

The Universally Conserved Unconventional G Protein YchF Is Critical for Growth and Stress Response
 , and
, and
Abstract
:1. Introduction
2. Structure of G Domain among G Proteins
2.1. Structural Characterization of G Domain of G Proteins
2.2. Structural Characterization of G Domain of YchF
2.3. Structural Comparison of G Domains among Selected YchF, Small G Protein, and Heterotrimeric G Protein α-Subunit
3. YchF Is Critical for Growth and Stress Response
3.1. YchF Works as a Conserved Negative Regulator in Response to Oxidative Stress
3.2. YchF Is Crucial for Environmental Stress Response
3.3. YchF Bridges Protein Biosynthesis and Degradation
3.4. YchF Is a Key Molecule in Maintaining Proteostasis
4. OsYchF1/AtYchF1 and Its Activator OsGAP1/AtGAP1 in Plants
5. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheung, M.Y.; Xue, Y.; Zhou, L.; Li, M.W.; Sun, S.S.M.; Lam, H.M. An ancient P-loop GTPase in rice is regulated by a higher plant-specific regulatory protein. J. Biol. Chem. 2010, 285, 37359–37369. [Google Scholar] [CrossRef] [PubMed]
- Yung, Y.L.; Cheung, M.Y.; Miao, R.; Fong, Y.H.; Li, K.P.; Yu, M.H.; Chye, M.L.; Wong, K.B.; Lam, H.M. Site-directed mutagenesis shows the significance of interactions with phospholipids and the G-protein OsYchF1 on the physiological functions of the rice GTPase-Activating Protein 1 (OsGAP1). J. Biol. Chem. 2015, 290, 23984–23996. [Google Scholar] [CrossRef] [PubMed]
- Buglino, J.; Shen, V.; Hakimian, P.; Lima, C.D. Structural and biochemical analysis of the Obg GTP binding protein. Structure 2002, 10, 1581–1592. [Google Scholar] [CrossRef] [PubMed]
- Assmann, S.M. Heterotrimeric and unconventional GTP binding proteins in plant cell signaling. Plant Cell 2002, 14 (Suppl. S1), S355–S373. [Google Scholar] [PubMed]
- Wennerberg, K.; Rossman, K.L.; Der, C.J. The Ras superfamily at a glance. J. Cell Sci. 2005, 118 Pt 5, 843–846. [Google Scholar]
- Milligan, G.; Kostenis, E. Heterotrimeric G-proteins: A short history. Br. J. Pharmacol. 2009, 147, 46–55. [Google Scholar] [CrossRef]
- Sprang, S.R. G protein mechanisms: Insights from structural analysis. Annu. Rev. Biochem. 1997, 66, 639–678. [Google Scholar] [CrossRef]
- Leipe, D.D.; Wolf, Y.I.; Koonin, E.V.; Aravind, L. Classification and evolution of P-loop GTPases and related ATPases. J. Mol. Biol. 2002, 317, 41–72. [Google Scholar]
- Luo, M.; Han, Z.; Huang, G.; Li, R.; Liu, Y.; Lu, J.; Liu, L.; Miao, R. Structural comparison of unconventional G protein YchF with heterotrimeric G protein and small G protein. Plant Signal. Behav. 2022, 17, 2024405. [Google Scholar] [CrossRef]
- Oldham, W.M.; Hamm, H.E. Heterotrimeric G protein activation by G-protein-coupled receptors. Nat. Rev. Mol. Cell. Biol. 2008, 9, 60–71. [Google Scholar] [CrossRef]
- Ma, Y.; Dai, X.; Xu, Y.; Luo, W.; Zheng, X.; Zeng, D.; Pan, Y.; Lin, X.; Liu, H.; Zhang, D.; et al. COLD1 confers chilling tolerance in rice. Cell 2015, 160, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Maruta, N.; Trusov, Y.; Jones, A.M.; Botella, J.R. Heterotrimeric G Proteins in Plants: Canonical and Atypical Gα Subunits. Int. J. Mol. Sci. 2021, 22, 11841. [Google Scholar] [CrossRef] [PubMed]
- Reiner, D.J.; Lundquist, E.A. Small GTPases. WormBook 2018, 16, 1–65. [Google Scholar] [CrossRef] [PubMed]
- Koller-Eichhorn, R.; Marquardt, T.; Gail, R.; Wittinghofer, A.; Kostrewa, D.; Kutay, U.; Kambach, C. Human OLA1 defines an ATPase subfamily in the Obg family of GTP-binding proteins. J. Biol. Chem. 2007, 282, 19928–19937. [Google Scholar] [CrossRef] [PubMed]
- Rosler, K.S.; Mercier, E.; Andrews, I.C.; Wieden, H.J. Histidine 114 Is Critical for ATP Hydrolysis by the Universally Conserved ATPase YchF. J. Biol. Chem. 2015, 290, 18650–18661. [Google Scholar] [CrossRef] [PubMed]
- Hannemann, L.; Suppanz, I.; Ba, Q.; MacInnes, K.; Drepper, F.; Warscheid, B.; Koch, H.G. Redox Activation of the Universally Conserved ATPase YchF by Thioredoxin 1. Antioxid. Redox Signal. 2016, 24, 141–156. [Google Scholar] [CrossRef]
- Tomar, S.K.; Kumar, P.; Prakash, B. Deciphering the catalytic machinery in a universally conserved ribosome binding ATPase YchF. Biochem. Biophys. Res. Commun. 2011, 408, 459–464. [Google Scholar] [CrossRef]
- Wenk, M.; Ba, Q.; Erichsen, V.; MacInnes, K.; Wiese, H.; Warscheid, B.; Koch, H.G. A universally conserved ATPase regulates the oxidative stress response in Escherichia coli. J. Biol. Chem. 2012, 287, 43585–43598. [Google Scholar] [CrossRef]
- Cheung, M.Y.; Ngo, J.C.; Chen, Z.; Jia, Q.; Li, T.; Gou, Y.; Wang, Y.; Lam, H.M. A structure model explaining the binding between a ubiquitous unconventional G-protein (OsYchF1) and a plant-specific C2-domain protein (OsGAP1) from rice. Biochem. J. 2020, 477, 3935–3949. [Google Scholar] [CrossRef]
- Sun, H.; Luo, X.; Montalbano, J.; Jin, W.; Shi, J.; Sheikh, M.S.; Huang, Y. DOC45, a novel DNA damage-regulated nucleocytoplasmic ATPase that is overexpressed in multiple human malignancies. Mol. Cancer Res. 2010, 8, 57–66. [Google Scholar] [CrossRef]
- Cheung, M.Y.; Li, M.W.; Yung, Y.L.; Wen, C.Q.; Lam, H.M. The unconventional P-loop NTPase OsYchF1 and its regulator OsGAP1 play opposite roles in salinity stress tolerance. Plant Cell Environ. 2013, 36, 2008–2020. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.Y.; Li, X.; Miao, R.; Fong, Y.H.; Li, K.P.; Yung, Y.L.; Yu, M.H.; Wong, K.B.; Chen, Z.; Lam, H.M. ATP binding by the P-loop NTPase OsYchF1 (an unconventional G protein) contributes to biotic but not abiotic stress responses. Proc. Natl. Acad. Sci. USA 2016, 113, 2648–2653. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.Y.; Li, X.; Ku, Y.S.; Chen, Z.; Lam, H.M. Co-crystalization reveals the interaction between AtYchF1 and ppGpp. Front. Mol. Biosci. 2022, 9, 1061350. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.; Gzyl, K.E.; Altamirano, A.M.; Vuong, A.; Urban, K.; Wieden, H.J. The 70S ribosome modulates the ATPase activity of Escherichia coli YchF. RNA Biol. 2012, 9, 1288–1301. [Google Scholar] [CrossRef] [PubMed]
- Brandon, H.E. Biophysical Studies of the Universally Conserved NTPases HflX and YchF; University of Lethbridge: Lethbridge, AB, Canada, 2021. [Google Scholar]
- Gradia, D.F.; Rau, K.; Umaki, A.C.; de Souza, F.S.; Probst, C.M.; Correa, A.; Holetz, F.B.; Avila, A.R.; Krieger, M.A.; Goldenberg, S.; et al. Characterization of a novel Obg-like ATPase in the protozoan Trypanosoma cruzi. Int. J. Parasitol. 2009, 39, 49–58. [Google Scholar] [CrossRef]
- Guerrero, C.; Tagwerker, C.; Kaiser, P.; Huang, L. An integrated mass spectrometry-based proteomic approach: Quantitative analysis of tandem affinity-purified in vivo cross-linked protein complexes (QTAX) to decipher the 26 S proteasome-interacting network. Mol. Cell Proteom. 2005, 5, 366–378. [Google Scholar] [CrossRef] [PubMed]
- Teplyakov, A.; Obmolova, G.; Chu, S.Y.; Toedt, J.; Eisenstein, E.; Howard, A.J.; Gilliland, G.L. Crystal structure of the YchF protein reveals binding sites for GTP and nucleic acid. J. Bacteriol. 2003, 185, 4031–4037. [Google Scholar] [CrossRef]
- Gavin, A.C.; Bösche, M.; Krause, R.; Grandi, P.; Marzioch, M.; Bauer, A.; Schultz, J.; Rick, J.M.; Michon, A.M.; Cruciat, C.M.; et al. Functional organization of the yeast proteome by systematic analysis of protein complexes. Nature 2002, 415, 141–147. [Google Scholar] [CrossRef]
- Landwehr, V.; Milanov, M.; Angebauer, L.; Hong, J.; Jüngert, G.; Hiersemenzel, A.; Siebler, A.; Schmit, F.; Öztürk, Y.; Dannenmaier, S.; et al. The Universally Conserved ATPase YchF Regulates Translation of Leaderless mRNA in Response to Stress Conditions. Front. Mol. Biosci. 2021, 8, 643696. [Google Scholar] [CrossRef]
- Chen, H.; Song, R.; Wang, G.; Ding, Z.; Yang, C.; Zhang, J.; Zeng, Z.; Rubio, V.; Wang, L.; Zu, N.; et al. OLA1 regulates protein synthesis and integrated stress response by inhibiting eIF2 ternary complex formation. Sci. Rep. 2015, 5, 13241. [Google Scholar] [CrossRef]
- Mao, R.F.; Rubio, V.; Chen, H.; Bai, L.; Mansour, O.C.; Shi, Z.Z. OLA1 protects cells in heat shock by stabilizing HSP70. Cell Death Dis. 2013, 4, e491. [Google Scholar] [CrossRef]
- Schultz, A.; Olorundami, O.A.; Teng, R.J.; Jarzembowski, J.; Shi, Z.Z.; Kumar, S.N.; Pritchard KJr Konduri, G.G.; Afolayan, A.J. Decreased OLA1 (Obg-Like ATPase-1) Expression Drives Ubiquitin-Proteasome Pathways to Downregulate Mitochondrial SOD2 (Superoxide Dismutase) in Persistent Pulmonary Hypertension of the Newborn. Hypertension 2019, 74, 957–966. [Google Scholar] [CrossRef]
- Zhang, J.; Rubio, V.; Lieberman, M.W.; Shi, Z.Z. OLA1, an Obg-like ATPase, suppresses antioxidant response via nontranscriptional mechanisms. Proc. Natl. Acad. Sci. USA 2009, 106, 15356–15361. [Google Scholar] [CrossRef]
- Guan, N.; Shin, H.D.; Chen, R.R.; Li, J.; Liu, L.; Du, G.; Chen, J. Understanding of how Propionibacterium acidipropionici respond to propionic acid stress at the level of proteomics. Sci. Rep. 2014, 4, 6951. [Google Scholar] [CrossRef]
- Chen, Y.C.; Chung, Y.T. A conserved GTPase YchF of Vibrio vulnificus is involved in macrophage cytotoxicity, iron acquisition, and mouse virulence. Int. J. Med. Microbiol. 2011, 301, 469–474. [Google Scholar] [CrossRef]
- Dong, Y.; Yin, A.; Xu, C.; Jiang, H.; Wang, Q.; Wu, W.; Guo, S. OLA1 is a potential prognostic molecular biomarker for endometrial cancer and promotes tumor progression. Oncol. Lett. 2021, 22, 576. [Google Scholar] [CrossRef]
- Gibbs, M.R.; Moon, K.M.; Warner, B.R.; Chen, M.; Bundschuh, R.; Foster, L.J.; Fredrick, K. Functional Analysis of BipA in E. coli Reveals the Natural Plasticity of 50S Subunit Assembly. J. Mol. Biol. 2020, 432, 5259–5272. [Google Scholar] [CrossRef]
- Beck, H.J.; Moll, I. Leaderless mRNAs in the Spotlight: Ancient but Not Outdated! Microbiol. Spectr. 2018, 6, RWR-0016-2017. [Google Scholar] [CrossRef]
- Vesper, O.; Amitai, S.; Belitsky, M.; Byrgazov, K.; Kaberdina, A.C.; Engelberg-Kulka, H.; Moll, I. Selective translation of leaderless mRNAs by specialized ribosomes generated by MazF in Escherichia coli. Cell 2011, 147, 147–157. [Google Scholar] [CrossRef]
- Amitai, S.; Kolodkin-Gal, I.; Hananya-Meltabashi, M.; Sacher, A.; Engelberg-Kulka, H. Escherichia coli MazF leads to the simultaneous selective synthesis of both “death proteins” and “survival proteins”. PLoS Genet. 2009, 5, e1000390. [Google Scholar] [CrossRef]
- Rodnina, M.V.; Wintermeyer, W. Protein elongation, Co-translational folding and targeting. J. Mol. Biol. 2016, 428, 2165–2185. [Google Scholar] [CrossRef]
- Spivak, J.L.; Hasselbalch, H. Hydroxycarbamide: A user’s guide for chronic myeloproliferative disorders. Expert Rev. Anti Cancer Ther. 2011, 11, 403–414. [Google Scholar] [CrossRef]
- Iyer, L.M.; Burroughs, A.M.; Aravind, L. The prokaryotic antecedents of the ubiquitin-signaling system and the early evolution of ubiquitin-like beta-grasp domains. Genome Biol. 2006, 7, R60. [Google Scholar] [CrossRef]
- Miao, R.; Russinova, E.; Rodriguez, P.L. Tripartite hormonal regulation of plasma membrane H+-ATPase activity. Trends Plant Sci. 2022, 27, 588–600. [Google Scholar] [CrossRef]
- Miao, R.; Yuan, W.; Wang, Y.; Garcia-Maquilon, I.; Dang, X.; Li, Y.; Zhang, J.; Zhu, Y.; Rodriguez, P.L.; Xu, W. Low ABA concentration promotes root growth and hydrotropism through relief of ABA INSENSITIVE 1-mediated inhibition of plasma membrane H+-ATPase 2. Sci. Adv. 2021, 7, eabd4113. [Google Scholar] [CrossRef]
- Miao, R.; Wang, M.; Yuan, W.; Ren, Y.; Li, Y.; Zhang, N.; Zhang, J.; Kronzucker, H.J.; Xu, W. Comparative analysis of Arabidopsis ecotypes reveals a role for brassinosteroids in root hydrotropism. Plant Physiol. 2018, 176, 2720–2736. [Google Scholar] [CrossRef]
- Cheung, M.Y.; Zeng, N.Y.; Tong, S.W.; Li, W.Y.; Xue, Y.; Zhao, K.J.; Wang, C.; Zhang, Q.; Fu, Y.; Sun, Z.; et al. Constitutive expression of a rice GTPase-activating protein induces defense responses. New Phytol. 2008, 179, 530–545. [Google Scholar] [CrossRef]
- Ito, D.; Kawamura, H.; Oikawa, A.; Ihara, Y.; Shibata, T.; Nakamura, N.; Asano, T.; Kawabata, S.I.; Suzuki, T.; Masuda, S. ppGpp functions as an alarmone in metazoa. Commun. Biol. 2020, 3, 671. [Google Scholar] [CrossRef]
- Caldon, C.E.; Yoong, P.; March, P.E. Evolution of a molecular switch: Universal bacterial GTPases regulate ribosome function. Mol. Microbiol. 2001, 41, 289–297. [Google Scholar] [CrossRef]
- Cruz-Vera, L.R.; Galindo, J.M.; Guarneros, G. Transcriptional analysis of the gene encoding peptidyl-tRNA hydrolase in Escherichia coli. Microbiology 2002, 148 Pt 11, 3457–3466. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
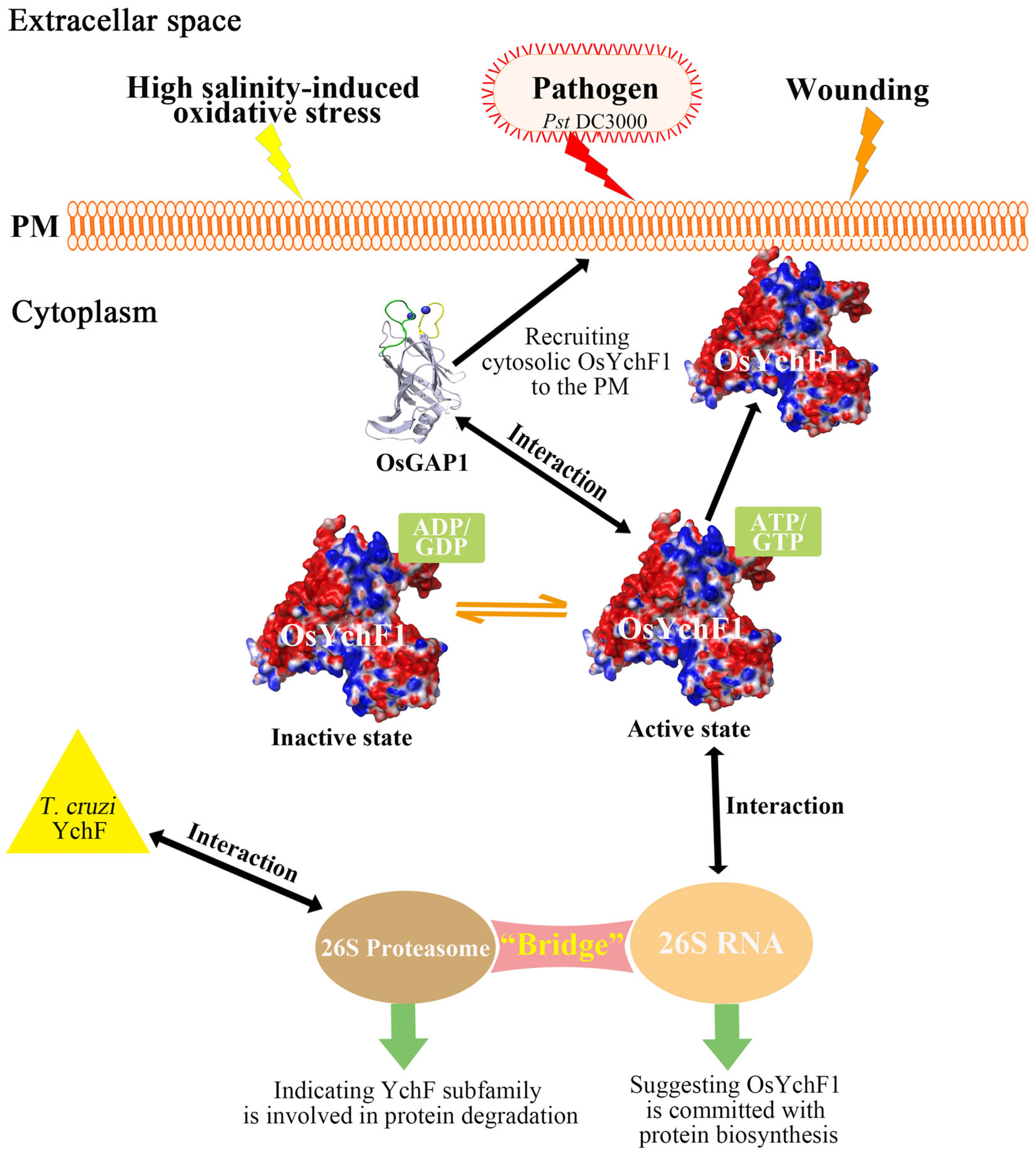{kind=link}
| Homolog | Species | Residues | Location | Supportive Reasons/Effects | Functions | References |
|---|---|---|---|---|---|---|
| E. coli YchF | Escherichia coli | His114 | A highly flexible loop of G domain | Supporting the flexible loop to reach a catalytically active conformation | Critical for ATPase activity (+) | [15] |
| E. coli YchF | Escherichia coli | Cys35 | G2 motif | Allows YchF dimerization via a disulfide bridge | Critical for ATPase activity (−) | [16] |
| E. coli YchF | Escherichia coli | Lys78 (Arg) | G domain | YchF-K78A mutant shows similar hydrolysis activities in presence of Na+ or K+, but K78R mutant retained potassium specific stimulation of ATPase activity | Plays a key role in determining the potassium dependent ATPase activity | [17] |
| hOLA1 | Homo sapiens | Leu96 | G domain (next to G3 motif) | A conserved Gln residue involved in GTP hydrolysis in Ras-like GTPases has been replaced | Inactivates Ras-like GTPases | [14] |
| E. coli YchF | Escherichia coli | Ser16 (Ser36 in H. sapiens) | G1 motif | Ser16 phosphorylated when H2O2 absence; Dissociation of KatG | Supports the ATPase activity; Detoxifies H2O2 | [14,18] |
| E. coli YchF | Escherichia coli | Leu76 | G3 motif | Hallmark for HAS-NTPase | Slightly affects ATPase activity (+) | [15] |
| hOLA1 | Homo sapiens | Thr37 | G domain | The main chain amide of Thr37 contacts the α-phosphate of AMPPCP | Supports the ATPase activity | [14] |
| hOLA1 | Homo sapiens | Ser36/Val33 | G1 motif | The main chain amide of Ser36 and Val33 contacts the β-phosphate of AMPPCP | Supports the ATPase activity | [14] |
| hOLA1 | Homo sapiens | Asn32 | G1 motif | The main chain amide of Asn32 forms a hydrogen bond to the γ-phosphate of AMPPCP | Supports the ATPase activity | [14] |
| hOLA1 | Homo sapiens | Asn230 | G4 motif | Its mutation to alanine abolished nucleotide binding | Contribute to nucleotide binding | [14] |
| hOLA1 | Homo sapiens | Leu231 | G4 motif | Specificity for adenine binding is based on the interaction between the adenine N-6 group and Leu231 main chain CO in G4 motif | Make YchF preference for ATP rather than GTP | [14] |
| hOLA1 | Homo sapiens | Ser310 | TGS domain | The H-bond between Ser310 Oγ and the exocyclic N-6 of an adenine is formed in a position similar to the ppGpp O-6 | Make YchF preference for ATP rather than GTP | [14] |
| hOLA1 | Homo sapiens | Phe127 | Coiled-coil domain | Mutating this residue to Ala diminishes ATP binding drastically | Contribute to base recognition | [14] |
| AtYchF1 | Arabidopsis thaliana | Glu345 | TGS domain | Conserved and solvent-exposed | Most critical for its interaction with the regulator, GAP1 | [19] |
| Homolog | Organism | Interactive Factors | Effects | References |
|---|---|---|---|---|
| OsYchF1 | Oryza sativa | OsGAP1 | Activating OsYchF1 GTPase and ATPase activity | [2,19,21,22] |
| AtYchF1 | Arabidopsis thaliana | AtGAP1 | Activating AtYchF1 GTPase and ATPase activity | [2,19,21,22] |
| AtYchF1 | Arabidopsis thaliana | ppGpp | AtYchF1 might be a critical regulator in controlling the cytosolic ppGpp-mediated growth inhibition in plants | [23] |
| E. coli YchF | Escherichia coli | 30S ribosome, 70S ribosome | The 70S ribosome act as an ATPase activating factor (AAF) to stimulate YchF’s ATPase activity | [17,24] |
| E. coli YchF | Escherichia coli | tRNA | YchF interacts with the 3′-CCA end of tRNA through its TGS-domain, indicating that YchF is involved in protein synthesis | [25] |
| T. cruzi YchF | Trypanosoma cruzi | 26S Proteasome | T. cruzi YchF co-immunoprecipitates with a regulatory subunit of the T. cruzi proteasome, involving in protein degradation | [26,27] |
| E. coli YchF | Escherichia coli | KatG | YchF interacts with KatG and inhibit its catalase activity, revealing that YchF regulates the oxidative stress response | [16] |
| S. cerevisiae YchF | Saccharomyces cerevisiae | Eukaryotic translation elongation factor 1 (eEF1) | Supporting a role for YchF during translation | [28,29] |
| E. coli YchF | Escherichia coli | Translation initiation factor 3 (IF3) | YchF enhances the anti-association activity of IF3, stimulates the translation of leaderless mRNAs | [30] |
| E. coli YchF | Escherichia coli | Thioredoxin 1 (TrxA) | YchF dimer is dissociated by TrxA, which stimulates the ATPase activity | [16] |
| hOLA1 | Homo sapiens | Eukaryotic elongation initiation factor 2 (eIF2) | hOLA1 effectively blocks the formation of TC (ternary complex) through its intrinsic GTPase activity, leading eIF2 unable to deliver Met-tRNAiMet to the 40S ribosome to initiate translation | [31] |
| hOLA1 | Homo sapiens | Heat shock protein 70 (HSP70) | OLA1 can interfere with the binding and function of the E3 ligase CHIP to HSP70, leading to the stabilization of HSP70, and response to heat shock | [32] |
| hOLA1 | Homo sapiens | Superoxide dismutase 2 (SOD2) | OLA1 deficiency can enhance CHIP affinity for HSP70-SOD2 complexes, facilitating SOD2 degradation, supporting OLA1 plays a role in response to mitochondrial oxidative stress | [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Z.; Li, R.; Han, Z.; Liu, Y.; Gao, L.; Huang, S.; Miao, Y.; Miao, R. The Universally Conserved Unconventional G Protein YchF Is Critical for Growth and Stress Response. Life 2023, 13, 1058. https://doi.org/10.3390/life13041058
Lin Z, Li R, Han Z, Liu Y, Gao L, Huang S, Miao Y, Miao R. The Universally Conserved Unconventional G Protein YchF Is Critical for Growth and Stress Response. Life. 2023; 13(4):1058. https://doi.org/10.3390/life13041058
Chicago/Turabian StyleLin, Zhaoheng, Rongfang Li, Zhiwei Han, Yi Liu, Liyang Gao, Suchang Huang, Ying Miao, and Rui Miao. 2023. "The Universally Conserved Unconventional G Protein YchF Is Critical for Growth and Stress Response" Life 13, no. 4: 1058. https://doi.org/10.3390/life13041058
APA StyleLin, Z., Li, R., Han, Z., Liu, Y., Gao, L., Huang, S., Miao, Y., & Miao, R. (2023). The Universally Conserved Unconventional G Protein YchF Is Critical for Growth and Stress Response. Life, 13(4), 1058. https://doi.org/10.3390/life13041058