


Apocynin Ameliorates Monosodium Glutamate Induced Testis Damage by Impaired Blood-Testis Barrier and Oxidative Stress Parameters
 , , and
, , and 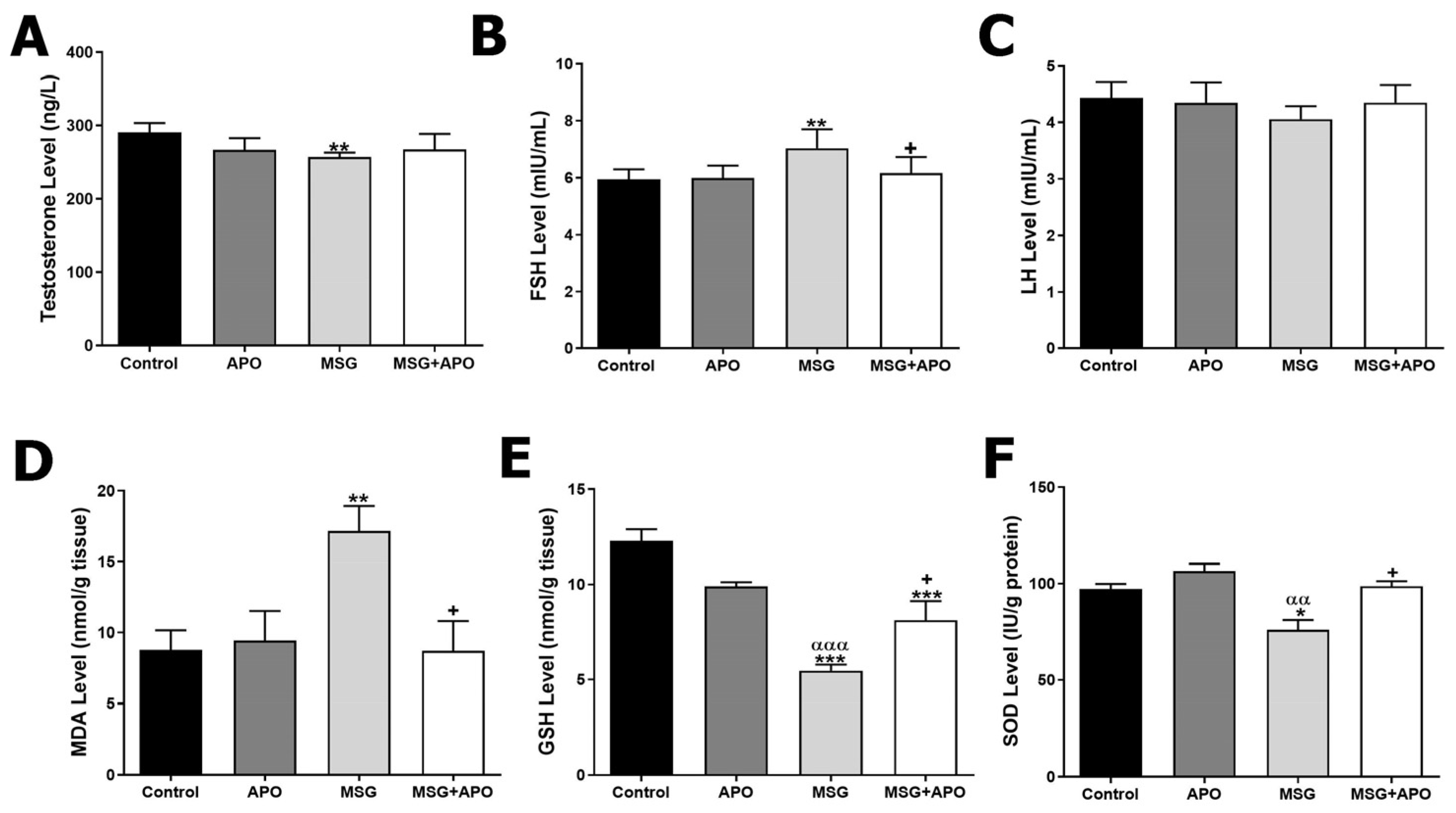{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Measurement of Serum Testosterone, FSH, and LH Concentrations
2.3. Measurement of Testicular MDA, GSH, and SOD Levels
2.4. Sperm Count, Motility, and Morphology
2.5. Tissue Processing for Light Microscopy
2.6. Terminal Deoxynucleotidyl Transferase dUTP Nick End Labelling (TUNEL) Immunochemistry
2.7. Proliferating Cell Nuclear Antigen (PCNA) Immunohistochemistry
2.8. Immunohistochemistry of ZO-1 and Occludin
2.9. ZO-1, Occludin and NOX-2 Immunofluorescence Analysis
2.10. Tissue Processing for Transmission Electron Microscopy
2.11. Statistical Analysis
3. Results
3.1. Serum Testosterone, FSH, and LH Levels
3.2. Testicular MDA, GSH, and SOD Levels
3.3. Sperm Count, Motility, and Morphology
3.4. Testicular Weight/Body Weight Ratio
3.5. Histopathological Results
3.6. Results for PCNA
3.7. Results for TUNEL Immunocytochemistry
3.8. Results for NOX-2 Immunofluorescence
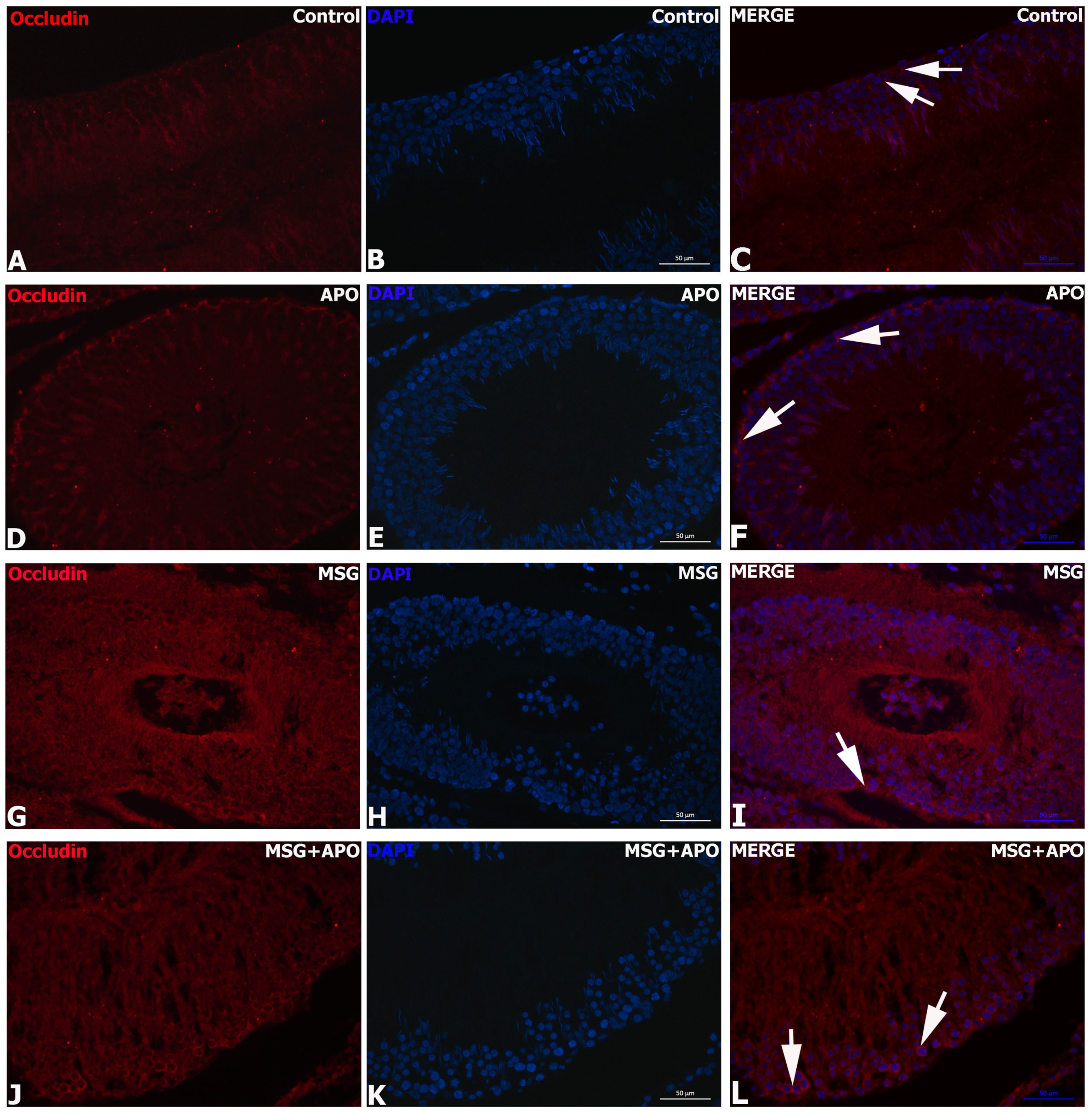
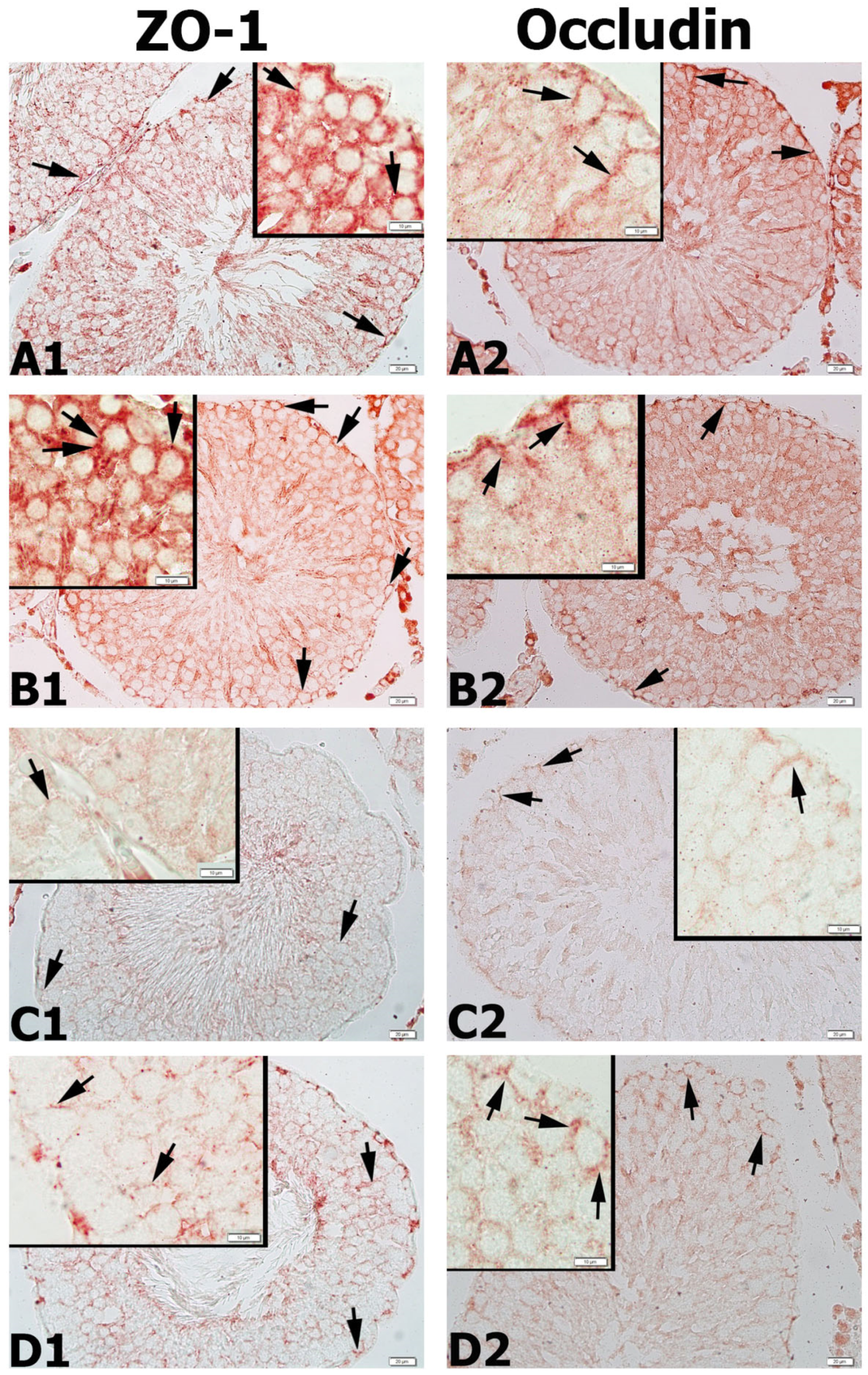
3.9. Results for ZO-1 and Occludin Immunohistochemistry
3.10. Transmission Electron Microscopical Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zanfirescu, A.; Ungurianu, A.; Tsatsakis, A.M.; Nițulescu, G.M.; Kouretas, D.; Veskoukis, A.; Tsoukalas, D.; Engin, A.B.; Aschner, M.; Margină, D. A review of the alleged health hazards of monosodium glutamate. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1111–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyreuther, K.; Biesalski, H.K.; Fernstrom, J.D.; Grimm, P.; Hammes, W.P.; Heinemann, U.; Kempski, O.; Stehle, P.; Steinhart, H.; Walker, R. Consensus meeting: Monosodium glutamate—An update. Eur. J. Clin. Nutr. 2007, 61, 304–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayode, O.T.; Rotimi, D.E.; Kayode, A.A.A.; Olaolu, T.D.; Adeyemi, O.S. Monosodium Glutamate (MSG)-Induced Male Reproductive Dysfunction: A Mini Review. Toxics 2020, 8, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jubaidi, F.F.; Mathialagan, R.D.; Noor, M.M.; Taib, I.S.; Budin, S.B. Monosodium glutamate daily oral supplementation: Study of its effects on male reproductive system on rat model. Syst. Biol. Reprod. Med. 2019, 65, 194–204. [Google Scholar] [CrossRef]
- Shin, J.-W.; Seol, I.-C.; Son, C.-G. Interpretation of animal dose and human equivalent dose for drug development. J. Korean Med. 2010, 31, 1–7. [Google Scholar]
- Igwebuike, U.M.; Nwankwo, I.A.; Ochiogu, I.S. Effects of oral administration of monosodium glutamate (MSG) on serum testosterone levels and muscle mass development in male rats. Anim. Res. Int. 2017, 7, 2. [Google Scholar]
- Abu Hanipah, E.N.; Yahya, N.J.; AJIK, E.; Yusoff, N.A.; Taib, I.S. Monosodium Glutamate Induced Oxidative Stress in Accessory Reproductive Organs of Male Sprague-Dawley Rats. Malays. J. Health Sci./J. Sains Kesihat. Malays. 2018, 16, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Sezer, K.; Keskin, M. Serbest oksijen radikallerinin hastalıkların patogenezisindeki rolü. FÜ Sağ. Bil. Vet. Derg. 2014, 28, 49–56. [Google Scholar]
- Hamada, A.; Sharma, R.; du Plessis, S.S.; Willard, B.; Yadav, S.P.; Sabanegh, E.; Agarwal, A. Two-dimensional differential in-gel electrophoresis-based proteomics of male gametes in relation to oxidative stress. Fertil. Steril. 2013, 99, 1216–1226.e2. [Google Scholar] [CrossRef]
- Büyükuslu, N.; Yiğitbaşı, T. Reaktif oksijen türleri ve obezitede oksidatif stres. Clin. Exp. Health Sci. 2015, 5, 197–203. [Google Scholar]
- Khosrowbeygi, A.; Zarghami, N. Levels of oxidative stress biomarkers in seminal plasma and their relationship with seminal parameters. BMC Clin. Pathol. 2007, 7, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, A.; Durairajanayagam, D.; Halabi, J.; Peng, J.; Vazquez-Levin, M. Proteomics, oxidative stress and male infertility. Reprod. Biomed. Online 2014, 29, 32–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velando, A.; Torres, R.; Alonso-Alvarez, C. Avoiding bad genes: Oxidatively damaged DNA in germ line and mate choice. Bioessays 2008, 30, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Virdis, A.; Gesi, M.; Taddei, S. Impact of apocynin on vascular disease in hypertension. Vascul. Pharmacol. 2016, 87, 1–5. [Google Scholar] [CrossRef]
- Barbieri, S.S.; Cavalca, V.; Eligini, S.; Brambilla, M.; Caiani, A.; Tremoli, E.; Colli, S. Apocynin prevents cyclooxygenase 2 expression in human monocytes through NADPH oxidase and glutathione redox-dependent mechanisms. Free Radic. Biol. Med. 2004, 37, 156–165. [Google Scholar] [CrossRef]
- Peters, E.A.; Hiltermann, J.T.; Stolk, J. Effect of apocynin on ozone-induced airway hyperresponsiveness to methacholine in asthmatics. Free Radic. Biol. Med. 2001, 31, 1442–1447. [Google Scholar] [CrossRef]
- Smit, H.F.; Kroes, B.H.; van den Berg, A.J.; van der Wal, D.; van den Worm, E.; Beukelman, C.J.; van Dijk, H.; Labadie, R.P. Immunomodulatory and anti-inflammatory activity of Picrorhiza scrophulariiflora. J. Ethnopharmacol. 2000, 73, 101–109. [Google Scholar] [CrossRef]
- Johnson, D.K.; Schillinger, K.J.; Kwait, D.M.; Hughes, C.V.; McNamara, E.J.; Ishmael, F.; O’Donnell, R.W.; Chang, M.M.; Hogg, M.G.; Dordick, J.S.; et al. Inhibition of NADPH oxidase activation in endothelial cells by ortho-methoxy-substituted catechols. Endothelium 2002, 9, 191–203. [Google Scholar] [CrossRef]
- Stolk, J.; Hiltermann, T.J.; Dijkman, J.H.; Verhoeven, A.J. Characteristics of the inhibition of NADPH oxidase activation in neutrophils by apocynin, a methoxy-substituted catechol. Am. J. Respir. Cell Mol. Biol. 1994, 11, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Contuk, G.; Orun, O.; Demiralp-Ekşioğlu, E.; Ercan, F. Morphological alterations and distribution of occludin in rat testes after bilateral vasectomy. Acta Histochem. 2012, 114, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Pawlina, W.; Ross, M.H. Histology: A Text and Atlas: With Correlated Cell and Molecular Biology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2018. [Google Scholar]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.F.; Feng, Q.; Ge, Z.Y.; Guo, Y.; Zhou, F.; Zhang, K.S.; Wang, X.W.; Lu, W.H.; Liang, X.W.; Gu, Y.Q. Obesity impairs male fertility through long-term effects on spermatogenesis. BMC Urol. 2018, 18, 42. [Google Scholar] [CrossRef] [Green Version]
- Connell, B.J.; Saleh, M.C.; Khan, B.V.; Saleh, T.M. Apocynin may limit total cell death following cerebral ischemia and reperfusion by enhancing apoptosis. Food Chem. Toxicol. 2011, 49, 3063–3069. [Google Scholar] [CrossRef] [PubMed]
- Stefanska, J.; Pawliczak, R. Apocynin: Molecular aptitudes. Mediat. Inflamm. 2008, 2008, 106507. [Google Scholar] [CrossRef] [Green Version]
- Köroğlu, K.M.; Çevik, Ö.; Şener, G.; Ercan, F. Apocynin alleviates cisplatin-induced testicular cytotoxicity by regulating oxidative stress and apoptosis in rats. Andrologia 2019, 51, e13227. [Google Scholar] [CrossRef]
- Beutler, E. Red Cell Metabolism: A Manual of Biochemical Methods, 2nd ed.; National Library of Medicine: Bethesda, MD, USA, 1984. [Google Scholar]
- Acikel Elmas, M.; Cakıcı, S.E.; Dur, I.R.; Kozluca, I.; Arınc, M.; Binbuga, B.; Bingol Ozakpınar, O.; Kolgazi, M.; Sener, G.; Ercan, F. Protective effects of exercise on heart and aorta in high-fat diet-induced obese rats. Tissue Cell 2019, 57, 57–65. [Google Scholar] [CrossRef]
- Gozen, A.; Demiryurek, S.; Taskin, A.; Ciralik, H.; Bilinc, H.; Kara, S.; Aydin, A.; Aksoy, N.; Ceylan, H. Protective activity of ischemic preconditioning on rat testicular ischemia: Effects of Y-27632 and 5-hydroxydecanoic acid. J. Pediatr. Surg. 2013, 48, 1565–1572. [Google Scholar] [CrossRef]
- Johnsen, S.G. Testicular biopsy score count—A method for registration of spermatogenesis in human testes: Normal values and results in 335 hypogonadal males. Hormones 1970, 1, 2–25. [Google Scholar] [CrossRef]
- Elmas, M.A.; Ozakpinar, O.B.; Kolgazi, M.; Sener, G.; Arbak, S.; Ercan, F. Exercise improves testicular morphology and oxidative stress parameters in rats with testicular damage induced by a high-fat diet. Andrologia 2022, 54, e14600. [Google Scholar] [CrossRef]
- Abd-Elkareem, M.; Abd El-Rahman, M.A.M.; Khalil, N.S.A.; Amer, A.S. Antioxidant and cytoprotective effects of Nigella sativa L. seeds on the testis of monosodium glutamate challenged rats. Sci. Rep. 2021, 11, 13519. [Google Scholar] [CrossRef] [PubMed]
- Hamza, R.Z.; Al-Harbi, M.S. Monosodium glutamate induced testicular toxicity and the possible ameliorative role of vitamin E or selenium in male rats. Toxicol. Rep. 2014, 1, 1037–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosnan, J.T.; Drewnowski, A.; Friedman, M.I. Is there a relationship between dietary MSG and obesity in animals or humans? Amino Acids 2014, 46, 2075–2087. [Google Scholar] [CrossRef] [PubMed]
- Oyola, M.G.; Handa, R.J. Hypothalamic-pituitary-adrenal and hypothalamic-pituitary-gonadal axes: Sex differences in regulation of stress responsivity. Stress 2017, 20, 476–494. [Google Scholar] [CrossRef]
- Corradi, P.; Corradi, R.; Greene, L. Physiology of the Hypothalamic Pituitary Gonadal Axis in the Male. Urol. Clin. N. Am. 2016, 43, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Salonia, A.; Rastrelli, G.; Hackett, G.; Seminara, S.B.; Huhtaniemi, I.T.; Rey, R.A.; Hellstrom, W.J.G.; Palmert, M.R.; Corona, G.; Dohle, G.R.; et al. Paediatric and adult-onset male hypogonadism. Nat. Rev. Dis. Prim. 2019, 5, 38. [Google Scholar] [CrossRef]
- Zhao, W.; Jing, J.; Shao, Y.; Zeng, R.; Wang, C.; Yao, B.; Hang, D. Circulating sex hormone levels in relation to male sperm quality. BMC Urol. 2020, 20, 101. [Google Scholar] [CrossRef]
- Keskin, M.Z.; Budak, S.; Zeyrek, T.; Çelik, O.; Mertoglu, O.; Yoldas, M.; Ilbey, Y. The relationship between serum hormone levels (follicle-stimulating hormone, luteinizing hormone, total testosterone) and semen parameters. Arch. Ital. Urol. Androl. 2015, 87, 194–197. [Google Scholar] [CrossRef] [Green Version]
- Hamza, R.Z.; Al-Baqami, N.M. Testicular protective effects of ellagic acid on monosodium glutamate-induced testicular structural alterations in male rats. Ultrastruct. Pathol. 2019, 43, 170–183. [Google Scholar] [CrossRef]
- Khan, S.; Sheikh, M.A.; Danyal, A.; Ali, I.; Tahir, F.; Bashir, M.; Rahat, N.; Asif, A.H. Does LH/Testo and Testo/LH ratio affect male fertility? Med. Forum Mon. 2006, 17, 5–9. [Google Scholar]
- Miao, L.; St Clair, D.K. Regulation of superoxide dismutase genes: Implications in disease. Free Radic. Biol. Med. 2009, 47, 344–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oestreicher, J.; Morgan, B. Glutathione: Subcellular distribution and membrane transport (1). Biochem. Cell Biol. Biochim. Biol. Cell. 2019, 97, 270–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandes, R.P.; Weissmann, N.; Schröder, K. Nox family NADPH oxidases: Molecular mechanisms of activation. Free Radic. Biol. Med. 2014, 76, 208–226. [Google Scholar] [CrossRef] [PubMed]
- Hamza, R.Z.; Diab, A.E.-A.A. Testicular protective and antioxidant effects of selenium nanoparticles on Monosodium glutamate-induced testicular structure alterations in male mice. Toxicol. Rep. 2020, 7, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.-L.; Dai, D.-Z.; Zhang, C.; Dai, Y. Apocynin and raisanberine alleviate intermittent hypoxia induced abnormal StAR and 3β-HSD and low testosterone by suppressing endoplasmic reticulum stress and activated p66Shc in rat testes. Reprod. Toxicol. 2013, 36, 60–70. [Google Scholar] [CrossRef]
- Eweka, A.; Igbigbi, P.; Ucheya, R. Histochemical studies of the effects of monosodium glutamate on the liver of adult wistar rats. Ann. Med. Health Sci. Res. 2011, 1, 21–29. [Google Scholar]
- Iamsaard, S.; Sukhorum, W.; Samrid, R.; Yimdee, J.; Kanla, P.; Chaisiwamongkol, K.; Hipkaeo, W.; Fongmoon, D.; Kondo, H. The sensitivity of male rat reproductive organs to monosodium glutamate. Acta Med. Acad. 2014, 43, 3–9. [Google Scholar] [CrossRef]
- Ulfanov, O.; Cil, N. Protective effects of vitamin E on aluminium sulphate-induced testicular damage. 2020, 36, 215–227. Toxicol. Ind. Health 2020, 36, 215–227. [Google Scholar] [CrossRef]
- Utomo, B.; Daningtia, N.R.; Yuliani, G.A.; Yuniarti, W.M. Effects of a standardized 40% ellagic acid pomegranate (Punica granatum L.) extract on seminiferous tubule histopathology, diameter, and epithelium thickness in albino Wistar rats after heat exposure. Vet. World 2019, 12, 1261–1265. [Google Scholar] [CrossRef]
- Nosseir, N.S.; Ali, M.H.M.; Ebaid, H.M. A histological and morphometric study of monosodium glutamate toxic effect on testicular structure and potentiality of recovery in adult albino rats. Res. J. Biol. 2012, 2, 66–78. [Google Scholar]
- Chakraborty, S.P. Patho-physiological and toxicological aspects of monosodium glutamate. Toxicol. Mech. Methods 2019, 29, 389–396. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Du, S.; Xun, P.; Sharma, S.; Wang, H.; Zhai, F.; Popkin, B. Consumption of monosodium glutamate in relation to incidence of overweight in Chinese adults: China Health and Nutrition Survey (CHNS). Am. J. Clin. Nutr. 2011, 93, 1328–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahadoran, Z.; Mirmiran, P.; Ghasemi, A. Monosodium Glutamate (MSG)-Induced Animal Model of Type 2 Diabetes. Methods Mol. Biol. 2019, 1916, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Hernández Bautista, R.J.; Mahmoud, A.M.; Königsberg, M.; López Díaz Guerrero, N.E. Obesity: Pathophysiology, monosodium glutamate-induced model and anti-obesity medicinal plants. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 111, 503–516. [Google Scholar] [CrossRef]
- Simonyi, A.; Serfozo, P.; Lehmidi, T.M.; Cui, J.; Gu, Z.; Lubahn, D.B.; Sun, A.Y.; Sun, G.Y. The neuroprotective effects of apocynin. Front. Biosci. 2012, 4, 2183–2193. [Google Scholar] [CrossRef]
- Ozbek, O.; Altintas, R.; Polat, A.; Vardi, N.; Parlakpinar, H.; Sagir, M.; Duran, Z.R.; Yildiz, A. The protective effect of apocynin on testicular ischemia-reperfusion injury. J. Urol. 2015, 193, 1417–1422. [Google Scholar] [CrossRef]
- Sener, T.E.; Yuksel, M.; Ozyilmaz-Yay, N.; Ercan, F.; Akbal, C.; Simsek, F.; Sener, G. Apocynin attenuates testicular ischemia-reperfusion injury in rats. J. Pediatr. Surg. 2015, 50, 1382–1387. [Google Scholar] [CrossRef]
- Kavram Sarihan, K.; Yardimoğlu Yilmaz, M. Protective effects of apocynin on damaged testes of rats exposed to methotrexate. Turk. J. Med. Sci. 2020, 50, 1409–1420. [Google Scholar] [CrossRef]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef]
- Peña-Blanco, A.; García-Sáez, A.J. Bax, Bak and beyond—Mitochondrial performance in apoptosis. FEBS J. 2018, 285, 416–431. [Google Scholar] [CrossRef] [Green Version]
- Shaha, C.; Tripathi, R.; Mishra, D.P. Male germ cell apoptosis: Regulation and biology. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2010, 365, 1501–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Saleh, F.; Khashab, F.; Fadel, F.; Al-Kandari, N.; Al-Maghrebi, M. Inhibition of NADPH oxidase alleviates germ cell apoptosis and ER stress during testicular ischemia reperfusion injury. Saudi J. Biol. Sci. 2020, 27, 2174–2184. [Google Scholar] [CrossRef] [PubMed]
- Fijak, M.; Meinhardt, A. The testis in immune privilege. Immunol. Rev. 2006, 213, 66–81. [Google Scholar] [CrossRef]
- Gerber, J.; Weider, K.; Hambruch, N.; Brehm, R. Loss of connexin43 (Cx43) in Sertoli cells leads to spatio-temporal alterations in occludin expression. Histol. Histopathol. 2014, 29, 935–948. [Google Scholar] [CrossRef] [PubMed]
- Saitou, M.; Furuse, M.; Sasaki, H.; Schulzke, J.D.; Fromm, M.; Takano, H.; Noda, T.; Tsukita, S. Complex phenotype of mice lacking occludin, a component of tight junction strands. Mol. Biol. Cell 2000, 11, 4131–4142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolbasi, B.; Bulbul, M.V.; Karabulut, S.; Altun, C.E.; Cakici, C.; Ulfer, G.; Mudok, T.; Keskin, I. Chronic unpredictable stress disturbs the blood-testis barrier affecting sperm parameters in mice. Reprod. Biomed. Online 2021, 42, 983–995. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acikel-Elmas, M.; Algilani, S.A.; Sahin, B.; Bingol Ozakpinar, O.; Gecim, M.; Koroglu, K.; Arbak, S. Apocynin Ameliorates Monosodium Glutamate Induced Testis Damage by Impaired Blood-Testis Barrier and Oxidative Stress Parameters. Life 2023, 13, 822. https://doi.org/10.3390/life13030822
Acikel-Elmas M, Algilani SA, Sahin B, Bingol Ozakpinar O, Gecim M, Koroglu K, Arbak S. Apocynin Ameliorates Monosodium Glutamate Induced Testis Damage by Impaired Blood-Testis Barrier and Oxidative Stress Parameters. Life. 2023; 13(3):822. https://doi.org/10.3390/life13030822
Chicago/Turabian StyleAcikel-Elmas, Merve, Salva Asma Algilani, Begum Sahin, Ozlem Bingol Ozakpinar, Mert Gecim, Kutay Koroglu, and Serap Arbak. 2023. "Apocynin Ameliorates Monosodium Glutamate Induced Testis Damage by Impaired Blood-Testis Barrier and Oxidative Stress Parameters" Life 13, no. 3: 822. https://doi.org/10.3390/life13030822
APA StyleAcikel-Elmas, M., Algilani, S. A., Sahin, B., Bingol Ozakpinar, O., Gecim, M., Koroglu, K., & Arbak, S. (2023). Apocynin Ameliorates Monosodium Glutamate Induced Testis Damage by Impaired Blood-Testis Barrier and Oxidative Stress Parameters. Life, 13(3), 822. https://doi.org/10.3390/life13030822