
Evidence for a Strong Relationship between the Cytotoxicity and Intracellular Location of β-Amyloid


 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Purification and Preparation of Aβ42 and tAβ42 Peptides
2.3. Cell Culture and Cytotoxicity Assay
2.4. Immunocytochemistry
2.5. Measurement of Caspase Activity
2.6. Western Blot Analysis
2.7. Fibrillogenesis
2.8. Analysis of Secondary Structure
2.9. Transmission Electron Microscopy (TEM)
3. Results
3.1. Cytotoxicity of Wild-Type Aβ42 and tAβ42
3.2. Internalization of Aβ42 into the Cells
3.3. Effect of the Inhibition of Aβ42 Cytotoxicity on Cell Viability and the Internalization of Aβ42 into Cells
3.4. Lamin Protein Fragmentation and Caspase Activation Induced by tAβ42
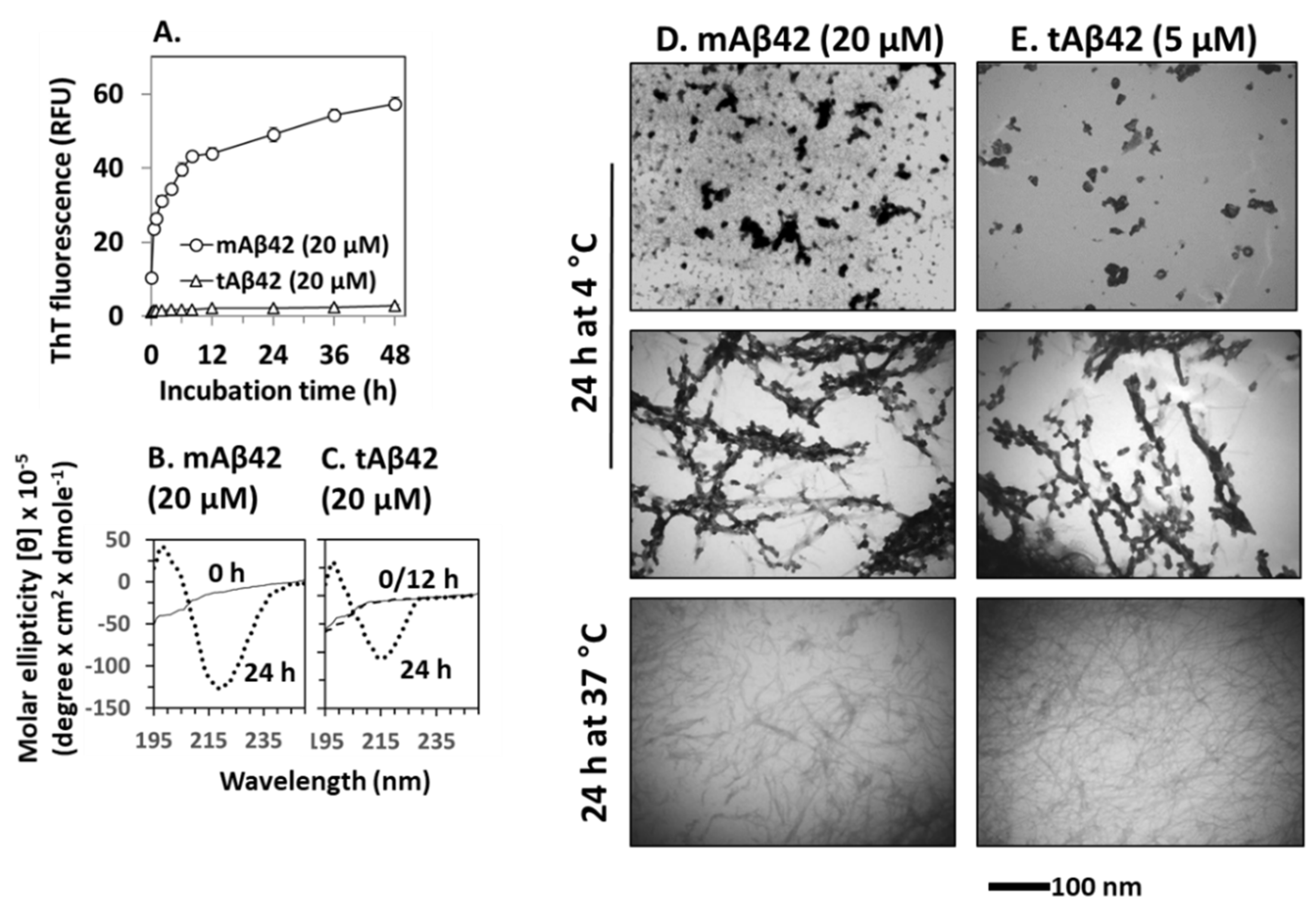
3.5. Structural Characterization of tAB42
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, X.; Chen, W.D.; Wang, Y.D. beta-Amyloid: The key peptide in the pathogenesis of Alzheimer’s disease. Front. Pharmacol. 2015, 6, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.F.; Xu, T.H.; Yan, Y.; Zhou, Y.R.; Jiang, Y.; Melcher, K.; Xu, H.E. Amyloid beta: Structure, biology and structure-based therapeutic development. Acta Pharmacol. Sin. 2017, 38, 1205–1235. [Google Scholar] [CrossRef] [PubMed]
- Terry, R.D.; Gonatas, N.K.; Weiss, M. Ultrastructural Studies in Alzheimer’s Presenile Dementia. Am. J. Pathol. 1964, 44, 269–297. [Google Scholar] [PubMed]
- Goate, A.; Chartier-Harlin, M.C.; Mullan, M.; Brown, J.; Crawford, F.; Fidani, L.; Giuffra, L.; Haynes, A.; Irving, N.; James, L.; et al. Segregation of a missense mutation in the amyloid precursor protein gene with familial Alzheimer’s disease. Nature 1991, 349, 704–706. [Google Scholar] [CrossRef] [PubMed]
- Suh, Y.H. An etiological role of amyloidogenic carboxyl-terminal fragments of the beta-amyloid precursor protein in Alzheimer’s disease. J. Neurochem. 1997, 68, 1781–1791. [Google Scholar] [CrossRef]
- McLean, C.A.; Cherny, R.A.; Fraser, F.W.; Fuller, S.J.; Smith, M.J.; Beyreuther, K.; Bush, A.I.; Masters, C.L. Soluble pool of Abeta amyloid as a determinant of severity of neurodegeneration in Alzheimer’s disease. Ann. Neurol. 1999, 46, 860–866. [Google Scholar] [CrossRef]
- Haass, C.; Selkoe, D.J. Soluble protein oligomers in neurodegeneration: Lessons from the Alzheimer’s amyloid beta-peptide. Nat. Rev. Mol. Cell Biol. 2007, 8, 101–112. [Google Scholar] [CrossRef]
- Cleary, J.P.; Walsh, D.M.; Hofmeister, J.J.; Shankar, G.M.; Kuskowski, M.A.; Selkoe, D.J.; Ashe, K.H. Natural oligomers of the amyloid-beta protein specifically disrupt cognitive function. Nat. Neurosci. 2005, 8, 79–84. [Google Scholar] [CrossRef]
- Shahnawaz, M.; Bilkis, T.; Park, I.S. Amyloid β cytotoxicity is enhanced or reduced depending on formation of amyloid β oligomeric forms. Biotechnol. Lett. 2021, 43, 165–175. [Google Scholar] [CrossRef]
- De, S.; Wirthensohn, D.C.; Flagmeier, P.; Hughes, C.; Aprile, F.A.; Ruggeri, F.S.; Whiten, D.R.; Emin, D.; Xia, Z.; Varela, J.A.; et al. Different soluble aggregates of Abeta42 can give rise to cellular toxicity through different mechanisms. Nat. Commun. 2019, 10, 1541. [Google Scholar] [CrossRef] [Green Version]
- Wells, C.; Brennan, S.; Keon, M.; Ooi, L. The role of amyloid oligomers in neurodegenerative pathologies. Int. J. Biol. Macromol. 2021, 181, 582–604. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Vidal, G.S.; Djurisic, M.; William, C.M.; Birnbaum, M.E.; Garcia, K.C.; Hyman, B.T.; Shatz, C.J. Human LilrB2 is a beta-amyloid receptor and its murine homolog PirB regulates synaptic plasticity in an Alzheimer’s model. Science 2013, 341, 1399–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, L.; Harris, D.A. Abeta receptors specifically recognize molecular features displayed by fibril ends and neurotoxic oligomers. Nat. Commun. 2021, 12, 3451. [Google Scholar] [CrossRef] [PubMed]
- Kienlen-Campard, P.; Miolet, S.; Tasiaux, B.; Octave, J.N. Intracellular amyloid-beta 1-42, but not extracellular soluble amyloid-beta peptides, induces neuronal apoptosis. J. Biol. Chem. 2002, 277, 15666–15670. [Google Scholar] [CrossRef] [Green Version]
- Wong, P.T.; Schauerte, J.A.; Wisser, K.C.; Ding, H.; Lee, E.L.; Steel, D.G.; Gafni, A. Amyloid-beta membrane binding and permeabilization are distinct processes influenced separately by membrane charge and fluidity. J. Mol. Biol. 2009, 386, 81–96. [Google Scholar] [CrossRef]
- Chafekar, S.M.; Baas, F.; Scheper, W. Oligomer-specific Abeta toxicity in cell models is mediated by selective uptake. Biochim. Biophys. Acta 2008, 1782, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Thapa, A.; Woo, E.R.; Chi, E.Y.; Sharoar, M.G.; Jin, H.G.; Shin, S.Y.; Park, I.S. Biflavonoids are superior to monoflavonoids in inhibiting amyloid-beta toxicity and fibrillogenesis via accumulation of nontoxic oligomer-like structures. Biochemistry 2011, 50, 2445–2455. [Google Scholar] [CrossRef]
- Sharoar, M.G.; Shahnawaz, M.; Islam, M.I.; Ramasamy, V.S.; Shin, S.Y.; Park, I.S. The inhibitory effects of Escherichia coli maltose binding protein on beta-amyloid aggregation and cytotoxicity. Arch. Biochem. Biophys. 2013, 538, 41–48. [Google Scholar] [CrossRef]
- Thapa, A.; Shahnawaz, M.; Karki, P.; Raj Dahal, G.; Sharoar, M.G.; Yub Shin, S.; Sup Lee, J.; Cho, B.; Park, I.S. Purification of inclusion body-forming peptides and proteins in soluble form by fusion to Escherichia coli thermostable proteins. Biotechniques 2008, 44, 787–796. [Google Scholar] [CrossRef]
- Münst, B.; Patsch, C.; Edenhofer, F. Engineering cell-permeable protein. J. Vis. Exp. 2009, 2009, 1627. [Google Scholar] [CrossRef] [Green Version]
- Vives, E.; Brodin, P.; Lebleu, B. A truncated HIV-1 Tat protein basic domain rapidly translocates through the plasma membrane and accumulates in the cell nucleus. J. Biol. Chem. 1997, 272, 16010–16017. [Google Scholar] [CrossRef] [Green Version]
- Shahnawaz, M.; Thapa, A.; Park, I.S. Stable activity of a deubiquitylating enzyme (Usp2-cc) in the presence of high concentrations of urea and its application to purify aggregation-prone peptides. Biochem. Biophys. Res. Commun. 2007, 359, 801–805. [Google Scholar] [CrossRef] [PubMed]
- Catanzariti, A.M.; Soboleva, T.A.; Jans, D.A.; Board, P.G.; Baker, R.T. An efficient system for high-level expression and easy purification of authentic recombinant proteins. Protein Sci. 2004, 13, 1331–1339. [Google Scholar] [CrossRef]
- Shahnawaz, M.; Sharoar, M.G.; Shin, S.Y.; Park, I.S. Wild-type, Flemish, and Dutch amyloid-beta exhibit different cytotoxicities depending on Abeta40 to Abeta42 interaction time and concentration ratio. J. Pept. Sci. 2013, 19, 545–553. [Google Scholar] [CrossRef]
- Karki, P.; Dahal, G.R.; Park, I.S. Both dimerization and interdomain processing are essential for caspase-4 activation. Biochem. Biophys. Res. Commun. 2007, 356, 1056–1061. [Google Scholar] [CrossRef] [PubMed]
- Sharoar, M.G.; Islam, M.I.; Shahnawaz, M.; Shin, S.Y.; Park, I.S. Amyloid beta binds procaspase-9 to inhibit assembly of Apaf-1 apoptosome and intrinsic apoptosis pathway. Biochim. Biophys. Acta 2014, 1843, 685–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slaughter, M.R.; Bugelski, P.J.; O’Brien, P.J. Evaluation of alamar blue reduction for the in vitro assay of hepatocyte toxicity. Toxicol. Vitr. 1999, 13, 567–569. [Google Scholar] [CrossRef]
- Wogulis, M.; Wright, S.; Cunningham, D.; Chilcote, T.; Powell, K.; Rydel, R.E. Nucleation-dependent polymerization is an essential component of amyloid-mediated neuronal cell death. J. Neurosci. 2005, 25, 1071–1080. [Google Scholar] [CrossRef] [Green Version]
- Ramasamy, V.S.; Islam, M.I.; Haque, M.A.; Shin, S.Y.; Park, I.S. beta-Amyloid induces nuclear protease-mediated lamin fragmentation independent of caspase activation. Biochim. Biophys. Acta 2016, 1863, 1189–1199. [Google Scholar] [CrossRef]
- Islam, M.I.; Hossain, M.S.; Park, I.S. Differential involvement of caspase-6 in amyloid-β-induced fragmentation of lamin A and B. Biochem. Biophys. Rep. 2020, 24, 100839. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Lim, G.P.; Begum, A.N.; Ubeda, O.J.; Simmons, M.R.; Ambegaokar, S.S.; Chen, P.P.; Kayed, R.; Glabe, C.G.; Frautschy, S.A.; et al. Curcumin inhibits formation of amyloid beta oligomers and fibrils, binds plaques, and reduces amyloid in vivo. J. Biol. Chem. 2005, 280, 5892–5901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano, F.X.; Galbete, J.L.; Forloni, G. Effect of beta-amyloid on endothelial cells: Lack of direct toxicity, enhancement of MTT-induced cell death and intracellular accumulation. Neurochem. Int. 2003, 43, 251–261. [Google Scholar] [CrossRef]
- Ono, K.; Condron, M.M.; Teplow, D.B. Structure-neurotoxicity relationships of amyloid beta-protein oligomers. Proc. Natl. Acad. Sci. USA 2009, 106, 14745–14750. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.I.; Sharoar, M.G.; Ryu, E.K.; Park, I.S. Limited activation of the intrinsic apoptotic pathway plays a main role in amyloid-beta-induced apoptosis without eliciting the activation of the extrinsic apoptotic pathway. Int. J. Mol. Med. 2017, 40, 1971–1982. [Google Scholar] [CrossRef] [Green Version]
- Tjernberg, L.O.; Pramanik, A.; Bjorling, S.; Thyberg, P.; Thyberg, J.; Nordstedt, C.; Berndt, K.D.; Terenius, L.; Rigler, R. Amyloid beta-peptide polymerization studied using fluorescence correlation spectroscopy. Chem. Biol. 1999, 6, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Pike, C.J.; Walencewicz, A.J.; Glabe, C.G.; Cotman, C.W. In vitro aging of beta-amyloid protein causes peptide aggregation and neurotoxicity. Brain Res. 1991, 563, 311–314. [Google Scholar] [CrossRef]
- Limbocker, R.; Chia, S.; Ruggeri, F.S.; Perni, M.; Cascella, R.; Heller, G.T.; Meisl, G.; Mannini, B.; Habchi, J.; Michaels, T.C.T.; et al. Trodusquemine enhances Abeta42 aggregation but suppresses its toxicity by displacing oligomers from cell membranes. Nat. Commun. 2019, 10, 225. [Google Scholar] [CrossRef] [Green Version]
- Thornberry, N.A.; Lazebnik, Y. Caspases: Enemies within. Science 1998, 281, 1312–1316. [Google Scholar] [CrossRef]
- Behl, C. Apoptosis and Alzheimer’s disease. J. Neural. Transm. 2000, 107, 1325–1344. [Google Scholar] [CrossRef]
- Shimohama, S. Apoptosis in Alzheimer’s disease—An update. Apoptosis 2000, 5, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Naiki, H.; Hasegawa, K.; Yamaguchi, I.; Nakamura, H.; Gejyo, F.; Nakakuki, K. Apolipoprotein E and antioxidants have different mechanisms of inhibiting Alzheimer’s beta-amyloid fibril formation in vitro. Biochemistry 1998, 37, 17882–17889. [Google Scholar] [CrossRef]
- Jan, A.; Gokce, O.; Luthi-Carter, R.; Lashuel, H.A. The ratio of monomeric to aggregated forms of Abeta40 and Abeta42 is an important determinant of amyloid-beta aggregation, fibrillogenesis, and toxicity. J. Biol. Chem. 2008, 283, 28176–28189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.; Davis, J.; Aucoin, D.; Sato, T.; Ahuja, S.; Aimoto, S.; Elliott, J.I.; Van Nostrand, W.E.; Smith, S.O. Structural conversion of neurotoxic amyloid-beta(1-42) oligomers to fibrils. Nat. Struct. Mol. Biol. 2010, 17, 561–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, L.; Liu, C.; Guo, Z. Structural insights into Abeta42 oligomers using site-directed spin labeling. J. Biol. Chem. 2013, 288, 18673–18683. [Google Scholar] [CrossRef] [Green Version]
- Viola, K.L.; Klein, W.L. Amyloid beta oligomers in Alzheimer’s disease pathogenesis, treatment, and diagnosis. Acta Neuropathol. 2015, 129, 183–206. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haque, M.A.; Hossain, M.S.; Bilkis, T.; Islam, M.I.; Park, I.-S. Evidence for a Strong Relationship between the Cytotoxicity and Intracellular Location of β-Amyloid. Life 2022, 12, 577. https://doi.org/10.3390/life12040577
Haque MA, Hossain MS, Bilkis T, Islam MI, Park I-S. Evidence for a Strong Relationship between the Cytotoxicity and Intracellular Location of β-Amyloid. Life. 2022; 12(4):577. https://doi.org/10.3390/life12040577
Chicago/Turabian StyleHaque, Md. Aminul, Md. Selim Hossain, Tahmina Bilkis, Md. Imamul Islam, and Il-Seon Park. 2022. "Evidence for a Strong Relationship between the Cytotoxicity and Intracellular Location of β-Amyloid" Life 12, no. 4: 577. https://doi.org/10.3390/life12040577
APA StyleHaque, M. A., Hossain, M. S., Bilkis, T., Islam, M. I., & Park, I. -S. (2022). Evidence for a Strong Relationship between the Cytotoxicity and Intracellular Location of β-Amyloid. Life, 12(4), 577. https://doi.org/10.3390/life12040577