
New Fossil Evidence Suggests That Angiosperms Flourished in the Middle Jurassic



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
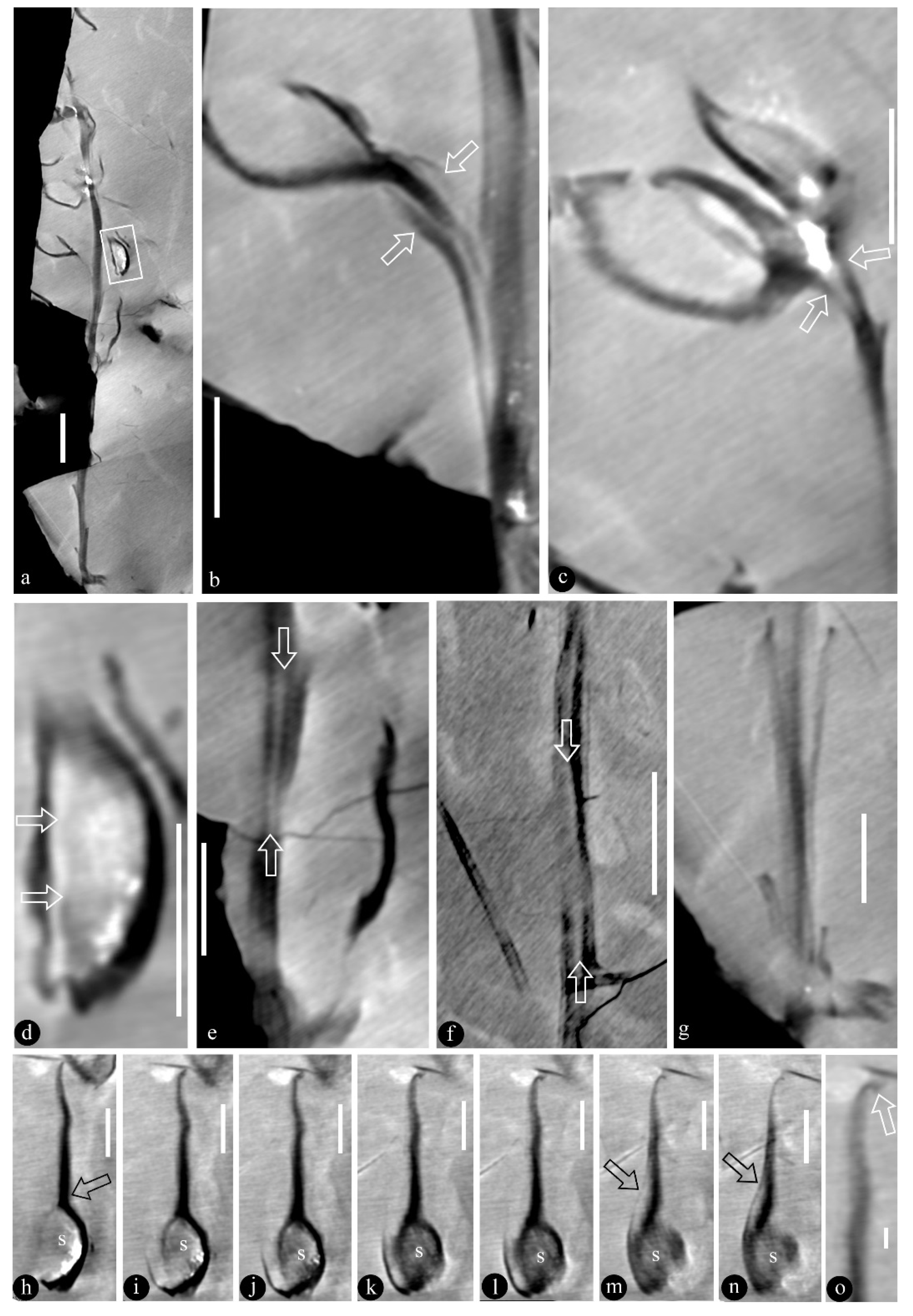
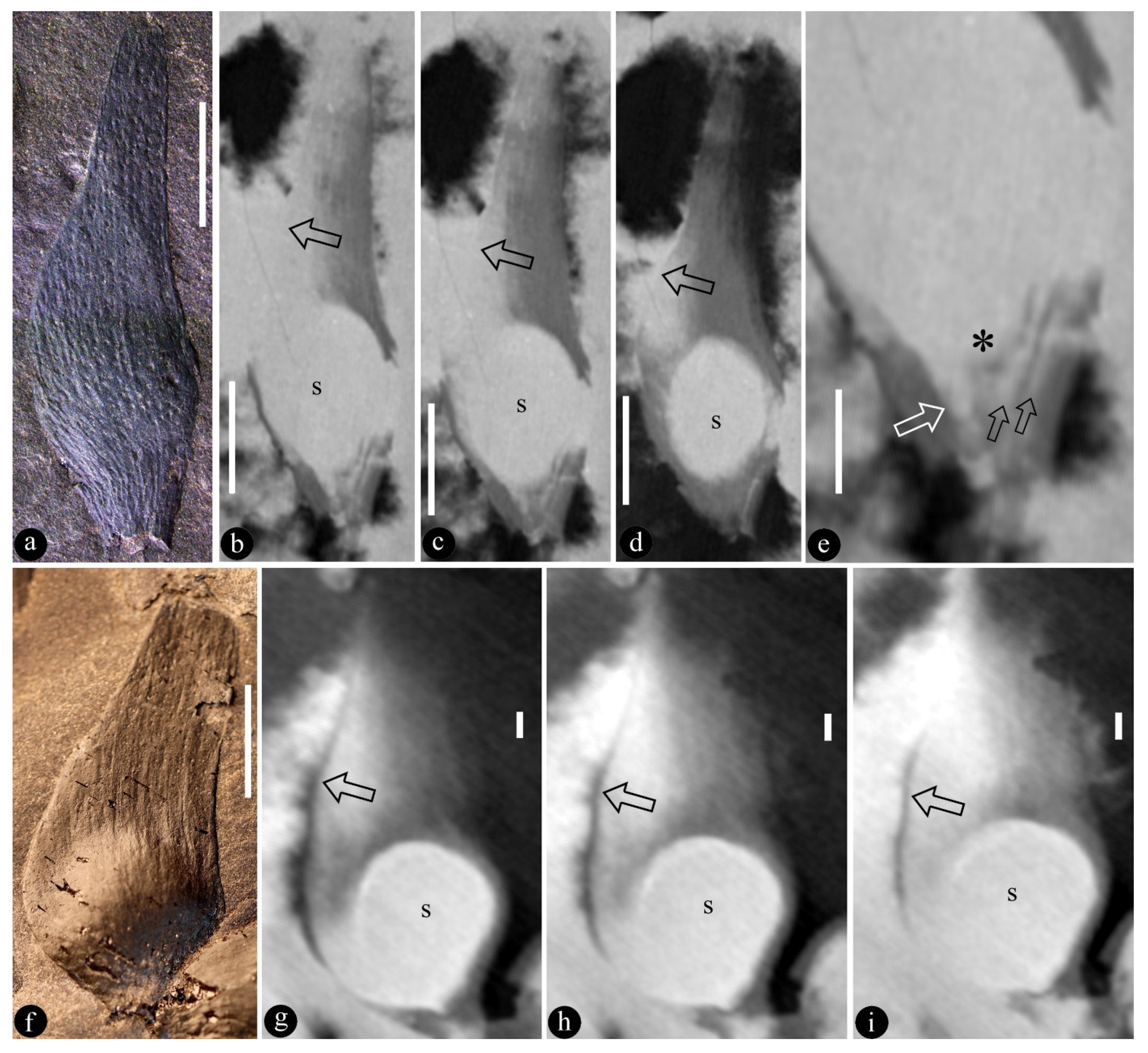
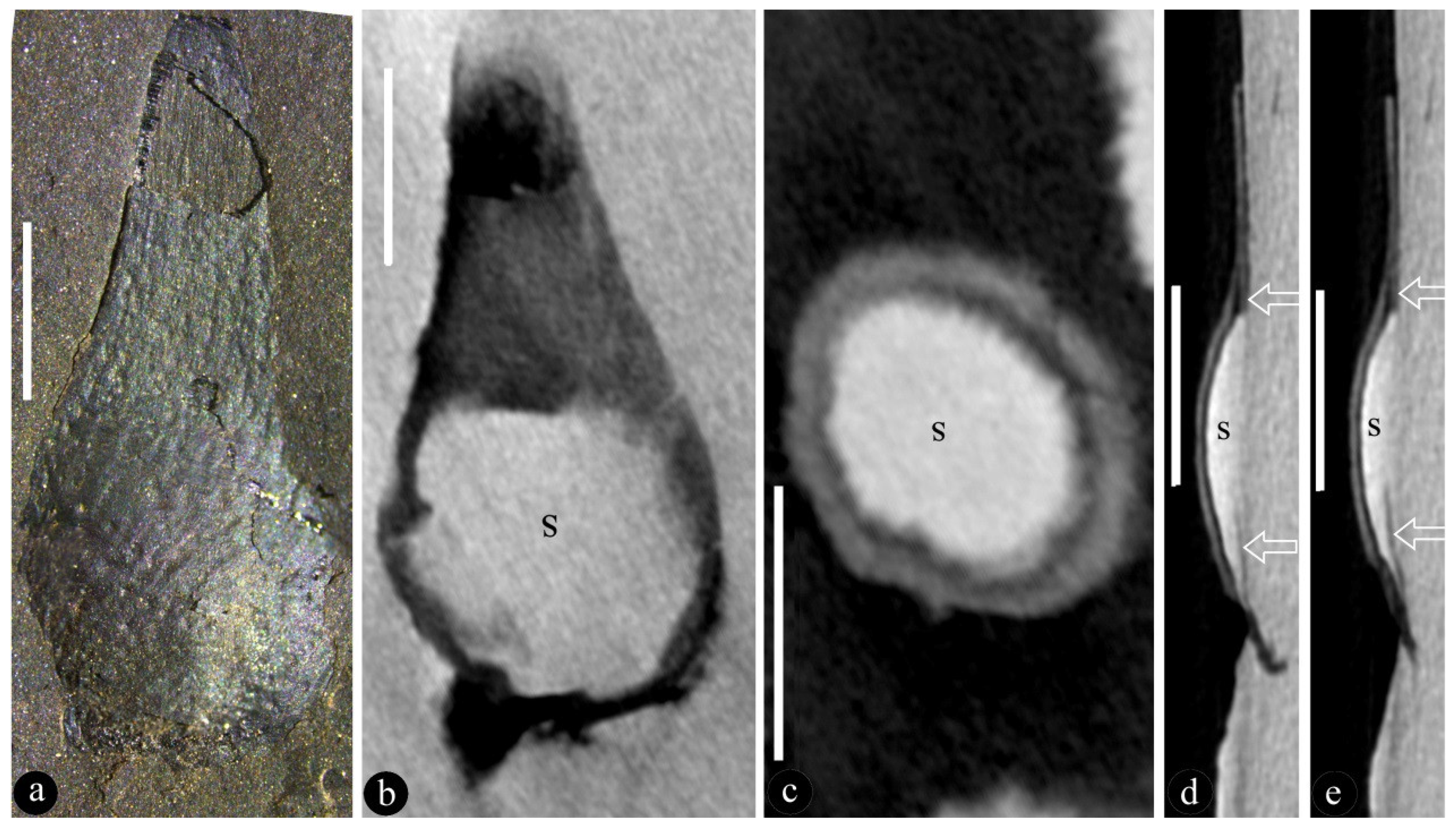
3. Systematic Paleobotany
- Genus Qingganninginfructus Wang and Sun, gen. nov.
- Qingganninginfructus formosa Wang and Sun, gen. et sp. nov.
- Qingganninginfructus sp.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, G.; Dilcher, D.L.; Zheng, S.; Zhou, Z. In search of the first flower: A Jurassic angiosperm, Archaefructus, from Northeast China. Science 1998, 282, 1692–1695. [Google Scholar] [CrossRef]
- Friis, E.M.; Pedersen, K.R.; Crane, P.R. Fossil evidence of water lilies (Nymphaeales) in the Early Cretaceous. Nature 2001, 410, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zheng, X.-T. Reconsiderations on two characters of early angiosperm Archaefructus. Palaeoworld 2012, 21, 193–201. [Google Scholar] [CrossRef]
- Ji, Q.; Li, H.; Bowe, M.; Liu, Y.; Taylor, D.W. Early Cretaceous Archaefructus eoflora sp. nov. with bisexual flowers from Beipiao, Western Liaoning, China. Acta Geol. Sin (Engl. Ed.) 2004, 78, 883–896. [Google Scholar]
- Gandolfo, M.A.; Nixon, K.C.; Crepet, W.L. Cretaceous flowers of Nymphaeaceae and implications for complex insect entrapment pollination mechanisms in early angiosperms. Proc. Natl. Acad. Sci. USA 2004, 101, 8056–8060. [Google Scholar] [CrossRef] [Green Version]
- Cornet, B. The leaf venation and reproductive structures of a late Triassic angiosperm, Sanmiguelia lewisii. Evol. Theory 1986, 7, 231–308. [Google Scholar]
- Cornet, B. Late Triassic angiosperm-like pollen from the Richmond rift basin of Virginia, USA. Palaeontogr. B 1989, 213, 37–87. [Google Scholar]
- Cornet, B. The reproductive morphology and biology of Sanmiguelia lewisii, and its bearing on angiosperm evolution in the late Triassic. Evol. Trends Plants 1989, 3, 25–51. [Google Scholar]
- Cornet, B. Dicot-like leaf and flowers from the Late Triassic tropical Newark Supergroup rift zone, U.S.A. Mod. Biol. 1993, 19, 81–99. [Google Scholar]
- Cornet, B.; Habib, D. Angiosperm-like pollen from the ammonite-dated Oxfordian (Upper Jurassic) of France. Rev. Palaeobot. Palynol. 1992, 71, 269–294. [Google Scholar] [CrossRef]
- Hochuli, P.A.; Feist-Burkhardt, S. A boreal early cradle of angiosperms? Angiosperm-like pollen from the Middle Triassic of the Barents Sea (Norway). J. Micropalaeontol. 2004, 23, 97–104. [Google Scholar] [CrossRef]
- Hochuli, P.A.; Feist-Burkhardt, S. Angiosperm-like pollen and Afropollis from the Middle Triassic (Anisian) of the Germanic Basin (Northern Switzerland). Front. Plant Sci. 2013, 4, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Q.; Diez, J.B.; Pole, M.; Garcia-Avila, M.; Liu, Z.-J.; Chu, H.; Hou, Y.; Yin, P.; Zhang, G.-Q.; Du, K.; et al. An unexpected noncarpellate epigynous flower from the Jurassic of China. eLife 2018, 7, e38827. [Google Scholar] [CrossRef]
- Fu, Q.; Diez, J.B.; Pole, M.; García-Ávila, M.; Wang, X. Nanjinganthus is an angiosperm, isn’t it? China Geol. 2020, 3, 359–361. [Google Scholar] [CrossRef]
- Fu, Q.; Hou, Y.; Yin, P.; Diez, J.B.; Pole, M.; García-Ávila, M.; Wang, X. Micro-CT results exhibit ovules enclosed in the ovaries of Nanjinganthus. Sci. Rep. 2023, 13, 426. [Google Scholar] [CrossRef]
- Liu, Z.-J.; Hou, Y.-M.; Wang, X. Zhangwuia: An enigmatic organ with bennettitalean appearance and enclosed ovules. Earth Environ. Sci. Trans. R. Soc. Edinb. 2019, 108, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.-J.; Wang, X. A perfect flower from the Jurassic of China. Hist. Biol. 2016, 28, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.-J.; Wang, X. Yuhania: A unique angiosperm from the Middle Jurassic of Inner Mongolia, China. Hist. Biol. 2017, 29, 431–441. [Google Scholar] [CrossRef] [Green Version]
- Wang, X. Schmeissneria: An angiosperm from the Early Jurassic. J. Syst. Evol. 2010, 48, 326–335. [Google Scholar] [CrossRef]
- Wang, X.; Duan, S.; Geng, B.; Cui, J.; Yang, Y. Schmeissneria: A missing link to angiosperms? BMC Evol. Biol. 2007, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Han, G. The earliest ascidiate carpel and its implications for angiosperm evolution. Acta Geol. Sin. 2011, 85, 998–1002. (In English) [Google Scholar] [CrossRef]
- Wang, X.; Wang, S. Xingxueanthus: An enigmatic Jurassic seed plant and its implications for the origin of angiospermy. Acta Geol. Sin. (Engl. Ed.) 2010, 84, 47–55. [Google Scholar] [CrossRef]
- Han, G.; Liu, Z.; Wang, X. A Dichocarpum-like angiosperm from the Early Cretaceous of China. Acta Geol. Sin. (Engl. Ed.) 2017, 90, 1–8. [Google Scholar]
- Han, G.; Liu, Z.-J.; Liu, X.; Mao, L.; Jacques, F.M.B.; Wang, X. A whole plant herbaceous angiosperm from the Middle Jurassic of China. Acta Geol. Sin. (Engl. Ed.) 2016, 90, 19–29. [Google Scholar]
- Scott, R.A.; Barghoorn, E.S.; Leopold, E.B. How old are the angiosperms? Am. J. Sci. 1960, 258-A, 284–299. [Google Scholar]
- Herendeen, P.S.; Friis, E.M.; Pedersen, K.R.; Crane, P.R. Palaeobotanical redux: Revisiting the age of the angiosperms. Nat. Plants 2017, 3, 17015. [Google Scholar] [CrossRef] [Green Version]
- Wang, X. Criterion is a touchstone in study of early angiosperms. Open J. Plant Sci. 2021, 6, 91–93. [Google Scholar]
- Wang, X. The Dawn Angiosperms; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Wang, X. The Dawn Angiosperms, 2nd ed.; Springer: Cham, Switzerland, 2018; p. 407. [Google Scholar]
- Sokoloff, D.D.; Remizowa, M.V.; El, E.S.; Rudall, P.J.; Bateman, R.M. Supposed Jurassic angiosperms lack pentamery, an important angiosperm-specific feature. New Phytol. 2020, 228, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Coiro, M.; Doyle, J.A.; Hilton, J. How deep is the conflict between molecular and fossil evidence on the age of angiosperms? New Phytol. 2019, 223, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Bateman, R.M. Hunting the snark: The flawed search for mythical Jurassic angiosperms. J. Exp. Bot. 2020, 71, 22–35. [Google Scholar] [CrossRef]
- Xu, X.; Feng, M. Micro-CT Technology and Its Applications in Plants; Science Press: Beijing, China, 2019; p. 97. [Google Scholar]
- Zhang, H.; Li, H.; Xiong, C.; Zhang, H.; Wang, Y.; He, Z.; Lin, G.; Sun, B. Jurassic Coaliferous Strata and Coal Accumulation in Northwest China; Geological Publishing House: Beijing, China, 1998; p. 317. [Google Scholar]
- Sun, B.-N. A preliminary study of Middle Jurassic fossil plants from Yaojie Coal-field of Lanzhou. J. Lanzhou Univ. 1986, 22, 113–118. [Google Scholar]
- Xin, C.; Zhang, Y.; Ji, J.; Wang, L. Characteristics of the Middle Jurassic Flora at Yaojie, Gansu Province, and its paleoecological significance. Acta Geol. Sin. 2019, 92, 1985–2001. [Google Scholar]
- Yan, D.; Sun, B.; Xie, S.; Li, X.; Wen, W. Response to paleoatmospheric CO2 concentration of Solenites vimineus (Phillips) Harris (Ginkgophyta) from the Middle Jurassic of the Yaojie Basin, Gansu Province, China. Sci. China Ser. D Earth Sci. 2009, 52, 2029–2039. [Google Scholar] [CrossRef]
- Sun, B.-N.; Shen, G.-L. Rhizomopteris yaojieensis sp. nov. a new species of the genus Rhizomopteris. J. Lanzhou Univ. 1986, 22, 127–130. [Google Scholar]
- Sun, B.-N.; Shen, G.-L. A new species of genus Nilssoniopteris. Acta Palaeontol. Sin. 1988, 27, 561–564. [Google Scholar]
- Sun, B.-N.; Shi, Y.-J.; Zhang, C.-J.; Wang, Y.-P. A Preliminary Study of Middle Jurassic Fossil Plants from Yaojie Coal-Field of Lanzhou; Science Press: Beijing, China, 2005. [Google Scholar]
- Sun, B.-N.; Xie, S.-P.; Yan, D.-F.; Cong, P.-Y. Fossil plant evidence for Early and Middle Jurassic paleoenvironmental changes in Lanzhou area, Northwest China. Palaeoworld 2008, 17, 215–221. [Google Scholar] [CrossRef]
- Schweitzer, H.-J.; Kirchner, M. Die Rhaeto-Jurassischen Floren des Iran und Afghanistans: 9. Coniferophyta. Palaeontogr. B 1996, 238, 77–139. [Google Scholar]
- Santos, A.A.; Wang, X. Pre-Carpels from the Middle Triassic of Spain. Plants 2022, 11, 2833. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Manchester, S.R. Red and silver maples in the Neogene of Western North America: Fossil leaves and samaras of Acer section Rubra. Int. J. Plant Sci. 2020, 181, 542–556. [Google Scholar] [CrossRef]
- Gabrielyan, I.; Kovar-Eder, J. The genus Acer from the lower/middle Pleistocene Sisian Formation, Syunik region, South Armenia. Rev. Palaeobot. Palynol. 2011, 165, 111–134. [Google Scholar] [CrossRef]
- Zhang, X. Floral ontogeny of Illicium lanceolatum (Schisandraceae) and its implications on carpel homology. Phytotaxa 2019, 416, 200–210. [Google Scholar] [CrossRef]
- Ren, Y.; Li, H.-F.; Zhao, L.; Endress, P.K. Floral Morphogenesis in Euptelea (Eupteleaceae, Ranunculales). Ann. Bot. 2007, 100, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Friis, E.M.; Pedersen, K.R.; von Balthazar, M.; Grimm, G.W.; Crane, P.R. Monetianthus mirus gen. et sp. nov. a nymphaealean flower from the Early Cretaceous of Portugal. Int. J. Plant Sci. 2009, 170, 1086–1101. [Google Scholar] [CrossRef]
- Friis, E.M.; Crane, P.R.; Pedersen, K.R. Hedyosmum-like fossils in the Early Cretaceous diversification of angiosperms. Int. J. Plant Sci. 2019, 180, 232–239. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, P.B.; Takaso, T. Seed cone structure in conifers in relation to development and pollination: A biological approach. Can. J. Bot. 2002, 80, 1250–1273. [Google Scholar] [CrossRef]
- Han, G.; Fu, X.; Liu, Z.-J.; Wang, X. A new angiosperm genus from the Lower Cretaceous Yixian Formation, Western Liaoning, China. Acta Geol. Sin. (Engl. Ed.) 2013, 87, 916–925. [Google Scholar]
- Han, G.; Wang, X. A new infructescence of angiosperms from the Early Cretaceous of China. Acta Geol. Sin. (Engl. Ed.) 2020, 94, 1711–1713. [Google Scholar] [CrossRef]
- Liu, Z.-J.; Wang, X. A novel angiosperm from the Early Cretaceous and its implications on carpel-deriving. Acta Geol. Sin. (Engl. Ed.) 2018, 92, 1293–1298. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Duan, S.Y.; Cui, J.Z. Several species of Schizolepis and their significance on the evolution of conifers. Taiwania 1997, 42, 73–85. [Google Scholar]
- Prasad, V.; Strömberg, C.A.E.; Leaché, A.D.; Samant, B.; Patnaik, R.; Tang, L.; Mohabey, D.M.; Ge, S.; Sahni, A. Late Cretaceous origin of the rice tribe provides evidence for early diversification in Poaceae. Nat. Commun. 2011, 2, 480. [Google Scholar] [CrossRef] [Green Version]
- Li, H.-T.; Yi, T.-S.; Gao, L.-M.; Ma, P.-F.; Zhang, T.; Yang, J.-B.; Gitzendanner, M.A.; Fritsch, P.W.; Cai, J.; Luo, Y.; et al. Origin of angiosperms and the puzzle of the Jurassic gap. Nat. Plants 2019, 5, 461–470. [Google Scholar] [CrossRef]
- Wu, Y.; You, H.-L.; Li, X.-Q. Dinosaur-associated Poaceae epidermis and phytoliths from the Early Cretaceous of China. Natl. Sci. Rev. 2018, 5, 721–727. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, L.; Zhao, Y.; Zhao, M.; Sun, J.; Sun, B.; Wang, X. New Fossil Evidence Suggests That Angiosperms Flourished in the Middle Jurassic. Life 2023, 13, 819. https://doi.org/10.3390/life13030819
Han L, Zhao Y, Zhao M, Sun J, Sun B, Wang X. New Fossil Evidence Suggests That Angiosperms Flourished in the Middle Jurassic. Life. 2023; 13(3):819. https://doi.org/10.3390/life13030819
Chicago/Turabian StyleHan, Lei, Ya Zhao, Ming Zhao, Jie Sun, Bainian Sun, and Xin Wang. 2023. "New Fossil Evidence Suggests That Angiosperms Flourished in the Middle Jurassic" Life 13, no. 3: 819. https://doi.org/10.3390/life13030819
APA StyleHan, L., Zhao, Y., Zhao, M., Sun, J., Sun, B., & Wang, X. (2023). New Fossil Evidence Suggests That Angiosperms Flourished in the Middle Jurassic. Life, 13(3), 819. https://doi.org/10.3390/life13030819