

Euglena, a Gravitactic Flagellate of Multiple Usages

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
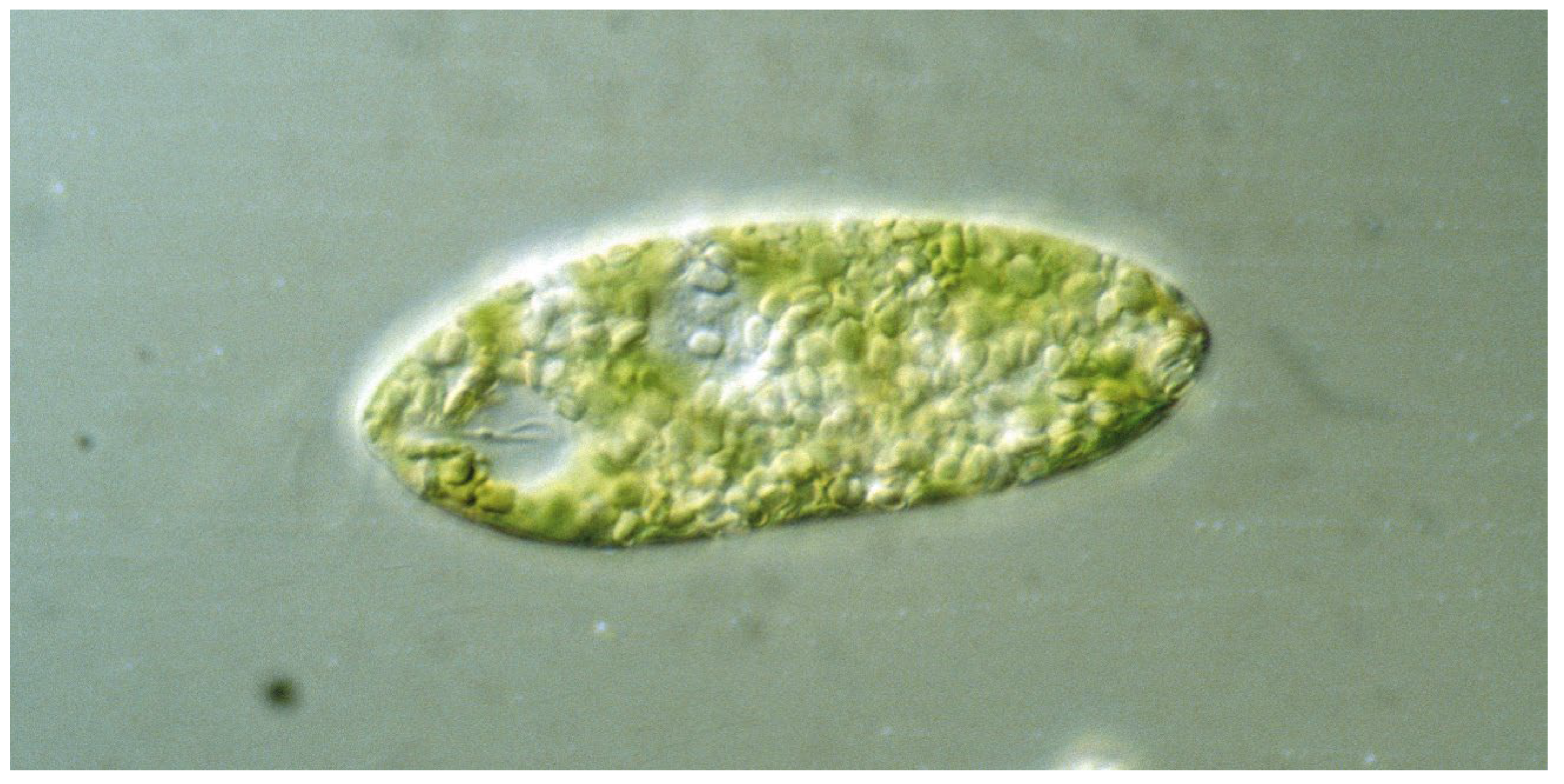
Characteristics of the Genus Euglena
2. Graviperception and Graviresponses in E. gracilis
Physiology of Gravitaxis
3. Gravireceptor and Molecular Sensory Transduction Chain in E. gracilis
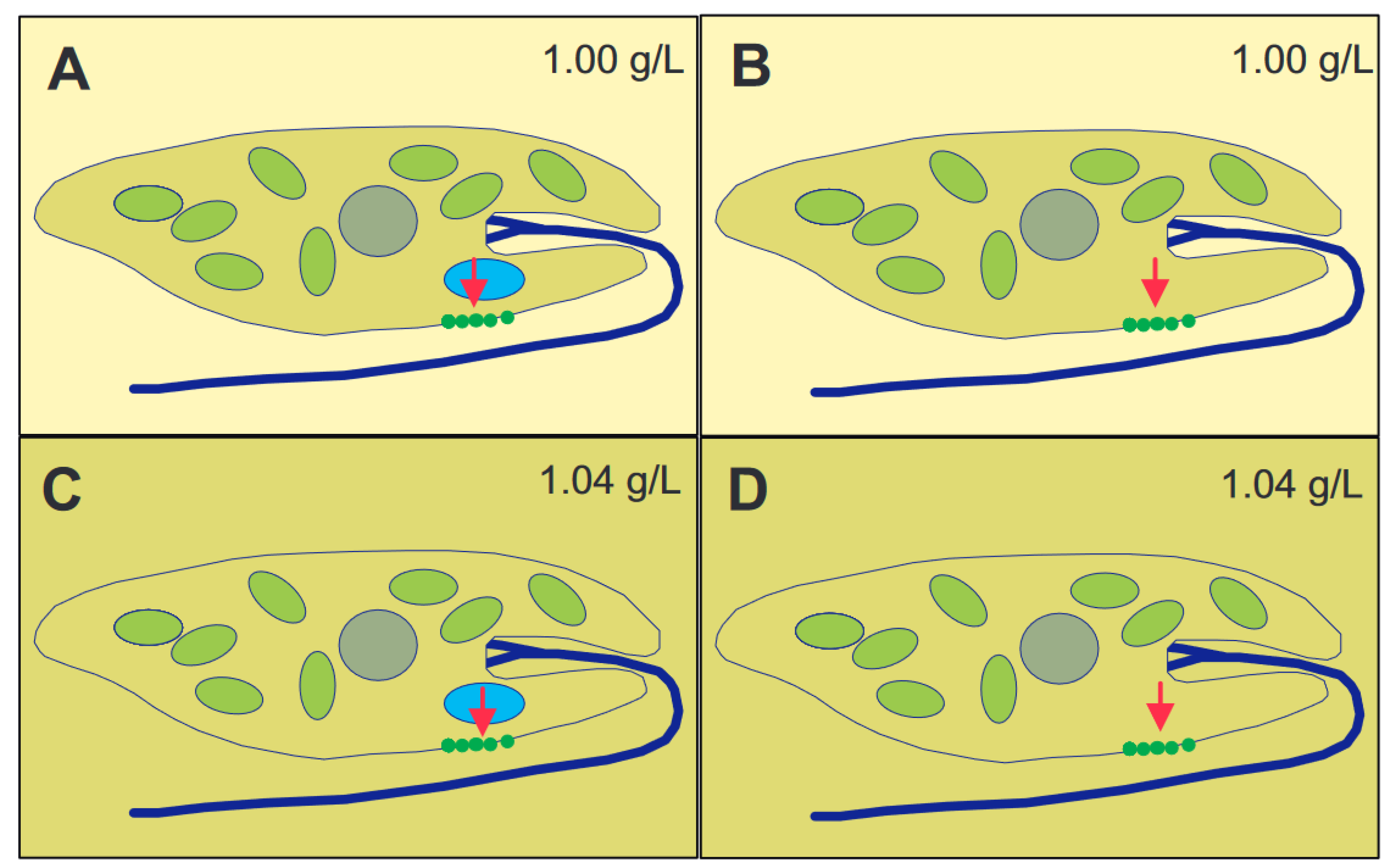
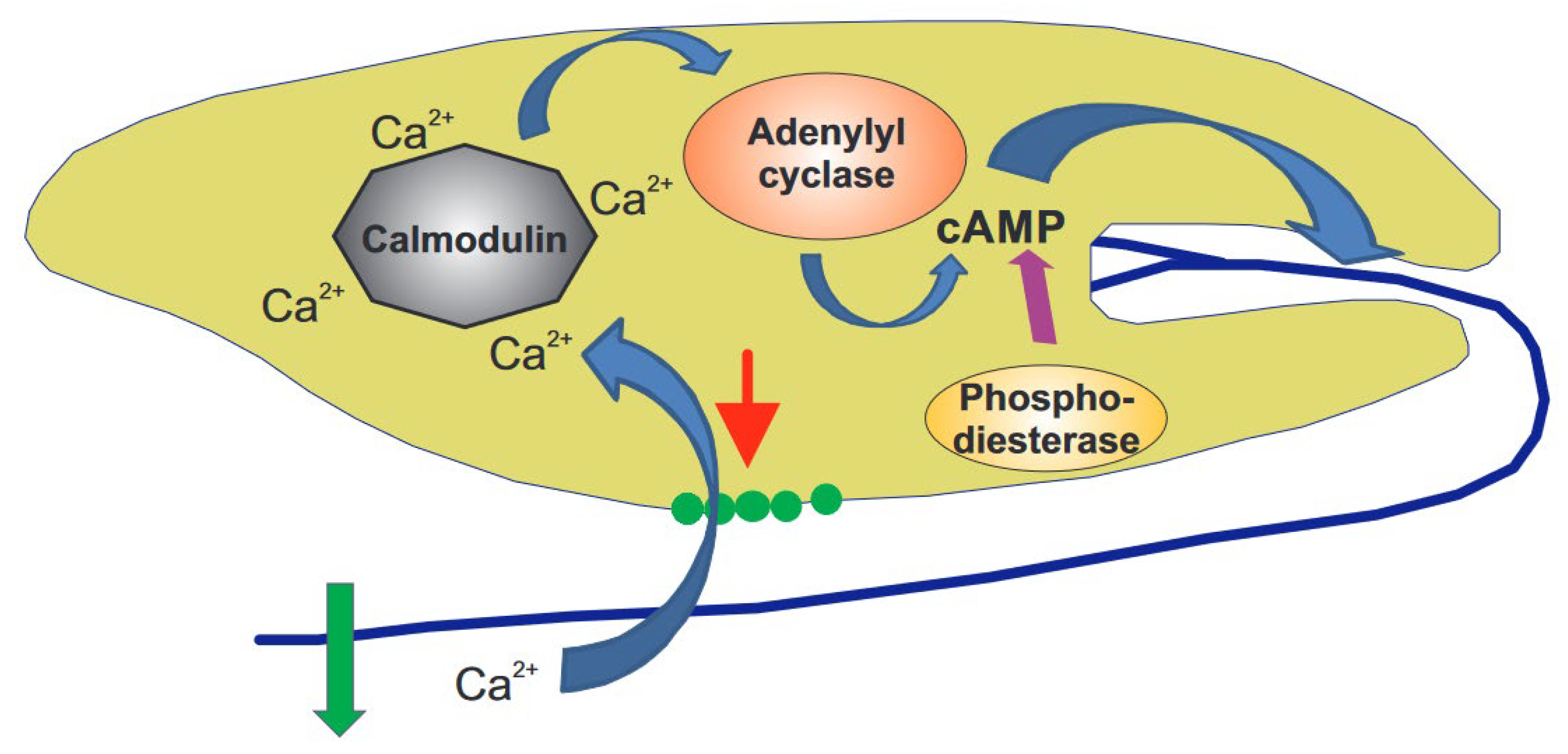
3.1. Mechanosensitive Channels and Calcium
3.2. Calmodulin
3.3. Cyclic Adenosine Monophosphate
3.4. Protein Kinase A and Course Correction
4. Methods to Modify the Influence of Gravity
4.1. Hypergravity as a Tool to Identify Gravity-Related Processes
4.2. How Much Gravity Force Is Needed for a Gravi-Response?
4.3. Neutralization of the Influence of Gravity
5. Real Microgravity
6. Biological Life Support Systems
6.1. Terrestrial Models
6.2. Bioregenerative Life Support System in Space
7. Euglena as Commercial Product and a Source for Medicine
8. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- O’Neill, E.C.; Trick, M.; Henrissat, B.; Field, R.A. Euglena in time: Evolution, control of central metabolic processes and multi-domain proteins in carbohydrate and natural product biochemistry. Perspect. Sci. 2015, 6, 84–93. [Google Scholar] [CrossRef]
- Dobell, C. Antony van Leeuwenhoek and his “Little Animals.” Being Some Account of the Father of Protozoology and Bacteriology and his Multifarious Discoveries in these Disciplines. Collected, Translated, and Edited, from his Printed Works, Unpublished Manuscripts, and Contemporary Records; Published on the 300th Anniversary of his Birth; John Bale, Sons and Daniellson LTD.: London, UK, 1932; p. 435. [Google Scholar]
- Lee, W.J. Three newly recorded heterotrophic euglenids (Protist), Entosiphon oblongum, Euglena longa and Keelungia pulex from South Korea. J. Species Res. 2022, 11, 89–93. [Google Scholar]
- Hindák, F.; Wolowski, K.; Hindáková, A. Cysts and their formation in some neustonic Euglena species. Int. J. Limnol. 2000, 36, 83–93. [Google Scholar] [CrossRef]
- Oliva-Martínez, M.G.; Godínez-Ortega, J.L.; Zuñiga-Ramos, C.A. Biodiversidad del fitoplancton de aguas continentales en México. Rev. Mex. Biodivers. 2014, 85, 54–61. [Google Scholar] [CrossRef]
- Häder, D.-P.; Hoiczyk, E. Gliding motility. In Algal Cell Motility; Melkonian, M., Ed.; Current Phycology; Chapman and Hall: New York, NY, USA, 1992; pp. 1–38. [Google Scholar]
- Häder, D.-P.; Melkonian, M. Phototaxis in the gliding flagellate, Euglena mutabilis. Arch. Microb. 1983, 135, 25–29. [Google Scholar] [CrossRef]
- Wolken, J.J. Euglena: The photoreceptor system for phototaxis. J. Protozool. 1977, 24, 518–522. [Google Scholar]
- Ligęza, S.; Wilk-Woźniak, E. The occurrence of a Euglena pascheri and Lepocinclis ovum bloom in an oxbow lake in southern Poland under extreme environmental conditions. Ecol. Indic. 2011, 11, 925–929. [Google Scholar] [CrossRef]
- Chaudhuri, D.; Ghate, N.B.; Deb, S.; Panja, S.; Sarkar, R.; Rout, J.; Mandal, N. Assessment of the phytochemical constituents and antioxidant activity of a bloom forming microalgae Euglena tuba. Biol. Res. 2014, 47, 1–11. [Google Scholar]
- Gerber, S.; Häder, D.-P. Effects of enhanced UV-B irradiation on the red coloured freshwater flagellate Euglena sanguinea. FEMS Microbiol. Ecol. 1994, 13, 177–184. [Google Scholar]
- Kings, A.J.; Raj, R.E.; Miriam, L.M.; Visvanathan, M.A. Cultivation, extraction and optimization of biodiesel production from potential microalgae Euglena sanguinea using eco-friendly natural catalyst. Energy Convers. Manag. 2017, 141, 224–235. [Google Scholar]
- Sato, N. Complex origins of chloroplast membranes with photosynthetic machineries: Multiple transfers of genes from divergent organisms at different times or a single endosymbiotic event? J. Plant Res. 2020, 133, 15–33. [Google Scholar] [CrossRef]
- Zakryś, B.; Milanowski, R.; Karnkowska, A. Evolutionary origin of Euglena. In Euglena: Biochemistry, Cell and Molecular Biology; Schwartzbach, S., Shigeoka, S., Eds.; Springer: Cham, Switzerland, 2017; pp. 3–17. [Google Scholar]
- Lebert, M.; Häder, D.-P. Behavioral mutants of Euglena gracilis: Functional and spectroscopic characterization. J. Plant Physiol. 1997, 151, 188–195. [Google Scholar] [CrossRef]
- Ozasa, K.; Kang, H.; Song, S.; Tamaki, S.; Shinomura, T.; Maeda, M. Regeneration of the eyespot and flagellum in Euglena gracilis during cell division. Plants 2021, 10, 2004. [Google Scholar] [CrossRef] [PubMed]
- Basterretxea, G.; Font-Munoz, J.S.; Tuval, I. Phytoplankton orientation in a turbulent ocean: A microscale perspective. Front. Mar. Sci. 2020, 7, 185. [Google Scholar] [CrossRef]
- Brodhun, B.; Häder, D.-P. Photoreceptor proteins and pigments in the paraflagellar body of the flagellate Euglena gracilis. Photochem. Photobiol. 1990, 52, 865–871. [Google Scholar] [CrossRef]
- Häder, D.-P.; Iseki, M. Photomovement in Euglena. In Euglena: Biochemistry, Cell and Molecular Biology; Schwartzbach, S., Shigeoka, S., Eds.; Springer: Cham, Switzerland, 2017; pp. 207–235. [Google Scholar]
- Richter, P.R.; Streb, C.; Häder, D.-P. Sign change of phototaxis in Euglena gracilis. Trends Photochem. Photobiol. 2006, 11, 57–61. [Google Scholar]
- Iseki, M.; Matsunaga, S.; Murakami, A.; Ohno, K.; Shiga, K.; Yoshida, C.; Sugai, M.; Takahashi, T.; Hori, T.; Watanabe, M. A blue-light-activated adenylyl cyclase mediates photoavoidance in Euglena gracilis. Nature 2002, 415, 1047–1051. [Google Scholar] [CrossRef]
- Ntefidou, M.; Iseki, M.; Watanabe, M.; Lebert, M.; Häder, D.-P. Photoactivated adenylyl cyclase controls phototaxis in the flagellate Euglena gracilis. Plant Physiol. 2003, 133, 1517–1521. [Google Scholar] [CrossRef]
- Häder, D.-P.; Ntefidou, M.; Iseki, M.; Watanabe, M. Phototaxis photoreceptor in Euglena gracilis. In Light Sensing in Plants; Wada, M., Shimazaki, K., Iino, M., Eds.; Springer: Tokyo, Japan, 2005; pp. 223–229. [Google Scholar]
- Ozasa, K.; Won, J.; Song, S.; Maeda, M. Behavior of Euglena gracilis under simultaneous competing optical and chemical stimuli. Algal Res. 2018, 35, 98–105. [Google Scholar] [CrossRef]
- Checcucci, A.; Colombetti, G.; del Carratore, G.; Ferrara, R.; Lenci, F. Red light induced accumulation of Euglena gracilis. Photochem. Photobiol. 1974, 19, 223–226. [Google Scholar] [CrossRef]
- Tanimoto, Y.; Izumi, S.; Furuta, K.; Suzuki, T.; Fujiwara, Y.; Fujiwara, M.; Hirata, T.; Yamada, S. Effects of high magnetic field on Euglena gracilis. Int. J. Appl. Electromagn. Mech. 2002, 14, 311–316. [Google Scholar] [CrossRef]
- Lebert, M.; Richter, P.; Porst, M.; Häder, D.-P. Mechanism of gravitaxis in the flagellate Euglena gracilis. In Proceedings of the 12th C.E.B.A.S.Workshops. Annual Issue 1996, Bochum, Germany; 1996; pp. 225–234. [Google Scholar]
- Häder, D.-P.; Lebert, M.; Richter, P. Gravitaxis and graviperception in Euglena gracilis. Adv. Space Res. 1998, 21, 1277–1284. [Google Scholar] [CrossRef]
- Ullrich, O.; Häder, D.-P. Editorial. Signal transduction in gravity perception: From microorganisms to mammals. Signal Transduct. 2006, 6, 377–379. [Google Scholar] [CrossRef]
- Hock, B.; Häder, D.-P. Graviresponses in fungi and slime molds. Signal Transduct. 2006, 6, 443–448. [Google Scholar] [CrossRef]
- Hemmersbach, R.; Volkmann, D.; Häder, D.-P. Graviorientation in protists and plants. J. Plant Physiol. 1999, 154, 1–15. [Google Scholar] [CrossRef]
- Ullrich, O.; Thiel, C.S. Gravitational Force: Triggered stress in cells of the immune system. In Stress Challenges and Immunity in Space; Springer: Berlin, Germany, 2012; pp. 187–202. [Google Scholar]
- Platt, J.B. On the specific gravity of Spirostomum, Paramecium and the tadpole in relation to the problem of geotaxis. Am. Nat. 1899, 33, 31. [Google Scholar] [CrossRef]
- Köhler, O. Über die Geotaxis von Paramecium. Verh. Dtsch. Zool. Ges. 1921, 26, 69–71. [Google Scholar]
- Dryl, S. Behavior and motor response of Paramecium. In Paramecium: A Current Survey; van Wagtendonk, W.J., Ed.; Elsevier Scientific: Amsterdam, The Netherlands, 1974; pp. 165–218. [Google Scholar]
- Haupt, W. Geotaxis. In Handbuch der Pflanzenphysiologie; Ruhland, W., Ed.; Springer: Berlin/Heidelberg, Germany, 1962; Volume 17/2, pp. 390–395. [Google Scholar]
- Kuznicki, L. Behavior of Paramecium in gravity fields. I. Sinking of immobilized specimens. Acta Protozool. 1968, 6, 109–117. [Google Scholar]
- Hemmersbach, R.; Voormanns, R.; Häder, D.-P. Graviresponses in Paramecium biaurelia under different accelerations: Studies on the ground and in space. J. Exp. Biol. 1999, 390, 2199–2205. [Google Scholar] [CrossRef]
- Machemer, H.; Bräucker, R. Gravireception and graviresponses in ciliates. Acta Protozool. 1992, 31, 185–214. [Google Scholar]
- Fukui, K.; Asai, H. Negative geotactic behavior of Paramecium caudatum is completely described by the mechanism of buoyancy-oriented upward swimming. Biophys. J. 1985, 47, 479–482. [Google Scholar] [CrossRef]
- Grolig, F.; Herkenrath, H.; Pumm, T.; Gross, A.; Galland, P. Gravity susception by buoyancy: Floating lipid globules in sporangiophores of Phycomyces. Planta 2004, 218, 658–667. [Google Scholar] [CrossRef]
- Grolig, F.; Döring, M.; Galland, P. Gravisusception by buoyancy: A mechanism ubiquitous among fungi? Protoplasma 2006, 229, 117–123. [Google Scholar] [CrossRef]
- Häder, D.-P.; Lebert, M. Photoorientation in photosynthetic flagellates. In Methods in Molecular Biology; Jin, T., Hereld, D., Eds.; Humana Press: Clifton, NJ, USA, 2009; Volume 571, pp. 51–65. [Google Scholar]
- Stallwitz, E.; Häder, D.-P. Motility and phototactic orientation of the flagellate Euglena gracilis impaired by heavy metal ions. J. Photochem. Photobiol. B Biol. 1993, 18, 67–74. [Google Scholar] [CrossRef]
- Stallwitz, E.; Häder, D.-P. Effects of heavy metals on motility and gravitactic orientation of the flagellate, Euglena gracilis. Eur. J. Protistol. 1994, 30, 18–24. [Google Scholar] [CrossRef]
- Richter, P.R.; Ntefidou, M.; Streb, C.; Faddoul, J.; Lebert, M.; Häder, D.-P. High light exposure leads to a sign change of gravitaxis in the flagellate Euglena gracilis. Acta Protozool. 2002, 41, 343–351. [Google Scholar]
- Ntefidou, M.; Richter, P.; Streb, C.; Lebert, M.; Häder, D.-P. High light exposure leads to a sign change in gravitaxis of the flagellate Euglena gracilis. In Proceedings of the Life in Space for Life on Earth. 8th European Symposium on Life Sciences Research in Space. 23rd Annual International Gravitational Physiology Meeting, Karolinska Institutet, Stockholm, Sweden, 2–7 June 2002; pp. 301–302. [Google Scholar]
- Richter, P.R.; Streb, C.; Ntefidou, M.; Lebert, M.; Häder, D.-P. High light-induced sign change of gravitaxis in the flagellate Euglena gracilis is mediated by reactive oxygen species. Acta Protozool. 2003, 42, 197–204. [Google Scholar]
- Häder, D.-P.; Liu, S.M. Motility and gravitactic orientation of the flagellate, Euglena gracilis, impaired by artificial and solar UV-B radiation. Curr. Microbiol. 1990, 21, 161–168. [Google Scholar] [CrossRef]
- Richter, P.; Börnig, A.; Streb, C.; Ntefidou, M.; Lebert, M.; Häder, D.-P. Effects of increased salinity on gravitaxis in Euglena gracilis. J. Plant Physiol. 2003, 160, 651–656. [Google Scholar] [CrossRef]
- Lebert, M.; Häder, D.-P. Negative gravitactic behavior of Euglena gracilis can not be described by the mechanism of buoyancy-oriented upward swimming. Adv. Space Res. 1999, 24, 851–860. [Google Scholar] [CrossRef]
- Staves, M.P. Cytoplasmic streaming and gravity sensing in Chara internodal cells. Planta 1997, 203, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Gadalla, D.; Braun, M.; Böhmer, M. Gravitropism in higher plants: Cellular aspects. In Gravitational Biology—Gravity Sensing and Graviorientation in Microorganisms and Plants; Braun, M., Häder, D.-P., Böhmer, M., Palme, K., Hemmersbach, R., Eds.; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Sack, F.D. Plastids and gravitropic sensing. Planta 1997, 203, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Schnabl, H. Gravistimulated effects in plants. In Astrobiology; Horneck, G., Baumstark-Khan, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar] [CrossRef]
- Lebert, M.; Porst, M.; Richter, P.; Häder, D.-P. Physical characterization of gravitaxis in Euglena gracilis. J. Plant Physiol. 1999, 155, 338–343. [Google Scholar] [CrossRef]
- Lebert, M.; Häder, D.-P. How Euglena tells up from down. Nature 1996, 379, 590. [Google Scholar] [CrossRef]
- Lebert, M.; Richter, P.; Häder, D.-P. Signal perception and transduction of gravitaxis in the flagellate Euglena gracilis. J. Plant Physiol. 1997, 150, 685–690. [Google Scholar] [CrossRef]
- Häder, D.-P.; Hemmersbach, R.; Lebert, M. Gravity and the Behavior of Unicellular Organisms; Cambridge University Press: Cambridge, UK, 2005; pp. 1–258. [Google Scholar]
- Kiyota, M.; Numayama, N.; Goto, K. Circadian rhythms of the L-ascorbic acid level in Euglena and spinach. J. Photochem. Photobiol. B Biol. 2006, 84, 197–203. [Google Scholar] [CrossRef]
- Bolige, A.; Goto, K. High irradiance responses involving photoreversible multiple photoreceptors as related to photoperiodic induction of cell division in Euglena. J. Photochem. Photobiol. B Biol. 2007, 86, 109–120. [Google Scholar] [CrossRef]
- Lebert, M.; Porst, M.; Häder, D.-P. Circadian rhythm of gravitaxis in Euglena gracilis. J. Plant Physiol. 1999, 155, 344–349. [Google Scholar] [CrossRef]
- Nasir, A.; Strauch, S.; Becker, I.; Sperling, A.; Schuster, M.; Richter, P.; Weißkopf, M.; Ntefidou, M.; Daiker, V.; An, Y. The influence of microgravity on Euglena gracilis as studied on Shenzhou 8. Plant Biol. 2014, 16, 113–119. [Google Scholar] [CrossRef]
- Häder, D.-P.; Lebert, M. Graviperception and gravitaxis in algae. Adv. Space Res. 2001, 27, 861–870. [Google Scholar] [CrossRef]
- Richter, P.R.; Strauch, S.M.; Ntefidou, M.; Schuster, M.; Daiker, V.; Nasir, A.; Haag, F.W.M.; Lebert, M. Influence of different light-dark cycles on motility and photosynthesis of Euglena gracilis in closed bioreactors. Astrobiology 2014, 14, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Appendino, G.; Owsianik, G. The transient receptor potential channel TRPA1: From gene to pathophysiology. Pflüg. Arch.—Eur. J. Physiol. 2012, 464, 425–458. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12, 218. [Google Scholar] [CrossRef] [PubMed]
- Häder, D.-P.; Richter, P.; Schuster, M.; Daiker, V.; Lebert, M. Molecular analysis of the graviperception signal transduction in the flagellate Euglena gracilis: Involvement of a transient receptor potential-like channel and a calmodulin. Adv. Space Res. 2009, 43, 1179–1184. [Google Scholar] [CrossRef]
- Borah, M.; Konakalla, N.C. RNAi Technology: A novel platform in crop protection. In Emerging Trends in Plant Pathology; Springer: Berlin, Germany, 2021; pp. 561–575. [Google Scholar]
- Björkman, T. Perception of gravity by plants. Adv. Space Res. 1992, 12, 195–201. [Google Scholar] [CrossRef]
- Weisenseel, M.; Meyer, A.J. Bioelectricity, gravity and plants. Planta 1997, 203, 98–106. [Google Scholar] [CrossRef]
- Häder, D.-P.; Lebert, M.; Richter, P. Gravitaxis and graviperception in flagellates and ciliates. In Proceedings of the 14th ESA Symposium on European Rocket and Balloon Programmes and Related Research (ESA SP-437), Potsdam, Germany, 31 May–3 June 1999; pp. 479–486. [Google Scholar]
- Häder, D.-P.; Hemmersbach, R. Graviperception and graviorientation in flagellates. Planta 1997, 203, 7–10. [Google Scholar] [CrossRef]
- Yoshimura, K. Stimulus perception and membrane excitation in unicellular alga Chlamydomonas. In Coding and Decoding of Calcium Signals in Plants; Springer: Berlin, Germany, 2011; pp. 79–91. [Google Scholar]
- Häder, D.-P.; Richter, P.; Lebert, M. Signal transduction in gravisensing of flagellates. Signal Transduct. 2006, 6, 422–431. [Google Scholar] [CrossRef]
- McLachlan, D.H.; Underwood, G.J.; Taylor, A.R.; Brownlee, C. Calcium release from intracellular stores is necessary for the photophobic response in the benthic diatom Navicula perminuta (Bacillariophyceae). J. Phycol. 2012, 48, 675–681. [Google Scholar] [CrossRef]
- Richter, P.; Lebert, M.; Tahedl, H.; Häder, D.-P. Calcium is involved in the gravitactic orientation in colorless flagellates. J. Plant Physiol. 2001, 158, 689–697. [Google Scholar] [CrossRef]
- Richter, P.R.; Schuster, M.; Wagner, H.; Lebert, M.; Häder, D.-P. Physiological parameters of gravitaxis in the flagellate Euglena gracilis obtained during a parabolic flight campaign. J. Plant Physiol. 2002, 159, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Häder, D.-P.; Lebert, M. Mechanism of gravitactic signal perception and signal transduction of Euglena gracilis. Microgravity News 1998, 11, 14. [Google Scholar]
- Sachs, F.; Morris, C.E. Mechanosensitive ion channels in nonspecialized cells. In Reviews of Physiology and Biochemistry and Pharmacology; Blaustein, M.P., Greger, R., Grunicke, H., Jahn, R., Mendell, L.M., Miyajima, A., Pette, D., Schultz, G., Schweiger, M., Eds.; Springer: Berlin/Heidelberg, Germany, 1998; Volume 132, pp. 1–78. [Google Scholar]
- Haugland, R.P. Handbook of Fluorescent Probes and Research Chemicals; Molecular Probes: Eugene, OR, USA, 1997. [Google Scholar]
- Richter, P.; Lebert, M.; Korn, R.; Häder, D.-P. Possible involvement of the membrane potential in the gravitactic orientation of Euglena gracilis. J. Plant Physiol. 2001, 158, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Lebert, M.; Häder, D.-P. Image analysis: A versatile tool for numerous applications. GIT Imaging Microsc. 1999, 1, 5–6. [Google Scholar]
- Richter, P.R.; Schuster, M.; Meyer, I.; Lebert, M.; Häder, D.-P. Indications for acceleration-dependent changes of membrane potential in the flagellate Euglena gracilis. Protoplasma 2006, 229, 101–108. [Google Scholar] [CrossRef]
- Finichiu, P.G.; James, A.M.; Larsen, L.; Smith, R.A.; Murphy, M.P. Mitochondrial accumulation of a lipophilic cation conjugated to an ionisable group depends on membrane potential, pH gradient and pK a: Implications for the design of mitochondrial probes and therapies. J. Bioenerg. Biomembr. 2013, 45, 165–173. [Google Scholar] [CrossRef]
- Kung, C.; Martinac, B.; Sukharev, S. Mechanosensitive channels in microbes. Annu. Rev. Microbiol. 2010, 64, 313–329. [Google Scholar] [CrossRef]
- Häder, D.-P.; Lebert, M.; Richter, P.; Ntefidou, M. Gravitaxis and graviperception in flagellates. Adv. Space Res. 2003, 31, 2181–2186. [Google Scholar] [CrossRef]
- Tian, W.; Wang, C.; Gao, Q.; Li, L.; Luan, S. Calcium spikes, waves and oscillations in plant development and biotic interactions. Nat. Plants 2020, 6, 750–759. [Google Scholar] [CrossRef]
- Adler, E.M. Bacteria under pressure, calcium channel internalization, and why cockroaches avoid glucose-baited traps. J. Gen. Physiol. 2013, 142, 1–2. [Google Scholar] [CrossRef]
- Panina, S.; Stephan, A.; la Cour, J.M.; Jacobsen, K.; Kallerup, L.K.; Bumbuleviciute, R.; Knudsen, K.V.; Sánchez-González, P.; Villalobo, A.; Olesen, U.H. Significance of calcium binding, tyrosine phosphorylation, and lysine trimethylation for the essential function of calmodulin in vertebrate cells analyzed in a novel gene replacement system. J. Biol. Chem. 2012, 287, 18173–18181. [Google Scholar] [CrossRef] [PubMed]
- Means, A.R. Molecular mechanisms of action of calmodulin. In Recent Progress in Hormone Research: Proceedings of the 1987 Laurentian Hormone Conference; Academic Press: Cambridge, MA, USA, 2013; p. 223. [Google Scholar]
- Shi, J.; Du, X. Identification, characterization and expression analysis of calmodulin and calmodulin-like proteins in Solanum pennellii. Sci. Rep. 2020, 10, 7474. [Google Scholar] [CrossRef] [PubMed]
- Daiker, V.; Häder, D.-P.; Lebert, M. Molecular characterization of calmodulins involved in the signal transduction chain of gravitaxis in Euglena. Planta 2010, 231, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Toda, H.; Yazawa, M.; Yagi, K. Amino acid sequence of calmodulin from Euglena gracilis. Eur. J. Biochem. 1992, 205, 653–660. [Google Scholar] [CrossRef]
- Nasir, A.; Le Bail, A.; Daiker, V.; Klima, J.; Richter, P.; Lebert, M. Identification of a flagellar protein implicated in the gravitaxis in the flagellate Euglena gracilis. Sci. Rep. 2018, 8, 7605. [Google Scholar] [CrossRef]
- Son, Y.K.; Li, H.; Jung, I.D.; Park, Y.-M.; Jung, W.-K.; Kim, H.S.; Choi, I.-W.; Park, W.S. The calmodulin inhibitor and antipsychotic drug trifluoperazine inhibits voltage-dependent K+ channels in rabbit coronary arterial smooth muscle cells. Biochem. Biophys. Res. Commun. 2014, 443, 321–325. [Google Scholar]
- Naccache, P.H. Neutrophil activation and calmodulin antagonists. In Calmodulin Antagonists and Cellular Physiology; Hidaka, H., Hartshorne, D.J., Eds.; Academic Press: Orlando, FL, USA, 1985; pp. 149–160. [Google Scholar]
- Russo, E.; Salzano, M.; De Falco, V.; Mian, C.; Barollo, S.; Secondo, A.; Bifulco, M.; Vitale, M. Calcium/calmodulin-dependent protein kinase II and its endogenous inhibitor α in medullary thyroid cancer. Clin. Cancer Res. 2014, 20, 1513–1520. [Google Scholar] [CrossRef]
- Gitelman, S.E.; Witman, G.B. Purification of calmodulin from Chlamydomonas: Calmodulin occurs in cell bodies and flagella. J. Cell Biol. 1980, 98, 764–770. [Google Scholar] [CrossRef]
- Tahedl, H.; Richter, P.; Lebert, M.; Häder, D.-P. cAMP is involved in gravitaxis signal transduction of Euglena gracilis. Microgravity Sci. Technol. 1998, 11, 173–178. [Google Scholar]
- Braun, M.; Böhmer, M.; Häder, D.P.; Hemmersbach, R.; Palme, K. Gravitational Biology I: Gravity Sensing and Graviorientation in Microorganisms and Plants; Springer: Cham, Switzerland, 2018; pp. 1–122. [Google Scholar]
- Yang, X.C.; Sachs, F. Block of stretch-activated ion channels in Xenopus oocytes by gadolinium and calcium ions. Science 1989, 243, 1068–1071. [Google Scholar] [CrossRef]
- Spoto, G.; Papponetti, M.; Barbacane, R.C.; Repola, D.; Beradi, S. Caffein, theophilline, and bamifylline are similar as competive inhibitors of 3′,5′-cyclic AMP phosphodiesterase in vitro. Int. J. Immunopathol. Pharmacol. 1997, 10, 153. [Google Scholar]
- Häder, D.-P.; Lebert, M. Graviorientation in flagellates. In Proceedings of the 2nd China-Germany Workshop on Microgravity Sciences, Dunhuang, China, 1–3 September 2002; pp. 189–194. [Google Scholar]
- Wang, J.; Sun, Y.; Tomura, H.; Okajima, F. Ovarian cancer G-protein-coupled receptor 1 induces the expression of the pain mediator prostaglandin E2 in response to an acidic extracellular environment in human osteoblast-like cells. Int. J. Biochem. Cell Biol. 2012, 44, 1937–1941. [Google Scholar] [CrossRef] [PubMed]
- Richter, P.; Ntefidou, M.; Streb, C.; Lebert, M.; Häder, D.-P. Physiological characterization of gravitaxis in Euglena gracilis. J. Gravit. Physiol. 2002, 9, 279–280. [Google Scholar]
- Schwer, C.I.; Lehane, C.; Guelzow, T.; Zenker, S.; Strosing, K.M.; Spassov, S.; Erxleben, A.; Heimrich, B.; Buerkle, H.; Humar, M. Thiopental inhibits global protein synthesis by repression of eukaryotic elongation factor 2 and protects from hypoxic neuronal cell death. PLoS ONE 2013, 8, e77258. [Google Scholar] [CrossRef]
- Hemmersbach, R.; Braun, M. Gravity-sensing and gravity-related signaling pathways in unicellular model systems of protists and plants. Signal Transduct. 2006, 6, 432–442. [Google Scholar] [CrossRef]
- Bräucker, R.; Cogoli, A.; Hemmersbach, R. Graviperception and graviresponse at the cellular level. In Astrobiology: The Quest for the Conditions of Life; Baumstark-Khan, C., Horneck, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 284–297. [Google Scholar]
- Renart, M.F.; Sebastian, J.; Mato, J.M. Adenylate cyclase activity in permeabilized cells from Dictyostelium discoideum. Cell Biol. Int. Rep. 1981, 5, 1045–1054. [Google Scholar] [CrossRef]
- Sultana, H.; Neelakanta, G.; Rivero, F.; Blau-Wasser, R.; Schleicher, M.; Noegel, A.A. Ectopic expression of cyclase associated protein CAP restores the streaming and aggregation defects of adenylyl cyclase a deficient Dictyostelium discoideum cells. BMC Dev. Biol. 2012, 12, 3. [Google Scholar] [CrossRef]
- Steck, R.; Hill, S.; Robison, R.A.; O’Neill, K.L. Pharmacological reversal of caffeine-mediated phagocytic suppression. Cancer Res. 2014, 74, 4861. [Google Scholar] [CrossRef]
- Cameron, R.; Baillie, G.S. cAMP-specific phosphodiesterases: Modulation, inhibition, and activation. In Therapeutic Targets: Modulation, Inhibition, and Activation; Botana, L.M., Loza, M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; p. 1. [Google Scholar]
- Streb, C.; Richter, P.; Ntefidou, M.; Lebert, M.; Häder, D.-P. Sensory transduction of gravitaxis in Euglena gracilis. J. Plant Physiol. 2002, 159, 855–862. [Google Scholar] [CrossRef]
- Favaro, E.; Granata, R.; Miceli, I.; Baragli, A.; Settanni, F.; Perin, P.C.; Ghigo, E.; Camussi, G.; Zanone, M. The ghrelin gene products and exendin-4 promote survival of human pancreatic islet endothelial cells in hyperglycaemic conditions, through phosphoinositide 3-kinase/Akt, extracellular signal-related kinase (ERK) 1/2 and cAMP/protein kinase A (PKA) signalling pathways. Diabetologia 2012, 55, 1058–1070. [Google Scholar]
- Chang, S.C.; Kaufman, P.B. Effects of staurosporine, okadaic acid and sodium fluoride on protein phophorylation in graviresponding oat shoot pulvini. Plant Physiol. Biochem. 2000, 38, 315–323. [Google Scholar] [CrossRef]
- Häder, D.-P.; Faddoul, J.; Lebert, M.; Richter, P.; Schuster, M.; Richter, R.; Strauch, S.M.; Daiker, V.; Sinha, R.; Sharma, N. Investigation of gravitaxis and phototaxis in Euglena gracilis. In Advances in Life Sciences; Sinha, R., Sharma, N.K., Rai, A.K., Eds.; IK International Publishing House: New Delhi, India, 2010; pp. 117–131. [Google Scholar]
- Daiker, V.; Häder, D.-P.; R., R.P.; Lebert, M. The involvement of a protein kinase in phototaxis and gravitaxis of Euglena gracilis. Planta 2011, 233, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Tsang, A.C.; Lam, A.T.; Riedel-Kruse, I.H. Polygonal motion and adaptable phototaxis via flagellar beat switching in the microswimmer Euglena gracilis. Nat. Phys. 2018, 14, 1216–1222. [Google Scholar] [CrossRef]
- Hammond, M.; Zoltner, M.; Garrigan, J.; Butterfield, E.; Varga, V.; Lukeš, J.; Field, M.C. The distinctive flagellar proteome of Euglena gracilis illuminates the complexities of protistan flagella adaptation. New Phytol. 2021, 232, 1323–1336. [Google Scholar] [CrossRef]
- Nasir, A.; Richter, P.R.; Le Bail, A.; Daiker, V.; Stoltze, J.; Prasad, B.; Strauch, S.M.; Lebert, M. Molecular cross-talk between gravity-and light-sensing mechanisms in Euglena gracilis. Int. J. Mol. Sci. 2022, 23, 2776. [Google Scholar] [CrossRef]
- Häder, D.-P. Rock ‘n’ Roll—Wie Mikroorganismen die Schwerkraft spüren. In Spektrum der Wissenschaft Extra: Schwerelos Europa Forscht im Weltall; Spektrum der Wissenschaft mbH: Heidelberg, Germany, 2010; pp. 114–120. [Google Scholar]
- Frett, T.; Petrat, G.; van Loon, J.J.; Hemmersbach, R.; Anken, R. Hypergravity facilities in the ESA ground-based facility program–currnt research activities and future tasks. Microgravity Sci. Technol. 2016, 28, 205–214. [Google Scholar] [CrossRef]
- Häder, D.-P.; Reinecke, E.; Vogel, K.; Kreuzberg, K. Responses of the photosynthetic flagellate, Euglena gracilis, to hypergravity. Eur. Biophys. J. 1991, 20, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Volkmann, D.; Sievers, A. Graviperception in multicellular organs. In Encyclopedia of Plant Physiology. N.S. Physiology of Movements; Haupt, W., Feinleib, M.E., Eds.; Springer: Berlin/Heidelberg, Germany, 1979; Volume 7, pp. 573–600. [Google Scholar]
- Friedrich, U.L.D.; Joop, O.; Pütz, C.; Willich, G. The slow rotating centrifuge microscope NIZEMI—A versatile instrument for terrestrial hypergravity and space microgravity research in biology and materials science. J. Biotechnol. 1996, 47, 225–238. [Google Scholar] [CrossRef]
- Brinckmann, E. BIOLAB, EPU and EMCS for cell culture experiments on the ISS. J. Gravit. Physiol. J. Int. Soc. Gravit. Physiol. 2004, 11, 67–74. [Google Scholar]
- Schuber, M.; Seibt, D.; Anken, R. BIOLAB on the international space station (ISS): Facility and experiments. Curr. Biotechnol. 2013, 2, 201–207. [Google Scholar] [CrossRef]
- Pletser, V.; Winter, J.; Duclos, F.; Bret-Dibat, T.; Friedrich, U.; Clervoy, J.-F.; Gharib, T.; Gai, F.; Minster, O.; Sundblad, P. The first joint european partial-g parabolic flight campaign at moon and Mars gravity levels for science and exploration. Microgravity Sci. Technol. 2012, 24, 383–395. [Google Scholar] [CrossRef]
- Hauslage, J.; Strauch, S.M.; Eßmann, O.; Haag, F.W.; Richter, P.; Krüger, J.; Stoltze, J.; Becker, I.; Nasir, A.; Bornemann, G. Eu: CROPIS–“Euglena gracilis: Combined Regenerative Organic-food Production in Space”-A Space Experiment Testing Biological Life Support Systems Under Lunar and Martian Gravity. Microgravity Sci. Technol. 2018, 30, 933–942. [Google Scholar] [CrossRef]
- Richter, P.R.; Lebert, M.; Tahedl, H.; Häder, D.-P. Physiological characterization of gravitaxis in Euglena gracilis and Astasia longa studied on sounding rocket flights. Adv. Space Res. 2001, 27, 983–988. [Google Scholar] [CrossRef]
- Häder, D.-P.; Richter, P.; Ntefidou, M.; Lebert, M. Gravitational sensory transduction chain in flagellates. Adv. Space Res. 2005, 36, 1182–1188. [Google Scholar] [CrossRef]
- Hemmersbach, R.; Simon, A.; Waßer, K.; Hauslage, J.; Christianen, P.C.; Albers, P.W.; Lebert, M.; Richter, P.; Alt, W.; Anken, R. Impact of a high magnetic field on the orientation of gravitactic unicellular organisms—A critical consideration about the application of magnetic fields to mimic functional weightlessness. Astrobiology 2014, 14, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Brungs, S.; Egli, M.; Wuest, S.L.; Christianen, P.C.; van Loon, J.J.; Anh, T.J.N.; Hemmersbach, R. Facilities for simulation of microgravity in the ESA ground-based facility programme. Microgravity Sci. Technol. 2016, 28, 191–203. [Google Scholar] [CrossRef]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.; Lebert, M. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ferranti, F.; Del Bianco, M.; Pacelli, C. Advantages and limitations of current microgravity platforms for space biology research. Appl. Sci. 2020, 11, 68. [Google Scholar] [CrossRef]
- Poon, C. Factors implicating the validity and interpretation of mechanobiology studies in simulated microgravity environments. Eng. Rep. 2020, 2, e12242. [Google Scholar] [CrossRef]
- Briegleb, W. Some qualitative and quantitative aspects of the fast-rotating clinostat as a research tool. ASGSB Bull. 1992, 5, 23–30. [Google Scholar]
- Borst, A.; Van Loon, J.J. Technology and developments for the random positioning machine, RPM. Microgravity Sci. Technol. 2009, 21, 287–292. [Google Scholar] [CrossRef]
- Wuest, S.L.; Richard, S.; Kopp, S.; Grimm, D.; Egli, M. Simulated microgravity: Critical review on the use of random positioning machines for mammalian cell culture. BioMed Res. Int. 2015, 2015, 971474. [Google Scholar] [CrossRef]
- Hauslage, J.; Cevik, V.; Hemmersbach, R. Pyrocystis noctiluca represents an excellent bioassay for shear forces induced in ground-based microgravity simulators (clinostat and random positioning machine). NPJ Microgravity 2017, 3, 12. [Google Scholar] [CrossRef]
- Wuest, S.L.; Stern, P.; Casartelli, E.; Egli, M. Fluid dynamics appearing during simulated microgravity using random positioning machines. PLoS ONE 2017, 12, e0170826. [Google Scholar]
- Van Loon, J.J. Mechanomics and physicomics in gravisensing. Microgravity Sci. Technol. 2009, 21, 159–167. [Google Scholar] [CrossRef]
- Häder, D.-P.; Braun, M.; Grimm, D.; Hemmersbach, R. Gravireceptors in eukaryotes—A comparison of case studies on the cellular level. NPJ Microgravity 2017, 3, 1–8. [Google Scholar] [CrossRef]
- Hammond, T.G.; Allen, P.L.; Gunter, M.A.; Chiang, J.; Giaever, G.; Nislow, C.; Birdsall, H.H. Physical forces modulate oxidative status and stress defense meditated metabolic adaptation of yeast colonies: Spaceflight and microgravity simulations. Microgravity Sci. Technol. 2018, 30, 195–208. [Google Scholar] [CrossRef]
- Vogel, K.; Hemmersbach-Krause, R.; Kühnel, C.; Häder, D.-P. Swimming behavior of the unicellular flagellate, Euglena gracilis, in simulated and real microgravity. Microgravity Sci. Technol. 1993, 4, 232–237. [Google Scholar]
- Hemmersbach, R.; Strauch, S.M.; Seibt, D.; Schuber, M.; Häder, D.-P. Comparative studies on gravisensitive protists on ground (2D and 3D clinostats) and in microgravity. Microgravity Sci. Technol. 2006, 18, 257–259. [Google Scholar] [CrossRef]
- Strauch, S.; Richter, P.; Schuster, M.; Häder, D.-P. The beating pattern of the flagellum of Euglena gracilis under altered gravity during parabolic flights. J. Plant Physiol. 2010, 167, 41–46. [Google Scholar] [CrossRef]
- Krüger, J.; Richter, P.; Stoltze, J.; Strauch, S.M.; Krüger, M.; Daiker, V.; Prasad, B.; Sonnewald, S.; Reid, S.; Lebert, M. Changes of gene expression in Euglena gracilis obtained during the 29th DLR parabolic flight campaign. Sci. Rep. 2019, 9, 14260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazendic-Galloway, J.; Overton, T. The Mars challenge. Australas. Sci. 2017, 38, 14. [Google Scholar]
- Kamire, R.; Yates, S. Electrochemical Carbon Dioxide Converter and Liquid Regenerator. U.S. Patent 10,941,497, 9 March 2021. [Google Scholar]
- Moores, J.E.; Lemmon, M.T.; Rafkin, S.C.; Francis, R.; Pla-Garcia, J.; de la Torre Juárez, M.; Bean, K.; Kass, D.; Haberle, R.; Newman, C. Atmospheric movies acquired at the Mars Science Laboratory landing site: Cloud morphology, frequency and significance to the Gale Crater water cycle and Phoenix mission results. Adv. Space Res. 2015, 55, 2217–2238. [Google Scholar] [CrossRef]
- Häder, D.-P.; Kreuzberg, K. Algal bioreactor-concept and experiment design. In Proceedings of the Workshop (DARA/CNES) on Artificial Ecological Systems, Marseille, France, 24–26 October 1990; pp. 211–221. [Google Scholar]
- Lebert, M.; Häder, D.-P. Aquarack: Long-term growth facility for ‘professional’ gravisensing cells. In Proceedings of the 2nd European Symposium on the Utilisation of the International Space Station, ESTEC, (ESA-SP 433), Noordwijk, The Netherlands, 16–18 November 1998; pp. 533–537. [Google Scholar]
- Porst, M.; Lebert, M.; Häder, D.-P. Long-term cultivation of the flagellate Euglena gracilis. Microgravity Sci. Technol. 1997, 10, 166–169. [Google Scholar] [PubMed]
- Strauch, S.; Schuster, M.; Lebert, M.; Richter, P.; Schmittnagel, M.; Häder, D.-P. A closed ecological system in a space experiment. In Proceedings of the Life in Space for Life on Earth, Angers, France, 22–27 June 2008; p. 41. [Google Scholar]
- Häder, D.-P.; Richter, P.R.; Strauch, S.M.; Schuster, M. Aquacells—Flagellates under long-term microgravity and potential usage for life support systems. Microgravity Sci. Technol. 2006, 18, 210–214. [Google Scholar] [CrossRef]
- Strauch, S.M. Euglena gracilis als Sauerstoffproduzent Eines Bioregenerativen Lebenserhaltungssystems und Ihre Physiologische Reaktion auf Änderungen der Schwerkraft. Ph.D. Thesis, Friedrich-Alexander Universität, Erlangen, Germany, 2009. [Google Scholar]
- Häder, D.-P.; Braun, M.; Hemmersbach, R. Bioregenerative life support systems in space research. In Gravitational Biology I; Springer: Berlin/Heidelberg, Germany, 2018; pp. 113–122. [Google Scholar]
- Fischer, J.; Schoppmann, K.; Laforsch, C. Life history responses and feeding behavior of Microcrustacea in altered gravity–Applicability in bioregenerative life support systems (BLSS). Microgravity Sci. Technol. 2017, 29, 241–249. [Google Scholar] [CrossRef]
- Fahrion, J.; Mastroleo, F.; Dussap, C.-G.; Leys, N. Use of photobioreactors in regenerative life support systems for human space exploration. Front. Microbiol. 2021, 12, 1748. [Google Scholar] [CrossRef]
- Hauslage, J.; Lebert, M.; Müller, H. Eu:CROPIS—Euglena and Combined Regenerative Organic-food Production in Space. In Proceedings of the Life in Space for Life on Earth, Waterloo, ON, Canada, 16–19 June 2014. [Google Scholar]
- Savage, C.J.; Tan, G.B.T.; Lasseur, C. ESA developments in life support technology: Achievements and future priorities. Acta Astronaut. 2001, 49, 331–344. [Google Scholar] [CrossRef]
- Volponi, M.; Lasseur, C. Considerations on life support systems for interstellar travel: A regenerative story. Acta Fut 2020, 12, 133–149. [Google Scholar]
- Verbeelen, T.; Leys, N.; Ganigué, R.; Mastroleo, F. Development of nitrogen recycling strategies for bioregenerative life support systems in space. Front. Microbiol. 2021, 12, 700810. [Google Scholar] [CrossRef]
- Hong, L.; Zhikai, Y.; Yuming, F. Research Status and Development Trend of Bio-regenerative Life Support System for Deep Space Exploration. J. Deep. Space Explor. 2020, 7, 489–499. [Google Scholar] [CrossRef]
- Suzuki, K. Large-scale cultivation of Euglena. In Euglena: Biochemistry, Cell and Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2017; pp. 285–293. [Google Scholar]
- Gupta, S.P.; Gupta, V.K.; Minhas, U.; Kumar, R.; Sharma, B. Euglena species: Bioactive compounds and their varied applications. Curr. Top. Med. Chem. 2021, 21, 2620–2633. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Yuan, W.; Pei, Z.; Wu, Q.; Mao, E. Microalgae mass production methods. Trans. ASABE 2009, 52, 1275–1287. [Google Scholar] [CrossRef]
- Pawar, S. Effectiveness mapping of open raceway pond and tubular photobioreactors for sustainable production of microalgae biofuel. Renew Sustain. Energy Rev. 2016, 62, 640–653. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, J.Y.; Oh, J.-J.; Jeon, M.S.; An, H.S.; Jin, C.R.; Choi, Y.-E. A strategic approach to apply bacterial substances for increasing metabolite productions of Euglena gracilis in the bioreactor. Appl. Microbiol. Biotechnol. 2021, 105, 5395–5406. [Google Scholar] [CrossRef]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA); Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A. Safety of dried whole cell Euglena gracilis as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2020, 18, e06100. [Google Scholar]
- Watanabe, F.; Yoshimura, K.; Shigeoka, S. Biochemistry and physiology of vitamins in Euglena. Euglena Biochem. Cell Mol. Biol. 2017, 65–90. [Google Scholar] [CrossRef]
- Takeyama, H.; Kanamaru, A.; Yoshino, Y.; Kakuta, H.; Kawamura, Y.; Matsunaga, T. Production of antioxidant vitamins, β-carotene, vitamin C, and vitamin E, by two-step culture of Euglena gracilis Z. Biotechnol. Bioeng. 1997, 53, 185–190. [Google Scholar] [CrossRef]
- Ishii, N.; Koiso, H.; Sakashita, T.; Fuma, S.; Takeda, H.; Uchida, S. Determination of trace elements in Euglena gracilis Z and its bleached mutant strains. Int. J. PIXE 2006, 16, 77–83. [Google Scholar] [CrossRef]
- Gissibl, A.; Sun, A.; Care, A.; Nevalainen, H.; Sunna, A. Bioproducts from Euglena gracilis: Synthesis and applications. Front. Bioeng. Biotechnol. 2019, 7, 108. [Google Scholar] [CrossRef]
- Guo, Q.; Bi, D.; Wu, M.; Yu, B.; Hu, L.; Liu, C.; Gu, L.; Zhu, H.; Lei, A.; Xu, X. Immune activation of murine RAW264. 7 macrophages by sonicated and alkalized paramylon from Euglena gracilis. BMC Microbiol. 2020, 20, 171. [Google Scholar] [CrossRef]
- Harada, R.; Nomura, T.; Yamada, K.; Mochida, K.; Suzuki, K. Genetic engineering strategies for Euglena gracilis and its industrial contribution to sustainable development goals: A review. Front. Bioeng. Biotech. 2020, 8, 790. [Google Scholar] [CrossRef]
- Khatiwada, B.; Sunna, A.; Nevalainen, H. Molecular tools and applications of Euglena gracilis: From biorefineries to bioremediation. Biotech. Bioeng. 2020, 117, 3952–3967. [Google Scholar] [CrossRef]
- Becker, I.; Prasad, B.; Ntefidou, M.; Daiker, V.; Richter, P.; Lebert, M. Agrobacterium tumefaciens-mediated nuclear transformation of a biotechnologically important microalga—Euglena gracilis. Int. J. Mol. Sci. 2021, 22, 6299. [Google Scholar] [CrossRef]
- Nomura, T.; Inoue, K.; Uehara-Yamaguchi, Y.; Yamada, K.; Iwata, O.; Suzuki, K.; Mochida, K. Highly efficient transgene-free targeted mutagenesis and single-stranded oligodeoxynucleotide-mediated precise knock-in in the industrial microalga Euglena gracilis using Cas9 ribonucleoproteins. Plant Biotechnol. J. 2019, 17, 2032. [Google Scholar] [CrossRef]
- Nomura, T.; Yoshikawa, M.; Suzuki, K.; Mochida, K. Highly efficient CRISPR-associated protein 9 ribonucleoprotein-based genome editing in Euglena gracilis. STAR Protoc. 2020, 1, 100023. [Google Scholar] [CrossRef]
- Chopra, T.S.; Dube, J.N. Changes of N content of a rice soil inoculated with Tolypothrix tenuis. Plant Soil 1971, 35, 453–462. [Google Scholar] [CrossRef]
- Lode, A.; Krujatz, F.; Brüggemeier, S.; Quade, M.; Schütz, K.; Knaack, S.; Weber, J.; Bley, T.; Gelinsky, M. Green bioprinting: Fabrication of photosynthetic algae—Laden hydrogel scaffolds for biotechnological and medical applications. Eng. Life Sci. 2015, 15, 177–183. [Google Scholar] [CrossRef]
- Nakashima, A.; Sasaki, K.; Sasaki, D.; Yasuda, K.; Suzuki, K.; Kondo, A. The alga Euglena gracilis stimulates Faecalibacterium in the gut and contributes to increased defecation. Sci. Rep. 2021, 11, 1074. [Google Scholar] [CrossRef]
- Sakanoi, Y.; E, S.; Yamamoto, K.; Ota, T.; Seki, K.; Imai, M.; Ota, R.; Asayama, Y.; Nakashima, A.; Suzuki, K. Simultaneous intake of Euglena gracilis and vegetables synergistically exerts an anti-inflammatory effect and attenuates visceral fat accumulation by affecting gut microbiota in mice. Nutrients 2018, 10, 1417. [Google Scholar] [CrossRef]
- Okouchi, R.; E, S.; Yamamoto, K.; Ota, T.; Seki, K.; Imai, M.; Ota, R.; Asayama, Y.; Nakashima, A.; Suzuki, K. Simultaneous intake of Euglena gracilis and vegetables exerts synergistic anti-obesity and anti-inflammatory effects by modulating the gut microbiota in diet-induced obese mice. Nutrients 2019, 11, 204. [Google Scholar] [CrossRef]
- Ishiguro, S.; Upreti, D.; Robben, N.; Burghart, R.; Loyd, M.; Ogun, D.; Le, T.; Delzeit, J.; Nakashima, A.; Thakkar, R. Water extract from Euglena gracilis prevents lung carcinoma growth in mice by attenuation of the myeloid-derived cell population. Biomed. Pharm. 2020, 127, 110166. [Google Scholar] [CrossRef]
- Upreti, D.; Ishiguro, S.; Robben, N.; Nakashima, A.; Suzuki, K.; Comer, J.; Tamura, M. Oral administration of water extract from Euglena gracilis alters the intestinal microbiota and prevents lung carcinoma hrowth in mice. Nutrients 2022, 14, 678. [Google Scholar] [CrossRef]
- Kusmic, C.; Barsanti, L.; Di Lascio, N.; Faita, F.; Evangelista, V.; Gualtieri, P. Anti-fibrotic effect of paramylon nanofibers from the WZSL mutant of Euglena gracilis on liver damage induced by CCl4 in mice. J. Funct. Foods 2018, 46, 538–545. [Google Scholar] [CrossRef]
- Nagayama, Y.; Isoo, N.; Nakashima, A.; Suzuki, K.; Yamano, M.; Nariyama, T.; Yagame, M.; Matsui, K. Renoprotective effects of paramylon, a β-1, 3-D-Glucan isolated from Euglena gracilis Z in a rodent model of chronic kidney disease. PLoS ONE 2020, 15, e0237086. [Google Scholar] [CrossRef]
- Nakashima, A.; Yasuda, K.; Murata, A.; Suzuki, K.; Miura, N. Effects of Euglena gracilis intake on mood and autonomic activity under mental workload, and subjective sleep quality: A randomized, double-blind, placebo-controlled trial. Nutrients 2020, 12, 3243. [Google Scholar] [CrossRef]
- Yasuda, K.; Ogushi, M.; Nakashima, A.; Nakano, Y.; Suzuki, K. Accelerated wound healing on the skin using a film dressing with β-glucan paramylon. In Vivo 2018, 32, 799–805. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Häder, D.-P.; Hemmersbach, R. Euglena, a Gravitactic Flagellate of Multiple Usages. Life 2022, 12, 1522. https://doi.org/10.3390/life12101522
Häder D-P, Hemmersbach R. Euglena, a Gravitactic Flagellate of Multiple Usages. Life. 2022; 12(10):1522. https://doi.org/10.3390/life12101522
Chicago/Turabian StyleHäder, Donat-P., and Ruth Hemmersbach. 2022. "Euglena, a Gravitactic Flagellate of Multiple Usages" Life 12, no. 10: 1522. https://doi.org/10.3390/life12101522
APA StyleHäder, D. -P., & Hemmersbach, R. (2022). Euglena, a Gravitactic Flagellate of Multiple Usages. Life, 12(10), 1522. https://doi.org/10.3390/life12101522