
Identification and Functional Analysis of SabHLHs in Santalum album L.


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Screening and Cloning of bHLH Transcription Factors
2.3. Bioinformatics Analysis of SabHLHs
2.4. Subcellular Localization Analysis
2.5. Interaction Detection of Transcription Factors SaMYC1, SabHLH1 and G-Box Elements
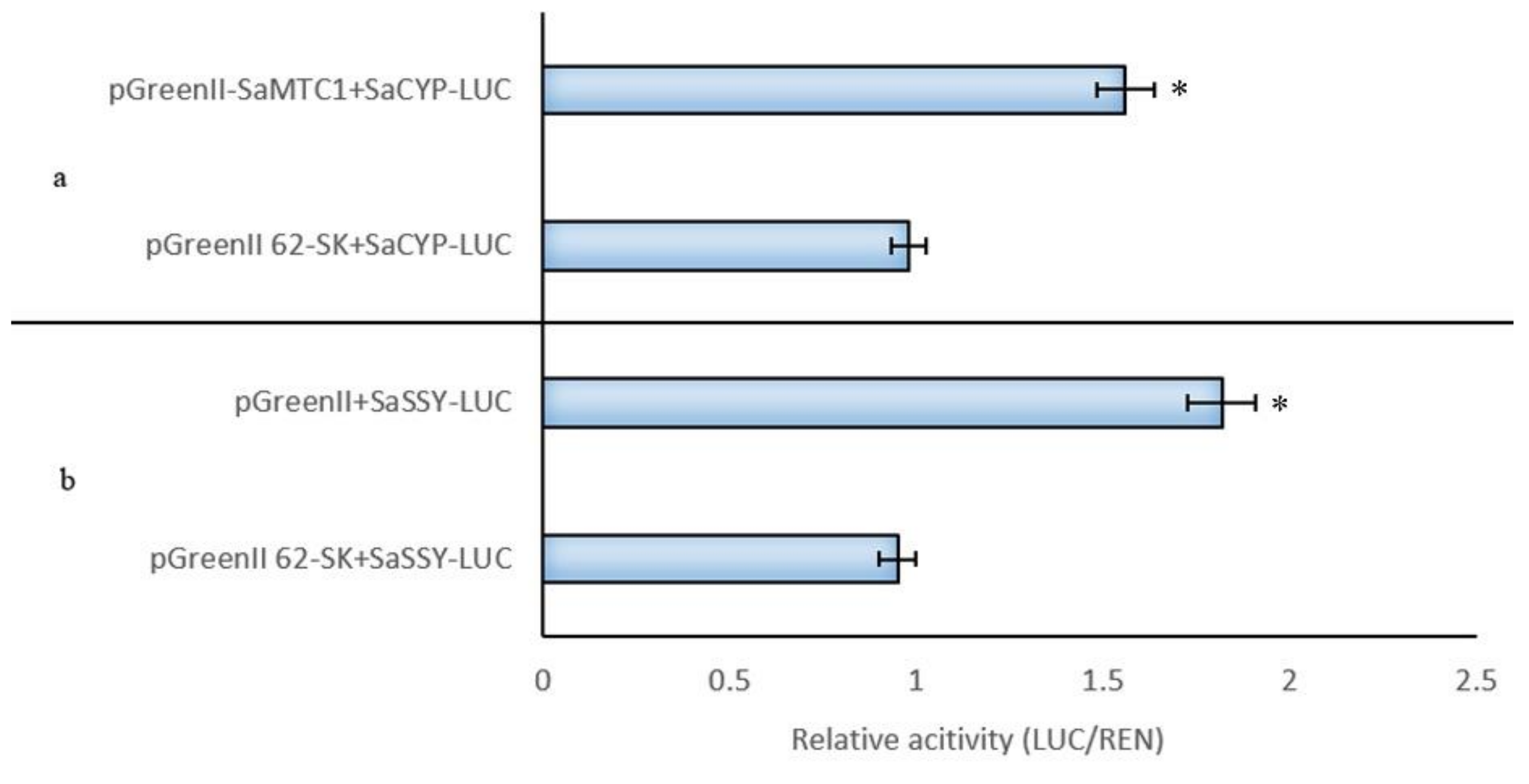
2.6. SaMYC1 Activated SaSSy and SaCYP736A167 Promoter Activity
2.7. Statistical Analysis
3. Results
3.1. Cloning SabHLHs ORFs
3.2. Analysis of the Physicochemical Properties of SabHLH Proteins
3.3. Conservative Motif Analysis of SabHLH Proteins
3.4. Phylogenetic Analysis of SabHLH Proteins
3.5. Subcellular Localization Analysis
3.6. SaMYC1 Conservative Domain Prediction
3.7. Analysis of G-Box Elements and E-Box Elements in Promoters SaSSy and SaCYP736A167
3.8. Verification of the Activity of Transcription Factors
3.9. Screening AbA Concentrations That Inhibiting the Growth of Bait Strains
3.10. Assessment of the AbA Concentration That Inhibits the Growth of G-Box Mutant Element Bait Strains
3.11. Detection of the Interaction between SaMYC1 and Mutant G-Box Elements
3.12. SaMYC1 Activated SaSSy and SaCYP736A167 Promoter Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jin, J.; Zhang, H.; Kong, L.; Gao, G.; Luo, J. PlantTFDB 3.0: A portal for the functional and evolutionary study of plant transcription factors. Nucl. Acid. Res. 2014, 42, 1182–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, Y.C.; Hung, Y.C.; Tsai, W.C.; Chen, W.H.; Chen, H.H. PbbHLH4 regulates floral monoterpene biosynthesis in Phalaenopsis orchids. J. Exp. Bot. 2018, 69, 4363–4377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.F.; Li, M.Z.; Xiong, Y.P.; Wu, J.M.; Teixeira da Silva, J.A.; Ma, G.H. Genome-wide characterization, expression profile analysis of WRKY family genes in Santalum album and functional identification of their role in abiotic stress. Int. J. Mol. Sci. 2019, 20, 5676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieuwenhuizen, N.J.; Chen, X.; Wang, M.Y.; Matich, A.J.; Perez, R.L.; Allan, A.C.; Green, S.A.; Atkinson, R.G. Natural variation in monoterpene synthesis in kiwifruit: Transcriptional regulation of terpene synthases by nac and ethylene-insensitive3-like transcription factors. Plant Physiol. 2015, 167, 1243–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, S. An overview of the basic helix-loop-helix proteins. Genom. Biol. 2004, 5, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carretero-Paulet, L.; Galstyan, A.; Roig-Villanova, I.; Carretero-Paulet, L.; Galstyan, A.; Roig-Villanova, I.; Martínez-García, J.F.; Bilbao-Castro, J.R.; Robertson, D.L. Genome-wide classification and evolutionary analysis of the bHLH family of transcription factors in Arabidopsis, poplar, rice, moss, and algae. Plant Physiol. 2010, 153, 1398–1412. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.Y.; Hu, Z.Z.; Zhao, T.M.; Yang, Y.W.; Chen, T.Z.; Yang, M.L.; Yu, W.G.; Zhang, B.L. Genome-wide analysis of bHLH transcription factor and involvement in the infection by yellow leaf curl virus in tomato (Solanum lycopersicum). BMC Genom. 2015, 16, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathalie, N.; Debeaujon, I.; Jond, C.; Pelletier, G.; Caboche, M.; Lepiniec, L. The TT8 gene encodes a basic helix-loop-helix domain protein required for expression of DFR and BAN genes in Arabidopsis siliques. Plant Cell 2014, 12, 1863–1878. [Google Scholar]
- Zhu, Z.; Liang, H.; Chen, G.; Li, F.; Hu, Z. The bHLH transcription factor SlPRE2 regulates tomato fruit development and modulates plant response to gibberellin. Plant Cell Rep. 2019, 38, 1053–1064. [Google Scholar] [CrossRef]
- Qi, T.-C.; Huang, H.; Song, S.-S.; Xie, D. Regulation of jasmonate-mediated stamen development and seed production by a bHLH-MYB complex in Arabidopsis. Plant Cell 2015, 27, 1620–1633. [Google Scholar] [CrossRef] [Green Version]
- Pillitteri, L.J.; Torii, K.U. Breaking the silence: Three bHLH proteins direct cell-fate decisions during stomatal development. Bioessays 2007, 29, 861–870. [Google Scholar] [CrossRef]
- Chater, C.C.C.; Caine, R.S.; Fleming, A.J.; Gray, J.E. Origins and evolution of stomatal development. Plant Physiol. 2007, 174, 624–638. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, G.; Theres, K. Shoot and inflorescence branching. Curr. Opin. Plant Biol. 2005, 8, 506–511. [Google Scholar] [CrossRef] [Green Version]
- Duek, P.D.; Fankhauser, C. bHLH class transcription factors take centre stage in phytochrome signalling. Trends Plant Sci. 2005, 10, 51–54. [Google Scholar] [CrossRef]
- Friedrichsen, D.M.; Nemhauser, J.; Muramitsu, T.; Maloof, J.N.; Alonso, J.; Ecker, J.R.; Furuya, M.; Chory, J. Three redundant brassinosteroid early response genes encode putative bHLH transcription factors required for normal growth. Genetics 2002, 162, 1445–1456. [Google Scholar] [CrossRef]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Chini, A.; Fonseca, S.; Fernández, G.; Adie, B.; Chico, J.M.; Lorenzo, O.; García-Casado, G.; López-Vidriero, I.; Lozano, F.M.; Ponce, M.R.; et al. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 2007, 448, 666–671. [Google Scholar] [CrossRef]
- Hu, D.G.; Yu, J.Q.; Han, P.L.; Hu, D.-G.; Yu, J.Q.; Han, P.L.; Xie, X.B.; Sun, C.H.; Zhang, Q.Y.; Wang, J.H.; et al. The regulatory module MdPUB29-MdbHLH3 connects ethylene biosynthesis with fruit quality in apple. N. Phytol. 2019, 221, 1966–1982. [Google Scholar] [CrossRef]
- Choi, H.; Oh, E. PIF4 Integrates multiple environmental and hormonal signals for plant growth regulation in Arabidopsis. Mol. Cells 2016, 39, 587–593. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Moon, S.; Jang, S.; Lee, J.; Moon, S.; Jang, S.; Lee, S.; An, G.; Jung, K.H.; Park, S.K. OsbHLH073 negatively regulates internode elongation and plant height by modulating GA homeostasis in rice. Plants 2020, 9, 547. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Hwang, G.; Kim, S.; Thi, T.N.; Oh, E. The epidermis coordinates thermoresponsive growth through the phyB-PIF4-auxin pathway. Nature Comm. 2020, 11, 1053. [Google Scholar] [CrossRef] [Green Version]
- Dozmorov, M.G.; Yang, Q.; Wu, W.J.; Wren, J.; Suhail, M.M.; Woolley, C.L.; Young, D.G.; Fung, K.M.; Lin, H.K. Differential effects of selective frankincense (Ru Xiang) essential oil versus non-selective sandalwood (Tan Xiang) essential oil on cultured bladder cancer cells: A microarray and bioinformatics study. Chin. Med. 2014, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Santha, S.; Dwivedi, C. Anticancer effects of sandalwood (Santalum album). Anticanc. Res. 2015, 35, 3137–3145. [Google Scholar]
- Scartezzini, P.; Speroni, E. Review on some plants of Indian traditional medicine with antioxidant activity. J. Ethnoph. 2000, 71, 23–43. [Google Scholar] [CrossRef]
- Soneja, A.; Kaushik, P.; Kaushik, D.; Kumar, S.; Kumar, D. Antioxidant, Analgesic and Anti-inflammatory Activities of Santalum album Linn. Plant. Med. 2009, 75, 452–453. [Google Scholar] [CrossRef]
- Suganya, K.; Liu, Q.F.; Koo, B.S. Santalum album extract exhibits neuroprotective effect against the TLR3-mediated neuroinflammatory response in human SH-SY5Y neuroblastoma cells. Phytother. Res. 2021, 35, 1991–2004. [Google Scholar] [CrossRef]
- Han, X.; Beaumont, C.; Stevens, N. Chemical composition analysis and in vitro biological activities of ten essential oils in human skin cells. Biochim. Open 2017, 5, 1–7. [Google Scholar] [CrossRef]
- Moy, R.L.; Levenson, C. Sandalwood album oil as a botanical therapeutic in dermatology. J. Clin. Aesth. Derm. 2017, 10, 34–39. [Google Scholar]
- Baldovini, N.; Delasalle, C.; Joulain, D. Phytochemistry of the heartwood from fragrant Santalum species: A review. Flav. Frag. J. 2011, 26, 7–26. [Google Scholar] [CrossRef]
- Jones, C.G.; Moniodis, J.; Zulak, K.G.; Scaffidi, A.; Plummer, J.A.; Ghisalberti, E.L.; Barbour, E.L.; Bohlmann, J. Sandalwood fragrance biosynthesis involves sesquiterpene synthases of both the terpene synthase (TPS)-a and TPS-b subfamilies, including santalene synthases. J. Biol. Chem. 2011, 286, 17445–17454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Chavez, M.L.; Moniodis, J.; Madilao, L.L.; Jancsik, S.; Keeling, C.I.; Barbour, E.L.; Ghisalberti, E.L.; Plummer, J.A.; Jones, C.G.; Bohlmann, J. Biosynthesis of sandalwood oil: Santalum album CYP76F Cytochromes P450 produce santalols and bergamotol. PLoS ONE 2013, 8, e75053. [Google Scholar] [CrossRef] [Green Version]
- Celedon, J.M.; Bohlmann, J. Genomics-based discovery of plant genes for synthetic biology of terpenoid fragrances: A case study in sandalwood oil biosynthesis. In Synthetic Biology and Metabolic Engineering in Plants and Microbes; Academic Press: Cambridge, MA, USA, 2016; Volume 576, pp. 47–67. [Google Scholar]
- Srivastava, P.L.; Daramwar, P.P.; Krithika, R.; Pandreka, A.; Shankar, S.S.; Thulasiram, H.V. Functional characterization of novel sesquiterpene synthases from Indian sandalwood, Santalum album. Sci. Rep. 2015, 5, 10095. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.H.; Niu, M.Y.; Teixeira da Silva, J.A.; Zhang, Y.Y.; Yuan, Y.F.; Jia, Y.X.; Xiao, Y.Y.; Li, Y.; Fang, L.; Zeng, S.J.; et al. Identification and functional characterization of three new terpene synthase genes involved in chemical defense and abiotic stresses in Santalum album. BMC Plant Biol. 2019, 19, 115. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.H.; Teixeira da Silva, J.A.; Niu, M.Y.; Zhang, T.; Liu, H.F.; Zheng, F.; Yuan, Y.F.; Li, Y.; Fang, L.; Zeng, S.J.; et al. Functional characterization of an Indian sandalwood (Santalum album L.) dual-localized bifunctional nerolidol/linalool synthase gene involved in stress response. Phytochemistry 2021, 183, 112610. [Google Scholar] [CrossRef]
- Niu, M.Y.; Yan, H.F.; Xiong, Y.P.; Zhang, Y.Y.; Zhang, X.H.; Li, Y.; Teixeira da Silva, J.A.; Ma, G.H. Cloning, characterization, and functional analysis of acetyl-CoA C-acetyltransferase and 3-hydroxy-3-methylglutaryl-CoA synthase genes in Santalum album. Sci. Rep. 2021, 11, 1082. [Google Scholar] [CrossRef]
- Niu, M.Y.; Xiong, Y.P.; Yan, H.F.; Zhang, X.H.; Li, Y.; Teixeira da Silva, J.A.; Ma, G.H. Cloning and expression analysis of mevalonate kinase and phosphomevalonate kinase genes associated with the MVA pathway in Santalum album. Sci. Rep. 2021, 11, 16913. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Yan, H.F.; Li, Y.; Xiong, Y.P.; Niu, M.Y.; Zhang, X.H.; Teixeira da Silva, J.A.; Ma, G.H. Molecular cloning and functional identification of 1-deoxy-D-xylulose 1,5-phosphate reductoisomerase from Santalum album. Genes 2021, 12, 626. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, T.; Zhang, H.; Fu, S.L.; Zhao, G.Y.; Jia, J.Z.; Kong, X.Y. LRRC31 inhibits DNA repair and sensitizes breast cancer brain metastasis to radiation therapy. Nat. Cell Biol. 2020, 22, 1276–1285. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, C.; Han, X.; Han, X.; Tang, S.; Liu, S.; Xia, X.L.; Yin, W.L. A novel bHLH transcription factor PebHLH35 from Populus euphratica confers drought tolerance through regulating stomatal development, photosynthesis and growth in Arabidopsis. Biochem. Biophys. Res. Comm. 2014, 450, 453–458. [Google Scholar] [CrossRef]
- Yao, P.F.; Sun, Z.X.; Li, C.L.; Li, M.; Deng, R.; Huang, Y.; Zhao, H.; Chen, H.; Wu, Q. Overexpression of Fagopyrum tataricum FtbHLH2 enhances tolerance to cold stress in transgenic Arabidopsis. Plant Physiol. Biochem. 2018, 125, 85–94. [Google Scholar] [CrossRef]
- Zhai, Y.; Zhang, L.; Xia, C.; Chuan, S.L.; Zhao, G.Y.; Kong, X.Y. The wheat transcription factor, TabHLH39, improves tolerance to multiple abiotic stressors in transgenic plants. Biochem. Bioph. Res. Comm. 2016, 473, 1321–1327. [Google Scholar] [CrossRef]
- Ludwig, S.R.; Habera, L.F.; Dellaporta, S.L.; Wessler, S.R. Lc, a member of the maize R gene family responsible for tissue-specific anthocyanin production, encodes a protein similar to transcriptional activators and contains the myc-homology region. Proc. Nat. Acad. Sci. USA 1989, 86, 7092–7096. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef]
- Schweizer, F.; Fernández-Calvo, P.; Zander, M.; Diez-Diaz, M.; Fonseca, S.; Glauser, G.; Lewsey, M.G.; Ecker, J.R.; Solano, R.; Reymond, P. Arabidopsis basic helix-loop-helix transcription factors MYC2, MYC3, and MYC4 regulate glucosinolate biosynthesis, insect performance, and feeding behavior. Plant Cell 2013, 25, 3117–3132. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.T.; Hedhili, S.; Montiel, G.; Zhang, Y.X.; Guillaume, C.; Martial, P.; Pascal, G.; Memelink, J. The basic helix-loop-helix transcription factor CrMYC2 controls the jasmonate-responsive expression of the ORCA genes that regulate alkaloid biosynthesis in Catharanthus roseus. Plant J. 2011, 67, 61–71. [Google Scholar] [CrossRef]
- Ji, Y.; Xiao, J.; Shen, Y.; Ma, D.M.; Li, Z.Q.; Pu, G.B.; Li, X.; Huang, L.L.; Liu, B.Y.; Ye, H.C.; et al. Cloning and characterization of AabHLH1, a bHLH transcription factor that positively regulates artemisinin biosynthesis in Artemisia annua. Plant Cell Physiol. 2014, 55, 1592–1604. [Google Scholar] [CrossRef] [Green Version]
- Hong, G.J.; Xue, X.Y.; Mao, Y.B.; Wang, L.J.; Chen, X.-Y. Arabidopsis MYC2 interacts with DELLA proteins in regulating sesquiterpene synthase gene expression. Plant Cell 2012, 24, 2635–2648. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.S.; van Herwijnen, Z.O.; Dräger, D.B.; Dräger, D.B.; Sui, C.; Haring, M.A.; Schuurink, R.C. SlMYC1 regulates type VI glandular trichome formation and terpene biosynthesis in tomato glandular cells. Plant Cell 2018, 30, 2988–3005. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.H.; Liao, Y.C.; Lv, F.F.; Zhang, Z.; Sun, P.W.; Gao, Z.H.; Hu, K.P.; Sui, C.; Jin, Y.; Wei, J.H. Transcription factor AsMYC2 controls the jasmonate-responsive expression of ASS1 regulating sesquiterpene biosynthesis in Aquilaria sinensis (Lour.) Gilg. Plant Cell Physiol. 2017, 58, 1924–1933. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Li, Y.; Gao, F.; Jin, W.; Li, S.; Shadrack, K.; Yang, S.; Bao, T.; Gao, X.; Wang, L. MYB21 interacts with MYC2 to control the expression of terpene synthase genes in flowers of Freesia hybrida and Arabidopsis thaliana. J. Exp. Bot. 2020, 71, 4140–4158. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.Z.; Lin, X.; Li, X.; Yang, N.; Chen, L. Molecular cloning and functional characterization of CpMYC2 and CpBHLH13 transcription factors from wintersweet (Chimonanthus praecox L.). Plants 2020, 9, 785. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.Q.; Ahmad, N.; Tian, Y.Y.; Liu, J.; Li, H. Genome-wide identification, expression analysis, and subcellular localization of Carthamus tinctorius bHLH transcription factors. Int. J. Mol. Sci. 2019, 20, 3044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.H.; Wang, J.W.; Wang, S.; Wang, J.Y.; Chen, X.Y. Characterization of GaWRKY1, a cotton transcription factor that regulates the sesquiterpene synthase gene (+)-δ-cadinene synthase-A. Plant Physiol. 2004, 135, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Wang, H.; Lundgren, A.; Brodelius, P.E. Effects of overexpression of AaWRKY1 on artemisinin biosynthesis in transgenic Artemisia annua plants. Phytochemistry 2014, 102, 89–96. [Google Scholar] [CrossRef]
- Tang, K.X.; Pan, Q.F.; Zhang, F.Y.; Fu, X.; Lv, Z.; Zhang, F.; Pan, Q.; Wang, G.; Sun, X.; Tang, K. The jasmonate-responsive AaMYC2 transcription factor positively regulates artemisinin biosynthesis in Artemisia annua. N. Phytol. 2016, 210, 1269–1281. [Google Scholar]
- Wang, Q.; Liu, H.; Zhang, M.; Liu, S.H.; Zhang, Y.H. MdMYC2 and MdERF3 positively co-regulate α-Farnesene biosynthesis in Apple. Front. Plant Sci. 2020, 11, 512844. [Google Scholar] [CrossRef]
- Feyissa, D.N.; Løvdal, T.; Olsen, K.M.; Slimestad, R.; Lillo, C. The endogenous GL3, but not EGL3, gene is necessary for anthocyanin accumulation as induced by nitrogen depletion in Arabidopsis rosette stage leaves. Planta 2009, 230, 747–754. [Google Scholar] [CrossRef]
- Xie, Y.; Tan, H.; Ma, Z.; Huang, J.R. DELLA proteins promote anthocyanin biosynthesis via sequestering MYBL2 and JAZ suppressors of the MYB/bHLH/WD40 complex in Arabidopsis thaliana. Mol. Plant 2016, 9, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.J.; Dubos, C.; Lepiniec, L. Transcriptional control of flavonoid biosynthesis by MYB-bHLH-WDR complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef]
- Baudry, A.; Heim, M.A.; Dubreucq, B.; Caboche, M.; Weisshaar, B.; Lepiniec, L. TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana. Plant J. 2004, 39, 366–380. [Google Scholar] [CrossRef]
- Hichri, I.; Heppel, S.C.; Pillet, J.; Léon, C.; Czemmel, S.; Delrot, S.; Lauvergeat, V.; Bogs, J. The basic helix-loop-helix transcription factor MYC1 is involved in the regulation of the flavonoid biosynthesis pathway in grapevine. Mol. Plant 2010, 3, 509–523. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | ORF Length (bp) | Amino Acid Length (aa) | Mw (kDa) | pI | Instability Index | Grand Average of Hydropathicity |
|---|---|---|---|---|---|---|
| SaMYC1 | 1800 | 599 | 67.32 | 5.69 | 52.17 | −0.358 |
| SaMYC3 | 2022 | 673 | 72.92 | 6.12 | 55.76 | −0.533 |
| SaMYC4 | 2151 | 716 | 77.72 | 5.12 | 64.21 | −0.558 |
| SaMYC5 | 1542 | 513 | 55.71 | 5.69 | 48.26 | −0.404 |
| SabHLH1 | 999 | 332 | 37.11 | 4.67 | 62.30 | −0.456 |
| SabHLH2 | 1302 | 433 | 47.54 | 6.09 | 46.59 | −0.700 |
| SabHLH3 | 927 | 308 | 32.09 | 5.91 | 51.81 | −0.374 |
| SabHLH4 | 708 | 235 | 25.87 | 7.71 | 51.73 | −0.766 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Chen, X.; Xiong, Y.; Niu, M.; Zhang, Y.; Yan, H.; Li, Y.; Zhang, X.; Ma, G. Identification and Functional Analysis of SabHLHs in Santalum album L. Life 2022, 12, 1017. https://doi.org/10.3390/life12071017
Zhang T, Chen X, Xiong Y, Niu M, Zhang Y, Yan H, Li Y, Zhang X, Ma G. Identification and Functional Analysis of SabHLHs in Santalum album L. Life. 2022; 12(7):1017. https://doi.org/10.3390/life12071017
Chicago/Turabian StyleZhang, Ting, Xiaohong Chen, Yuping Xiong, Meiyun Niu, Yueya Zhang, Haifeng Yan, Yuan Li, Xinhua Zhang, and Guohua Ma. 2022. "Identification and Functional Analysis of SabHLHs in Santalum album L." Life 12, no. 7: 1017. https://doi.org/10.3390/life12071017
APA StyleZhang, T., Chen, X., Xiong, Y., Niu, M., Zhang, Y., Yan, H., Li, Y., Zhang, X., & Ma, G. (2022). Identification and Functional Analysis of SabHLHs in Santalum album L. Life, 12(7), 1017. https://doi.org/10.3390/life12071017