome, M.; Gutierrez-Repiso, C.; Fernandez-Garcia, D.; Ocaña-Wilhelmi, L.; Rivas-Becerra, J.; Tatzber, F.; Pursch, E.; Tinahones, F.J.; et al. Oxidized LDL Increase the Proinflammatory Profile of Human Visceral Adipocytes Produced by Hypoxia. Biomedicines 2021, 9, 1715. [Google Scholar] [CrossRef]
- Wang, B.; Wood, I.S.; Trayhurn, P. Dysregulation of the expression and secretion of inflammation-related adipokines by hypoxia in human adipocytes. Pflug. Arch. 2007, 455, 479–492. [Google Scholar] [CrossRef] [Green Version]
- De Glisezinski, I.; Crampes, F.; Harant, I.; Havlik, P.; Gardette, B.; Jammes, Y.; Souberbielle, J.C.; Richalet, J.P.; Rivière, D. Decrease of subcutaneous adipose tissue lipolysis after exposure to hypoxia during a simulated ascent of Mt Everest. Pflug. Arch. 1999, 439, 134–140. [Google Scholar] [CrossRef]
- Ward, M. High Altitude Medicine and Physiology, 3rd ed.; Arnold: London, UK, 2000; ISBN 0340759801. [Google Scholar]
- Bailey, D.P.; Smith, L.R.; Chrismas, B.C.; Taylor, L.; Stensel, D.J.; Deighton, K.; Douglas, J.A.; Kerr, C.J. Appetite and gut hormone responses to moderate-intensity continuous exercise versus high-intensity interval exercise, in normoxic and hypoxic conditions. Appetite 2015, 89, 237–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasse, L.K.; Sunderland, C.; King, J.A.; Batterham, R.L.; Stensel, D.J. Influence of rest and exercise at a simulated altitude of 4000 m on appetite, energy intake, and plasma concentrations of acylated ghrelin and peptide YY. J. Appl. Physiol. 2012, 112, 552–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rausch, L.K.; Netzer, N.C.; Hoegel, J.; Pramsohler, S. The Linkage between Breast Cancer, Hypoxia, and Adipose Tissue. Front. Oncol. 2017, 7, 211. [Google Scholar] [CrossRef] [Green Version]
- Matu, J.; O’Hara, J.; Hill, N.; Clarke, S.; Boos, C.; Newman, C.; Holdsworth, D.; Ispoglou, T.; Duckworth, L.; Woods, D.; et al. Changes in appetite, energy intake, body composition, and circulating ghrelin constituents during an incremental trekking ascent to high altitude. Eur. J. Appl. Physiol. 2017, 117, 1917–1928. [Google Scholar] [CrossRef]
- Netzer, N.; Gatterer, H.; Faulhaber, M.; Burtscher, M.; Pramsohler, S.; Pesta, D. Hypoxia, Oxidative Stress and Fat. Biomolecules 2015, 5, 1143–1150. [Google Scholar] [CrossRef] [Green Version]
- Debevec, T.; Simpson, E.J.; Macdonald, I.A.; Eiken, O.; Mekjavic, I.B. Exercise training during normobaric hypoxic confinement does not alter hormonal appetite regulation. PLoS ONE 2014, 9, e98874. [Google Scholar] [CrossRef]
- Dorling, J.; Broom, D.R.; Burns, S.F.; Clayton, D.J.; Deighton, K.; James, L.J.; King, J.A.; Miyashita, M.; Thackray, A.E.; Batterham, R.L.; et al. Acute and Chronic Effects of Exercise on Appetite, Energy Intake, and Appetite-Related Hormones: The Modulating Effect of Adiposity, Sex, and Habitual Physical Activity. Nutrients 2018, 10, 1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochachka, P.W.; Stanley, C.; Matheson, G.O.; McKenzie, D.C.; Allen, P.S.; Parkhouse, W.S. Metabolic and work efficiencies during exercise in Andean natives. J. Appl. Physiol. 1991, 70, 1720–1730. [Google Scholar] [CrossRef] [PubMed]
- Pramsohler, S.; Burtscher, M.; Faulhaber, M.; Gatterer, H.; Rausch, L.; Eliasson, A.; Netzer, N.C. Endurance Training in Normobaric Hypoxia Imposes Less Physical Stress for Geriatric Rehabilitation. Front. Physiol. 2017, 8, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beall, C.M. Two routes to functional adaptation: Tibetan and Andean high-altitude natives. Proc. Natl. Acad. Sci. USA 2007, 104 (Suppl. S1), 8655–8660. [Google Scholar] [CrossRef] [Green Version]
- Bigham, A.W.; Lee, F.S. Human high-altitude adaptation: Forward genetics meets the HIF pathway. Genes Dev. 2014, 28, 2189–2204. [Google Scholar] [CrossRef] [Green Version]
- Horscroft, J.A.; Kotwica, A.O.; Laner, V.; West, J.A.; Hennis, P.J.; Levett, D.Z.H.; Howard, D.J.; Fernandez, B.O.; Burgess, S.L.; Ament, Z.; et al. Metabolic basis to Sherpa altitude adaptation. Proc. Natl. Acad. Sci. USA 2017, 114, 6382–6387. [Google Scholar] [CrossRef] [Green Version]
- Barbacini, P.; Casas, J.; Torretta, E.; Capitanio, D.; Maccallini, G.; Hirschler, V.; Gelfi, C. Regulation of Serum Sphingolipids in Andean Children Born and Living at High Altitude (3775 m). Int. J. Mol. Sci. 2019, 20, 2835. [Google Scholar] [CrossRef] [Green Version]
- Voss, J.D.; Allison, D.B.; Webber, B.J.; Otto, J.L.; Clark, L.L. Lower obesity rate during residence at high altitude among a military population with frequent migration: A quasi experimental model for investigating spatial causation. PLoS ONE 2014, 9, e93493. [Google Scholar] [CrossRef] [Green Version]
- Gatterer, H.; Haacke, S.; Burtscher, M.; Faulhaber, M.; Melmer, A.; Ebenbichler, C.; Strohl, K.P.; Högel, J.; Netzer, N.C. Normobaric Intermittent Hypoxia over 8 Months Does Not Reduce Body Weight and Metabolic Risk Factors—A Randomized, Single Blind, Placebo-Controlled Study in Normobaric Hypoxia and Normobaric Sham Hypoxia. Obes. Facts 2015, 8, 200–209. [Google Scholar] [CrossRef]
- Wiesner, S.; Haufe, S.; Engeli, S.; Mutschler, H.; Haas, U.; Luft, F.C.; Jordan, J. Influences of normobaric hypoxia training on physical fitness and metabolic risk markers in overweight to obese subjects. Obesity 2010, 18, 116–120. [Google Scholar] [CrossRef]
- Debevec, T. Hypoxia-Related Hormonal Appetite Modulation in Humans during Rest and Exercise: Mini Review. Front. Physiol. 2017, 8, 366. [Google Scholar] [CrossRef] [PubMed]
- Cowburn, A.S.; Macias, D.; Summers, C.; Chilvers, E.R.; Johnson, R.S. Cardiovascular adaptation to hypoxia and the role of peripheral resistance. eLife 2017, 6, e28755. [Google Scholar] [CrossRef] [PubMed]
- Rintamäki, H. Performance and energy expenditure in cold environments. Alsk. Med. 2007, 49, 245–246. [Google Scholar]
- Matu, J.; Deighton, K.; Ispoglou, T.; Duckworth, L. The effect of moderate versus severe simulated altitude on appetite, gut hormones, energy intake and substrate oxidation in men. Appetite 2017, 113, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M. Re-Examining High-Fat Diets for Sports Performance: Did We Call the ‘Nail in the Coffin’ Too Soon? Sports Med. 2015, 45 (Suppl. S1), S33–S49. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.C.; Butterfield, G.E.; Cymerman, A.; Reeves, J.T.; Wolfel, E.E.; Brooks, G.A. Acclimatization to 4,300-m altitude decreases reliance on fat as a substrate. J. Appl. Physiol. 1996, 81, 1762–1771. [Google Scholar] [CrossRef] [Green Version]
- Young, A.J.; Berryman, C.E.; Kenefick, R.W.; Derosier, A.N.; Margolis, L.M.; Wilson, M.A.; Carrigan, C.T.; Murphy, N.E.; Carbone, J.W.; Rood, J.C.; et al. Altitude Acclimatization Alleviates the Hypoxia-Induced Suppression of Exogenous Glucose Oxidation During Steady-State Aerobic Exercise. Front. Physiol. 2018, 9, 830. [Google Scholar] [CrossRef]
- Fuhrmann, D.C.; Olesch, C.; Kurrle, N.; Schnütgen, F.; Zukunft, S.; Fleming, I.; Brüne, B. Chronic Hypoxia Enhances β-Oxidation-Dependent Electron Transport via Electron Transferring Flavoproteins. Cells 2019, 8, 172. [Google Scholar] [CrossRef] [Green Version]
- Bai, Z.; Wuren, T.; Liu, S.; Han, S.; Chen, L.; McClain, D.; Ge, R.-L. Intermittent cold exposure results in visceral adipose tissue “browning” in the plateau pika (Ochotona curzoniae). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 184, 171–178. [Google Scholar] [CrossRef]
- Hock, R.J. The physiology of high altitude. Sci. Am. 1970, 222, 52–62. [Google Scholar] [CrossRef]
- Jeremic, N.; Chaturvedi, P.; Tyagi, S.C. Browning of White Fat: Novel Insight into Factors, Mechanisms, and Therapeutics. J. Cell. Physiol. 2017, 232, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.M.; Sanchez-Gurmaches, J.; Guertin, D.A. Brown Adipose Tissue Development and Metabolism. Handb. Exp. Pharmacol. 2019, 251, 3–36. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Sebaa, R.; Malholtra, N.; Lacoste, B.; El Hankouri, Z.; Kirby, A.; Bennett, N.C.; van Jaarsveld, B.; Hart, D.W.; Tattersall, G.J.; et al. Naked mole-rat brown fat thermogenesis is diminished during hypoxia through a rapid decrease in UCP1. Nat. Commun. 2021, 12, 6801. [Google Scholar] [CrossRef] [PubMed]
- Coulson, S.Z.; Robertson, C.E.; Mahalingam, S.; McClelland, G.B. Plasticity of non-shivering thermogenesis and brown adipose tissue in high-altitude deer mice. J. Exp. Biol. 2021, 224, jeb242279. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.; Tóth, B.B.; Arianti, R.; Csomós, I.; Póliska, S.; Vámos, A.; Bacso, Z.; Győry, F.; Fésüs, L.; Kristóf, E. BMP7 Increases UCP1-Dependent and Independent Thermogenesis with a Unique Gene Expression Program in Human Neck Area Derived Adipocytes. Pharmaceuticals 2021, 14, 1078. [Google Scholar] [CrossRef]
- Tsagkaraki, E.; Nicoloro, S.M.; DeSouza, T.; Solivan-Rivera, J.; Desai, A.; Lifshitz, L.M.; Shen, Y.; Kelly, M.; Guilherme, A.; Henriques, F.; et al. CRISPR-enhanced human adipocyte browning as cell therapy for metabolic disease. Nat. Commun. 2021, 12, 6931. [Google Scholar] [CrossRef]
- Igawa, H.; Takamura, T. High-carb or low-carb, that is a question. Diabetol. Int. 2017, 8, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Brouns, F. Overweight and diabetes prevention: Is a low-carbohydrate-high-fat diet recommendable? Eur. J. Nutr. 2018, 57, 1301–1312. [Google Scholar] [CrossRef] [Green Version]
- Pitt, C.E. Cutting through the Paleo hype: The evidence for the Palaeolithic diet. Aust. Fam. Physician 2016, 45, 35–38. [Google Scholar]
- De Punder, K.; Pruimboom, L. The dietary intake of wheat and other cereal grains and their role in inflammation. Nutrients 2013, 5, 771–787. [Google Scholar] [CrossRef] [Green Version]
- Pruimboom, L.; Reheis, D. Werde Wieder Mensch: Die Rückkehr des Homo Sapiens, 1st ed.; Nova MD: Vachendorf, Germany, 2020; ISBN 9783966983075. [Google Scholar]
- National Geographic. The Evolution of Diet. Available online: https://www.nationalgeographic.com/foodfeatures/evolution-of-diet/ (accessed on 22 November 2020).
- Maixner, F.; Turaev, D.; Cazenave-Gassiot, A.; Janko, M.; Krause-Kyora, B.; Hoopmann, M.R.; Kusebauch, U.; Sartain, M.; Guerriero, G.; O’Sullivan, N.; et al. The Iceman’s Last Meal Consisted of Fat, Wild Meat, and Cereals. Curr. Biol. 2018, 28, 2348–2355.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobmann, E.D.; Byers, T.; Lanier, A.P.; Hankin, J.H.; Jackson, M.Y. The diet of Alaska Native adults: 1987–1988. Am. J. Clin. Nutr. 1992, 55, 1024–1032. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Authors | Main Outcomes |
|---|---|
| Matu et al. (2017) | Changes in plasma acylated and total ghrelin concentrations may contribute to the suppression of appetite during a high-altitude trek |
| Devebec et al. (2014) | No change in appetite regulation but improved lipid profile after exercise training in normobaric hypoxia |
| McClelland et al. (1998) | Oxygen-sparing glucose oxidation at high altitude is outweighed by limited carbohydrate stores; exercise intensity is the primary determinant for fuel selection in rats |
| Horscroft et al. (2017) | Lower capacity of fatty acid oxidation is a major metabolic adaptation in Sherpas |
| Barbacini et al. (2019) | Serum sphingolipid concentration is the primary target of hypoxia adaptation in Andean children |
| Voss et al. (2014) | There are lower obesity rates when living at higher altitudes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pramsohler, S.; Burtscher, M.; Rausch, L.; Netzer, N.C. Weight Loss and Fat Metabolism during Multi-Day High-Altitude Sojourns: A Hypothesis Based on Adipocyte Signaling. Life 2022, 12, 545. https://doi.org/10.3390/life12040545
Pramsohler S, Burtscher M, Rausch L, Netzer NC. Weight Loss and Fat Metabolism during Multi-Day High-Altitude Sojourns: A Hypothesis Based on Adipocyte Signaling. Life. 2022; 12(4):545. https://doi.org/10.3390/life12040545
Chicago/Turabian StylePramsohler, Stephan, Martin Burtscher, Linda Rausch, and Nikolaus C. Netzer. 2022. "Weight Loss and Fat Metabolism during Multi-Day High-Altitude Sojourns: A Hypothesis Based on Adipocyte Signaling" Life 12, no. 4: 545. https://doi.org/10.3390/life12040545
APA StylePramsohler, S., Burtscher, M., Rausch, L., & Netzer, N. C. (2022). Weight Loss and Fat Metabolism during Multi-Day High-Altitude Sojourns: A Hypothesis Based on Adipocyte Signaling. Life, 12(4), 545. https://doi.org/10.3390/life12040545