

Bacterial Virulence and Prevention for Human Spaceflight

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
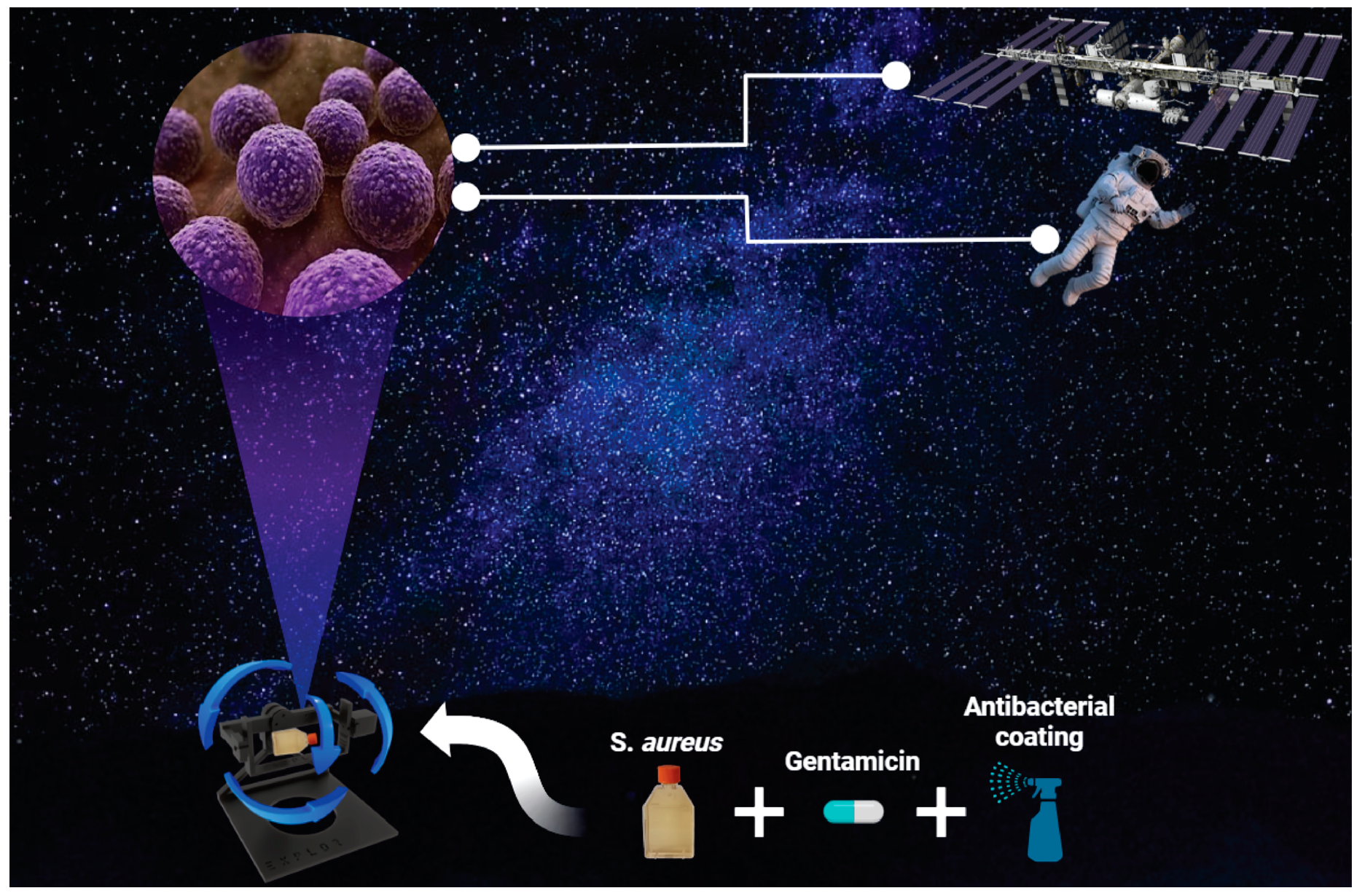
2.1. S. aureus Culture and Maintenance
2.2. Antibiotic Treatment on S. aureus
2.3. Antibacterial Coated Flask
2.4. Cell Viability Analysis of S. aureus
2.5. Live/Dead Analysis of S. aureus
2.6. Image Analysis
2.7. Statistical Analysis
3. Results
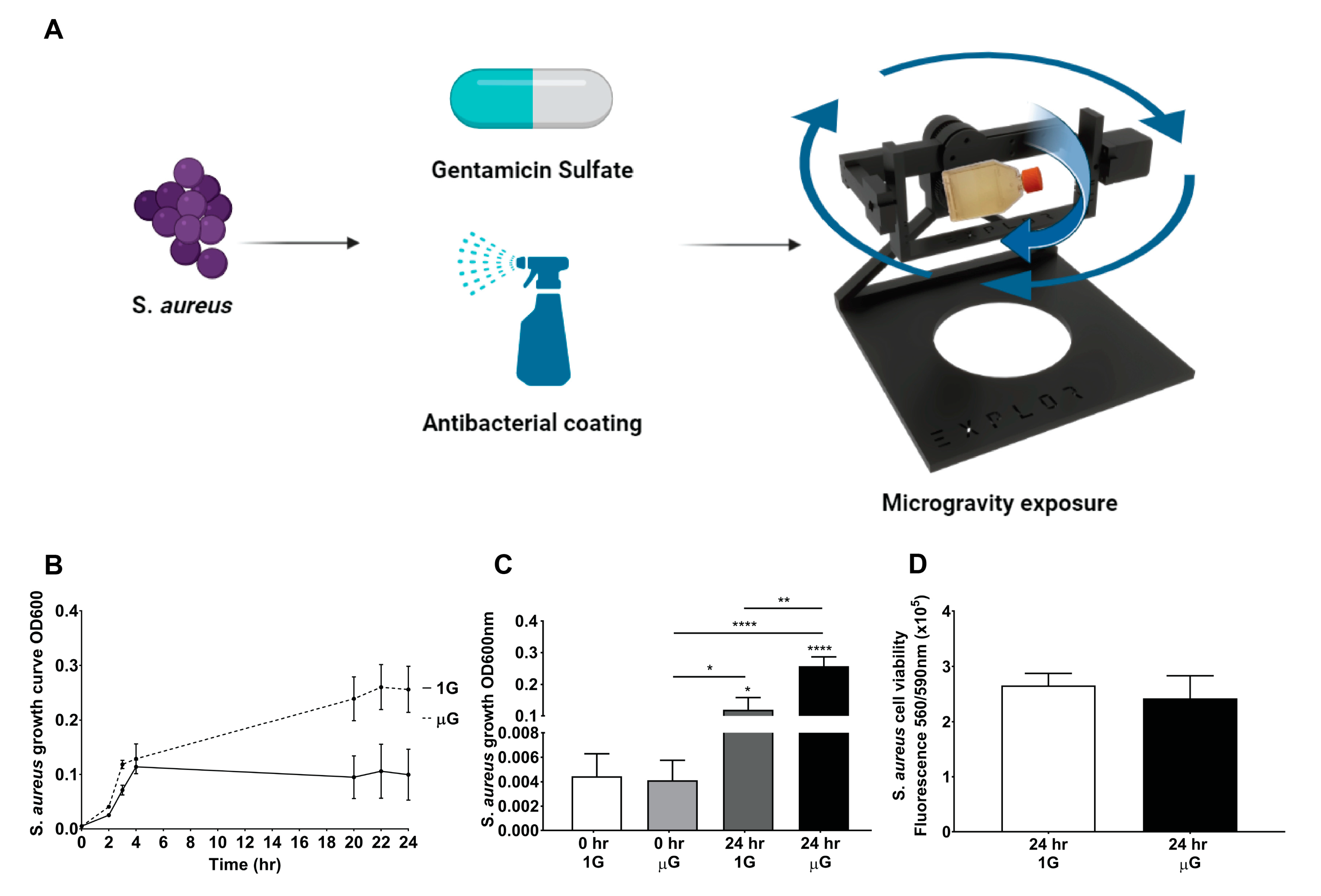
3.1. Significant Increase in S. aureus Proliferation under Simulated Microgravity Conditions
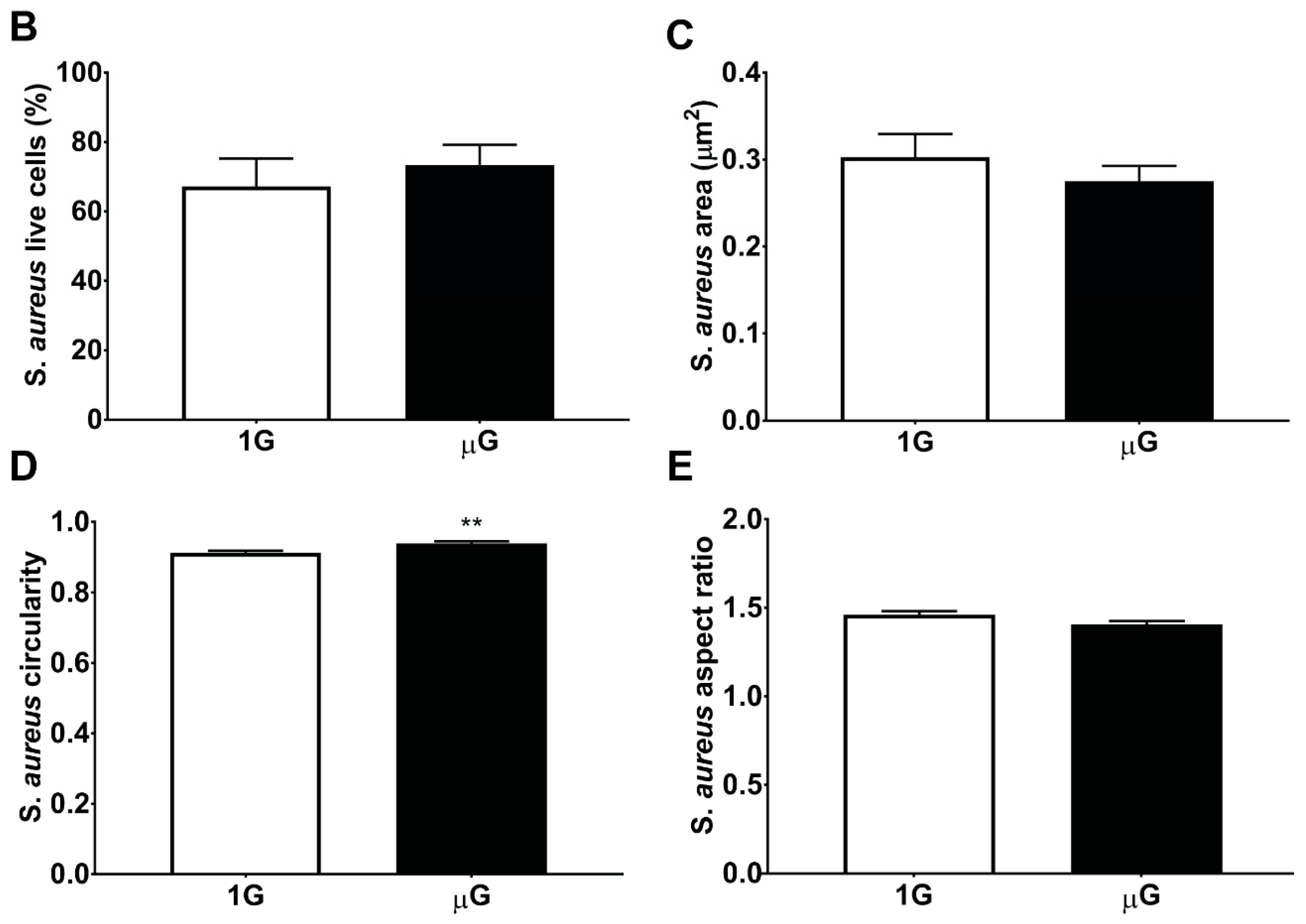
3.2. S. aureus Morphology in Simulated Microgravity Is Comparable to Gravity Conditions
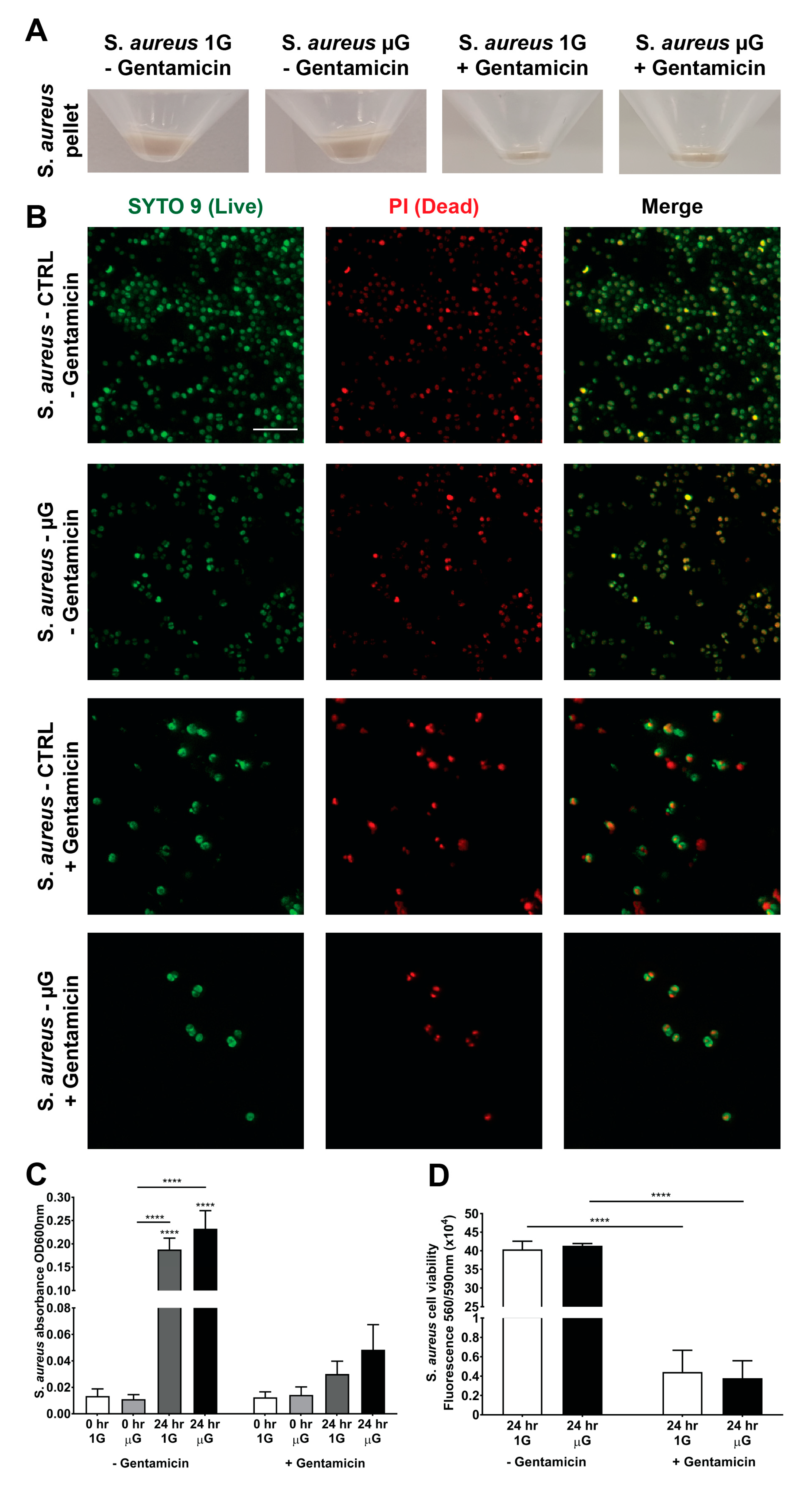
3.3. Gentamicin Suppresses the Growth of S. aureus in Gravity and Simulated Microgravity Conditions
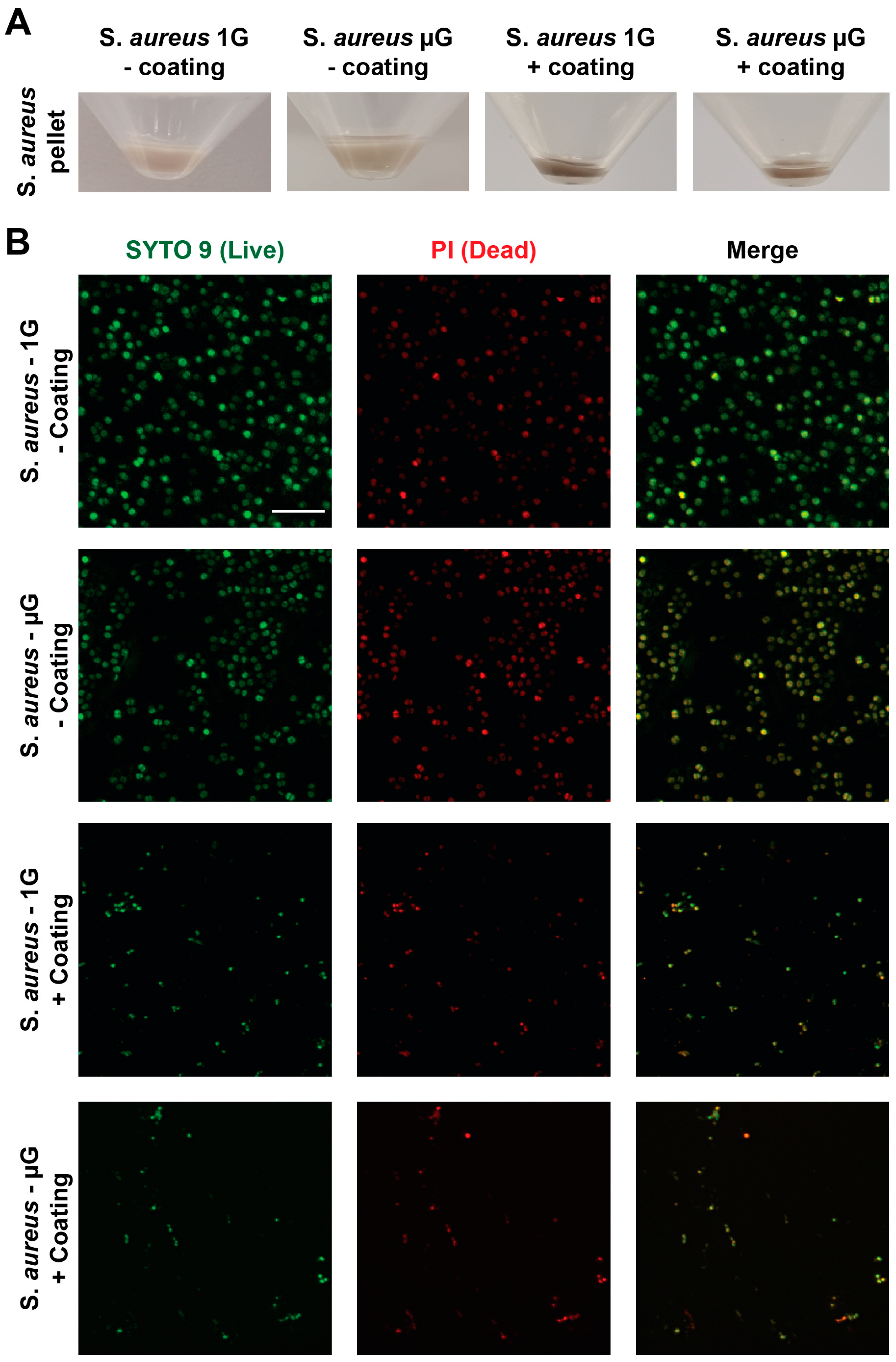
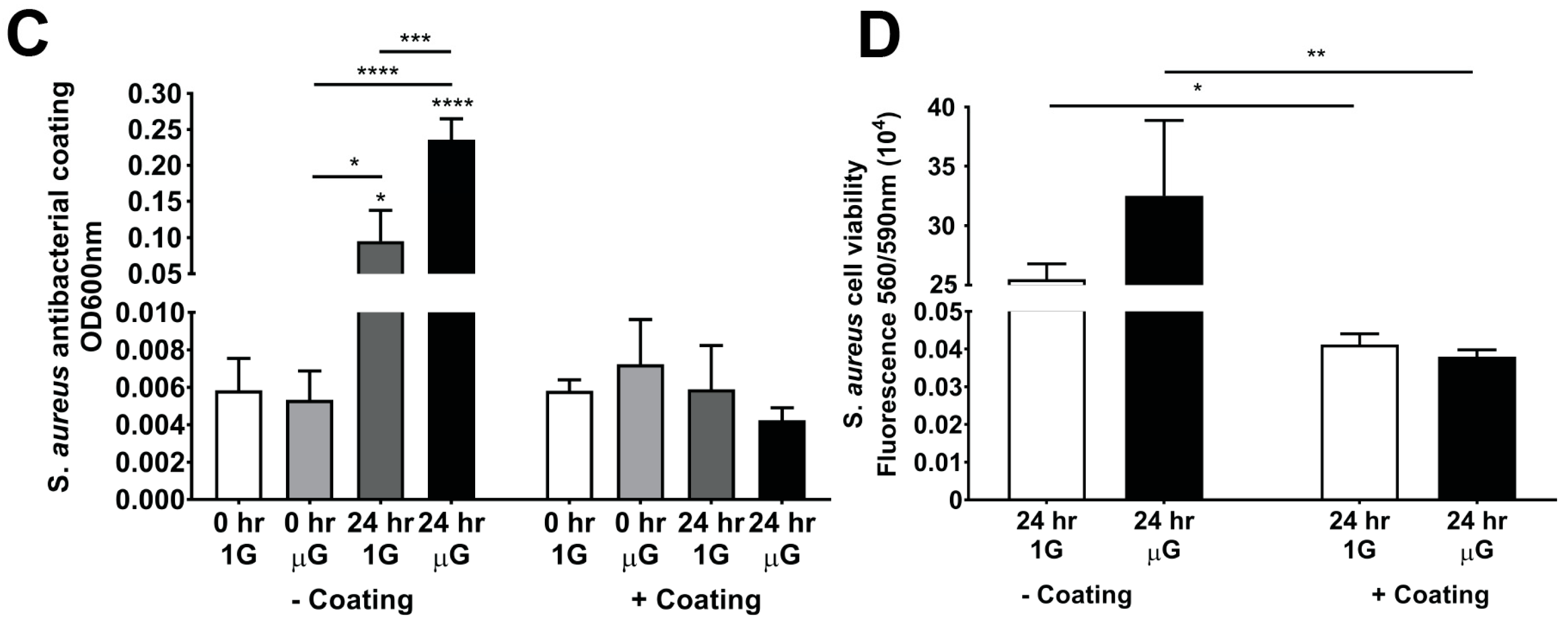
3.4. Antibacterial Coating Inhibits the Growth of S. aureus in Simulated Microgravity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wilkinson, F. The History of Space Exploration. 2022. Available online: https://education.nationalgeographic.org/resource/history-space-exploration (accessed on 16 October 2022).
- Jiang, P.; Green, S.J.; Chlipala, G.E.; Turek, F.W.; Vitaterna, M.H. Reproducible changes in the gut microbiome suggest a shift in microbial and host metabolism during spaceflight. Microbiome 2019, 7, 113. [Google Scholar] [CrossRef] [PubMed]
- Barratt, M.R.; Pool, S.L. Principles of Clinical Medicine for Space Flight; Springer Science & Business Media: Berlin, Germany, 2008. [Google Scholar]
- Voorhies, A.A.; Lorenzi, H.A. The challenge of maintaining a healthy microbiome during long-duration space missions. Front. Astron. Space Sci. 2016, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Castro, S.L.; Nelman-Gonzalez, M.; Nickerson, C.A.; Ott, C.M. Induction of attachment-independent biofilm formation and repression of Hfq expression by low-fluid-shear culture of Staphylococcus aureus. Appl. Environ. Microbiol. 2011, 77, 6368–6378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, R.; Akbar, N.; Khan, N. Gut microbiome and human health under the space environment. J. Appl. Microbiol. 2021, 130, 14–24. [Google Scholar] [CrossRef]
- Garrett-Bakelman, F.E.; Darshi, M.; Green, S.J.; Gur, R.C.; Lin, L.; Macias, B.R.; McKenna, M.J.; Meydan, C.; Mishra, T.; Nasrini, J.; et al. The NASA Twins Study: A multidimensional analysis of a year-long human spaceflight. Science 2019, 364, eaau8650. [Google Scholar] [CrossRef]
- Ritchie, L.E.; Taddeo, S.S.; Weeks, B.R.; Lima, F.; Bloomfield, S.A.; Azcarate-Peril, M.A.; Zwart, S.R.; Smith, S.M.; Turner, N.D. Space environmental factor impacts upon murine colon microbiota and mucosal homeostasis. PLoS ONE 2015, 10, e0125792. [Google Scholar] [CrossRef]
- Smirnov, K.; Lizko, N. Problems of space gastroenterology and microenvironment. Food/Nahrung 1987, 31, 563–566. [Google Scholar] [CrossRef]
- Rosado, H.; Doyle, M.; Hinds, J.; Taylor, P.W. Low-shear modelled microgravity alters expression of virulence determinants of Staphylococcus aureus. Acta Astronaut. 2010, 66, 408–413. [Google Scholar] [CrossRef]
- Kitts, C.; Ronzano, K.; Rasay, R.; Mas, I.; Acain, J.; Neumann, M.; Bica, L.; Mahacek, P.; Minelli, G.; Beck, E.; et al. Initial Flight Results from the PharmaSat Biological Microsatellite Mission. 2009. Available online: https://digitalcommons.usu.edu/smallsat/2009/all2009/27/ (accessed on 20 October 2020).
- Ricco, A.; Parra, M.; Piccini, M.; Ly, D.; Niesel, D.; McGinnis, M.; Kudlicki, A.; Hines, J.; Timucin, L.; Beasley, C.; et al. 2010 Solid-State Sensors, Actuators, and Microsystems Workshop; Transducer Research Foundation: San Diego, CA, USA, 2010; pp. 110–113. [Google Scholar]
- Wilson, J.; Ott, C.M.; Zu Bentrup, K.H.; Ramamurthy, R.; Quick, L.; Porwollik, S.; Cheng, P.; McClelland, M.; Tsaprailis, G.; Radabaugh, T.; et al. Space flight alters bacterial gene expression and virulence and reveals a role for global regulator Hfq. Proc. Natl. Acad. Sci. USA 2007, 104, 16299–16304. [Google Scholar] [CrossRef] [Green Version]
- Holinka, J.; Pilz, M.; Kubista, B.; Presterl, E.; Windhager, R. Effects of selenium coating of orthopaedic implant surfaces on bacterial adherence and osteoblastic cell growth. Bone Jt. J. 2013, 95, 678–682. [Google Scholar] [CrossRef]
- Tsang, S.J.; Simpson, A.H.R. Racing for the Surface; Springer: Berlin, Germany, 2020; pp. 109–169. [Google Scholar]
- Missiakas, D.M.; Schneewind, O. Growth and laboratory maintenance of Staphylococcus aureus. Curr. Protoc. Microbiol. 2013, 28, 9C.1.1–9C.1.9. [Google Scholar] [CrossRef]
- Grosz, M.; Kolter, J.; Paprotka, K.; Winkler, A.-C.; Schäfer, D.; Chatterjee, S.S.; Geiger, T.; Wolz, C.; Ohlsen, K.; Otto, M.; et al. Cytoplasmic replication of S taphylococcus aureus upon phagosomal escape triggered by phenol-soluble modulin α. Cell. Microbiol. 2014, 16, 451–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borst, A.; Van Loon, J.J. Technology and developments for the random positioning machine, RPM. Microgravity Sci. Technol. 2009, 21, 287–292. [Google Scholar] [CrossRef]
- Mann, V.; Grimm, D.; Corydon, T.J.; Krüger, M.; Wehland, M.; Riwaldt, S.; Sahana, J.; Kopp, S.; Bauer, J.; Reseland, J.E.; et al. Changes in human foetal osteoblasts exposed to the random positioning machine and bone construct tissue engineering. Int. J. Mol. Sci. 2019, 20, 1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, B.J.; Tadi, P. Gentamicin; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Allevi. Live/Dead Quantification Using Fiji–Step-By-Step Guide. 2022. Available online: https://www.allevi3d.com/livedead-assay-quantification-fiji/ (accessed on 6 October 2022).
- Nickerson, C.A.; Ott, C.M.; Mister, S.J.; Morrow, B.J.; Burns-Keliher, L.; Pierson, D.L. Microgravity as a novel environmental signal affecting Salmonella enterica serovar Typhimurium virulence. Infect. Immun. 2000, 68, 3147–3152. [Google Scholar] [CrossRef] [Green Version]
- Nickerson, C.A.; Ott, C.M.; Wilson, J.W.; Ramamurthy, R.; Pierson, D.L. Microbial responses to microgravity and other low-shear environments. Microbiol. Mol. Biol. Rev. 2004, 68, 345–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, J.W.; Ott, C.M.; Quick, L.; Davis, R.; zu Bentrup, K.H.; Crabbé, A.; Richter, E.; Sarker, S.; Barrila, J.; Porwollik, S.; et al. Media ion composition controls regulatory and virulence response of Salmonella in spaceflight. PLoS ONE 2008, 3, e3923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, P.W. Impact of space flight on bacterial virulence and antibiotic susceptibility. Infect. Drug Resist. 2015, 8, 249. [Google Scholar] [CrossRef] [Green Version]
- Tixador, R.; Richoilley, G.; Gasset, G.; Planel, H.; Moatti, N.; Lapchine, L.; Enjalbert, L.; Raffin, J.; Bost, R.; Zaloguev, S.; et al. Preliminary results of Cytos 2 experiment. Acta Astronaut. 1985, 12, 131–134. [Google Scholar] [CrossRef]
- Vaishampayan, A.; Grohmann, E. Multi-resistant biofilm-forming pathogens on the International Space Station. J. Biosci. 2019, 44, 125. [Google Scholar] [CrossRef]
- Ilyin, V. Microbiological status of cosmonauts during orbital spaceflights on Salyut and Mir orbital stations. Acta Astronaut. 2005, 56, 839–850. [Google Scholar] [CrossRef]
- Mermel, L.A. Infection prevention and control during prolonged human space travel. Clin. Infect. Dis. 2013, 56, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Shao, D.; Yao, L.; Riaz, M.S.; Zhu, J.; Shi, J.; Jin, M.; Huang, Q.; Yang, H. Simulated microgravity affects some biological characteristics of Lactobacillus acidophilus. Appl. Microbiol. Biotechnol. 2017, 101, 3439–3449. [Google Scholar] [CrossRef]
- Carniello, V.; Peterson, B.W.; van der Mei, H.C.; Busscher, H.J. Role of adhesion forces in mechanosensitive channel gating in Staphylococcus aureus adhering to surfaces. NPJ Biofilms Microbiomes 2020, 6, 31. [Google Scholar] [CrossRef]
- Kuehnert, M.J.; Hill, H.; Kupronis, B.; Tokars, J.; Solomon, S.; Jernigan, D. Methicillin-resistant–Staphylococcus aureus hospitalizations, United States. Emerg. Infect. Dis. 2005, 11, 868. [Google Scholar] [CrossRef]
- Patti, J.M.; Allen, B.L.; McGavin, M.J.; Höök, M. MSCRAMM-mediated adherence of microorganisms to host tissues. Annu. Rev. Microbiol. 1994, 48, 585–617. [Google Scholar] [CrossRef]
- O’Seaghdha, M.; van Schooten, C.; Kerrigan, S.; Emsley, J.; Silverman, G.; Cox, D.; Lenting, P.; Foster, T. Staphylococcus aureus protein A binding to von Willebrand factor A1 domain is mediated by conserved IgG binding regions. FEBS J. 2006, 273, 4831–4841. [Google Scholar] [CrossRef]
- Pawar, P.; Shin, P.K.; Mousa, S.A.; Ross, J.M.; Konstantopoulos, K. Fluid shear regulates the kinetics and receptor specificity of Staphylococcus aureus binding to activated platelets. J. Immunol. 2004, 173, 1258–1265. [Google Scholar] [CrossRef] [Green Version]
- Nauman, E.A.; Ott, C.M.; Sander, E.; Tucker, D.L.; Pierson, D.; Wilson, J.W.; Nickerson, C.A. Novel quantitative biosystem for modeling physiological fluid shear stress on cells. Appl. Environ. Microbiol. 2007, 73, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Allen, L.A.; Kalani, A.; Estante, F.; Rosengren, A.; Stodieck, L.; Klaus, D.; Zea, L. Simulated Micro-, Lunar, and Martian Gravities on Earth—Effects on Escherichia coli Growth, Phenotype, and Sensitivity to Antibiotics. Life 2022, 12, 1399. [Google Scholar] [CrossRef]
- Matin, A.; Wang, J.-H.; Keyhan, M.; Singh, R.; Benoit, M.; Parra, M.; Padgen, M.; Ricco, A.; Chin, M.; Friedericks, C.; et al. Payload hardware and experimental protocol development to enable future testing of the effect of space microgravity on the resistance to gentamicin of uropathogenic Escherichia coli and its σs-deficient mutant. Life Sci. Space Res. 2017, 15, 1–10. [Google Scholar] [CrossRef]
- Savage, C. Smelly Tablets Survive Better in Space. 2022. Available online: https://www.adelaide.edu.au/newsroom/news/list/2022/07/01/smelly-tablets-survive-better-in-space (accessed on 26 October 2022).
- Aunins, T.R.; Erickson, K.E.; Prasad, N.; Levy, S.E.; Jones, A.; Shrestha, S.; Mastracchio, R.; Stodieck, L.; Klaus, D.; Zea, L.; et al. Spaceflight modifies Escherichia coli gene expression in response to antibiotic exposure and reveals role of oxidative stress response. Front. Microbiol. 2018, 9, 310. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wazir, H.U.; Narang, P.; Silvani, G.; Mehner, C.; Poole, K.; Burke, C.; Chou, J. Bacterial Virulence and Prevention for Human Spaceflight. Life 2023, 13, 656. https://doi.org/10.3390/life13030656
Wazir HU, Narang P, Silvani G, Mehner C, Poole K, Burke C, Chou J. Bacterial Virulence and Prevention for Human Spaceflight. Life. 2023; 13(3):656. https://doi.org/10.3390/life13030656
Chicago/Turabian StyleWazir, Hakim Ullah, Pooja Narang, Giulia Silvani, Christine Mehner, Kate Poole, Catherine Burke, and Joshua Chou. 2023. "Bacterial Virulence and Prevention for Human Spaceflight" Life 13, no. 3: 656. https://doi.org/10.3390/life13030656
APA StyleWazir, H. U., Narang, P., Silvani, G., Mehner, C., Poole, K., Burke, C., & Chou, J. (2023). Bacterial Virulence and Prevention for Human Spaceflight. Life, 13(3), 656. https://doi.org/10.3390/life13030656