
Antioxidant and Anti-Inflammatory Activities of Stellera chamaejasme L. Roots and Aerial Parts Extracts

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. S. chamaejasme Extracts Preparation
2.3. Characterization of S. chamaejasme Extracts Composition
2.3.1. TLC Analysis
2.3.2. GC-MS Analysis
2.4. Antioxidant Activity Evaluation
2.4.1. DPPH• Scavenging Assay
2.4.2. ABTS•+ Scavenging Assay
2.4.3. Antioxidant Activity against ROO•
2.5. Biological Studies
2.5.1. S. chamaejasme Extracts Solutions Preparation
2.5.2. Anti-Inflammatory Activity Evaluation
2.5.3. Metabolic Activity
2.5.4. DNA Quantification
2.6. Statistical Analysis
3. Results
3.1. Extraction Yield
3.2. Chemical Composition of S. chamaejasme Extracts
3.2.1. TLC Analysis of S. chamaejasme Extracts
3.2.2. GC-MS Analysis of S. chamaejasme DCM-R Extracts
3.3. Extracts Antioxidant Activity
3.3.1. Antiradical Activity of S. chamaejasme Extracts against DPPH• and ABTS•+
3.3.2. Antioxidant Activity of S. chamaejasme Extracts against ROO•
3.4. Anti-Inflammatory Activity Evaluation
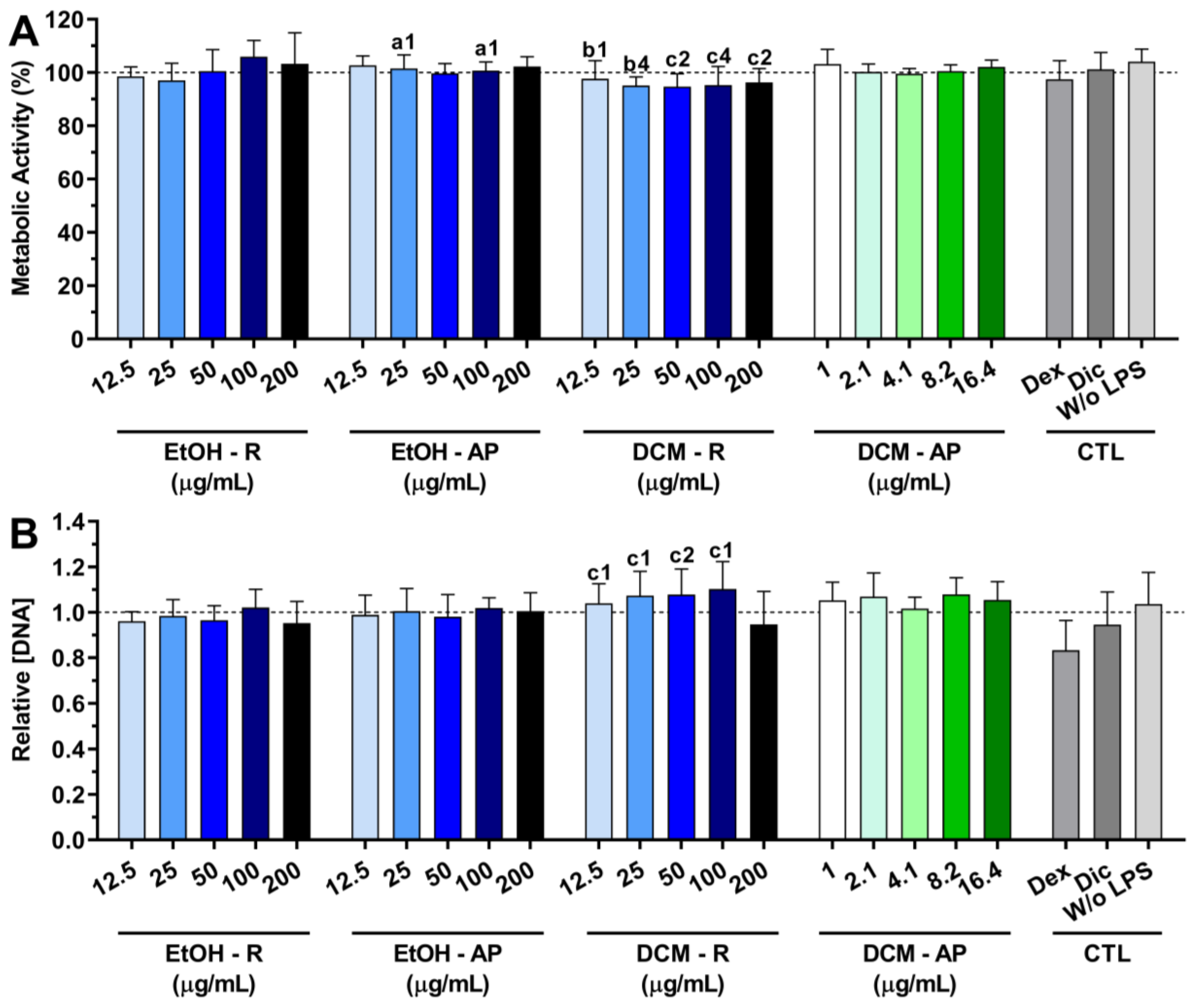
3.4.1. Cytocompatibility of S. chamaejasme Extracts
3.4.2. Anti-Inflammatory Activity of S. chamaejasme Extracts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.H.; Yue, J.P.; Sun, H. Identification of Twelve Novel Polymorphic Microsatellite Loci in the Severe Weed, Stellera chamaejasme L. (Thymelaeaceae). J. Genet. 2015, 94, 24–26. [Google Scholar] [CrossRef]
- Volodya, T.; Tserenbaljir, D.; Lamjav, T. Medicinal Plant of Mongolia, 2nd ed.; ADMON: Ulaanbaatar, Mongolia, 2010; ISBN 978-99929-0-431-3. [Google Scholar]
- Kim, M.; Lee, H.J.; Randy, A.; Yun, J.H.; Oh, S.R.; Nho, C.W. Stellera chamaejasme and Its Constituents Induce Cutaneous Wound Healing and Anti-Inflammatory Activities. Sci. Rep. 2017, 7, 42490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Medicinal Plants in Mongolia; World Health Organization, Western Pacific Region, WHO Regional Office for the Western Pacific: Geneva, Switzerland, 2013; ISBN 9789290616320.
- Biswas, S.K. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxid. Med. Cell Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.; Chen, Y.; Zhong, S.; Ji, B.; Wang, J.; Bai, X.; Shi, G. Anti-Inflammatory Activity of N-Butanol Extract from Ipomoea Stolonifera in Vivo and in Vitro. PLoS ONE 2014, 9, e95931. [Google Scholar] [CrossRef] [PubMed]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free Radicals, Antioxidants in Disease and Health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Nariya, P.; Nariya, M.; Shukla, V.; Acharya, R.; Bhalodia, N. In Vitro Evaluation of Antioxidant Activity of Cordia Dichotoma (Forst f.) Bark. AYU (Int. Q. J. Res. Ayurveda) 2013, 34, 124–128. [Google Scholar] [CrossRef] [Green Version]
- Vieira, S.F.; Ferreira, H.; Neves, N.M. Antioxidant and Anti-Inflammatory Activities of Cytocompatible Salvia Officinalis Extracts: A Comparison between Traditional and Soxhlet Extraction. Antioxidants 2020, 9, 1157. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhu, X. Stellera chamaejasme L. Extract Induces Apoptosis of Human Lung Cancer Cells via Activation of the Death Receptor-Dependent Pathway. Exp. Ther. Med. 2012, 4, 605–610. [Google Scholar] [CrossRef]
- Calixto, J.B. The Role of Natural Products in Modern Drug Discovery. An. Acad. Bras. Cienc. 2019, 91, e20190105. [Google Scholar] [CrossRef]
- Li, X.-Q.; Rahman, K.; Zhu, J.-Y.; Zhang, H. Chemical Constituents and Pharmacological Activities of Stellera chamaejasme. Curr. Pharm. Des. 2018, 24, 2825–2838. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yang, Q.; Zhang, G.; Li, Y.; Chen, Y.; Weng, X.; Wang, Y.; Wang, Y.; Zhu, X. Anti-Tumor Pharmacological Evaluation of Extracts from Stellera chamaejasme L Based on Hollow Fiber Assay. BMC Complement. Altern. Med. 2014, 14, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsolmon, S.; Han, J.; Isoda, H. Inhibition of Cell Growth by Stellera chamaejasme Extract Is Associated with Induction of Autophagy and Differentiation in Chronic Leukemia K562 Cells. J. Biosci. Bioeng. 2010, 110, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Lu, Y.; Chen, C.H.; Zhao, Y.; Lee, K.H.; Chen, D.F. Stelleralides D-J and Anti-HIV Daphnane Diterpenes from Stellera chamaejasme. J. Nat. Prod. 2015, 78, 2712–2718. [Google Scholar] [CrossRef] [Green Version]
- Shagjjav, O.; Bhattarai, H.D.; Yim, J.H.; Dondog, P. Antioxidant Activity of Some Mongolian Plants. Mong. J. Biol. Sci. 2014, 12, 27–32. [Google Scholar] [CrossRef]
- Venkatesan, T.; Choi, Y.W.; Kim, Y.K. Impact of Different Extraction Solvents on Phenolic Content and Antioxidant Potential of Pinus Densiflora Bark Extract. Biomed. Res. Int. 2019, 2019, 3520675. [Google Scholar] [CrossRef] [Green Version]
- Mai, C.; Sci, J. Stellera chamaejasme Methanolic Extract Attenuates Nitric Oxide Production and Enhances Heme Oxygenase 1 Expression in Murine Macrophages. Chiang Mai J. Sci. 2017, 44, 858–868. [Google Scholar]
- Ma, L.; Wu, H.; Bai, R.; Zhou, L.; Yuan, X.; Hou, D. Phytotoxic Effects of Stellera chamaejasme L. Root Extract. Afr. J. Agric. Res. 2011, 6, 1170–1176. [Google Scholar]
- Jo, B.G.; Bong, S.K.; Jegal, J.; Kim, S.N.; Yang, M.H. Antiallergic Effects of Phenolic Compounds Isolated from Stellera chamaejasme on RBL-2H3 Cells. Nat. Prod. Commun. 2020, 15, 1934578X20942352. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, J.; Wang, X.; Wang, H. Glycoglycerolipids from Stellera chamaejasme. Nat. Prod. Commun. 2012, 7, 1499–1500. [Google Scholar] [CrossRef] [Green Version]
- Vieira, S.F.; Gonçalves, V.M.F.; Llaguno, C.P.; Macías, F.; Tiritan, M.E.; Reis, R.L.; Ferreira, H.; Neves, N.M. On the Bioactivity of Echinacea Purpurea Extracts to Modulate the Production of Inflammatory Mediators. Int. J. Mol. Sci. 2022, 23, 13616. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Bladt, S.; Zgainski, E.M. Plant Drug Analysis, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1984; Volume XIII, ISBN 978-3-662-02400-3. [Google Scholar]
- Cidade, H.; Rocha, V.; Palmeira, A.; Marques, C.; Tiritan, M.E.; Ferreira, H.; Lobo, J.S.; Almeida, I.F.; Sousa, M.E.; Pinto, M. In Silico and in Vitro Antioxidant and Cytotoxicity Evaluation of Oxygenated Xanthone Derivatives. Arab. J. Chem. 2020, 13, 17–26. [Google Scholar] [CrossRef]
- Huyut, Z.; Beydemir, Ş.; Gülçin, I. Antioxidant and Antiradical Properties of Selected Flavonoids and Phenolic Compounds. Biochem. Res. Int. 2017, 2017, 7616791. [Google Scholar] [CrossRef] [Green Version]
- Lúcio, M.; Nunes, C.; Gaspar, D.; Ferreira, H.; Lima, J.L.F.C.; Reis, S. Antioxidant Activity of Vitamin E and Trolox: Understanding of the Factors That Govern Lipid Peroxidation Studies in Vitro. Food Biophys. 2009, 4, 312–320. [Google Scholar] [CrossRef]
- Cao, G.; Alessio, H.M.; Cutler, R.G. Oxygen-Radical Absorbance Capacity Assay for Antioxidants. Free Radic. Biol. Med. 1993, 14, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Genin, M.; Clement, F.; Fattaccioli, A.; Raes, M.; Michiels, C. M1 and M2 Macrophages Derived from THP-1 Cells Differentially Modulate the Response of Cancer Cells to Etoposide. BMC Cancer 2015, 15, 577. [Google Scholar] [CrossRef] [Green Version]
- Borquaye, L.S.; Laryea, M.K.; Gasu, E.N.; Boateng, M.A.; Baffour, P.K.; Kyeremateng, A.; Doh, G. Anti-Inflammatory and Antioxidant Activities of Extracts of Reissantia indica, Cissus cornifolia and Grosseria vignei. Cogent Biol. 2020, 6, 1785755. [Google Scholar] [CrossRef]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant Activity and Phenolic Compounds of 112 Traditional Chinese Medicinal Plants Associated with Anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef]
- Truong, D.H.; Nguyen, D.H.; Ta, N.T.A.; Bui, A.V.; Do, T.H.; Nguyen, H.C. Evaluation of the Use of Different Solvents for Phytochemical Constituents, Antioxidants, and in Vitro Anti-Inflammatory Activities of Severinia buxifolia. J. Food Qual. 2019, 2019, 8178294. [Google Scholar] [CrossRef] [Green Version]
- Yang, G. Bioactive Constituents of Stellera chamaejasme, Zanthoxylum nitidum and Geranium strictipes. PhD Dissertation, FuDan University, Shanghai, China, 2005. [Google Scholar]
- Jiang, Z.H.; Tanaka, T.; Sakamoto, T.; Kouno, I.; Duan, J.A.; Zhou, R.H. Biflavanones, Diterpenes, and Coumarins from the Roots of Stellera chamaejasme L. Chem. Pharm. Bull. 2002, 50, 137–139. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Chen, D. Biflavanones, Flavonoids, and Coumarins from the Roots of Stellera chamaejasme and Their Antiviral Effect on Hepatits B Virus. Chem. Biodivers. 2008, 5, 1419–1424. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Bladt, S. Drugs Containing Essential Oils (Aetherolea), Balsams and Oleo-Gum-Resins. In Plant Drug Analysis; Springer: Berlin/Heidelberg, Germany, 1996; Volume XV, pp. 149–193. [Google Scholar]
- Shirai, K.; Okamoto, Y.; Tori, M.; Kawahara, T.; Gong, X.; Noyama, T.; Watanabe, E.; Kuroda, C. Diversity in the Flavonoid Composition of Stellera chamaejasme in the Hengduan Mountains. Nat. Prod. Commun. 2015, 10, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Tatematsu, H.; Kurokawa, M.; Niwa, M.; Hirata, Y. Piscicidal Constituents of Stellera chamaejasme L. II. Chem Pharm Bull 1984, 32, 1612–1613. [Google Scholar] [CrossRef] [Green Version]
- Asada, Y.; Sukemori, A.; Watanabe, T.; Malla, K.J.; Yoshikawa, T.; Li, W.; Kuang, X.; Koike, K.; Chen, C.H.; Akiyama, T.; et al. Isolation, Structure Determination, and Anti-HIV Evaluation of Tigliane-Type Diterpenes and Biflavonoid from Stellera chamaejasme. J. Nat. Prod. 2013, 76, 852–857. [Google Scholar] [CrossRef] [Green Version]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef] [Green Version]
- Masood, N.; Fatima, K.; Luqman, S. A Modified Method for Studying Behavioral Paradox of Antioxidants and Their Disproportionate Competitive Kinetic Effect to Scavenge the Peroxyl Radical Formation. Sci. World J. 2014, 2014, 931581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, H.; Pengal, R.A.; Cao, X.; Ganesan, L.P.; Wewers, M.D.; Marsh, C.B.; Tridandapani, S. Lipopolysaccharide-Induced Macrophage Inflammatory Response Is Regulated by SHIP. J. Immunol. 2004, 173, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Araki, H.; Imaoka, A.; Kuboyama, N.; Abiko, Y. Reduction of Interleukin-6 Expression in Human Synoviocytes and Rheumatoid Arthritis Rat Joints by Linear Polarized near Infrared Light (SuperLizer) Irradiation. Laser Ther. 2011, 20, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Leyva-López, N.; Gutierrez-Grijalva, E.P.; Ambriz-Perez, D.L.; Basilio Heredia, J. Flavonoids as Cytokine Modulators: A Possible Therapy for Inflammation-Related Diseases. Int. J. Mol. Sci. 2016, 17, 921. [Google Scholar] [CrossRef] [Green Version]
- Wassmann, S.; Stumpf, M.; Strehlow, K.; Schmid, A.; Schieffer, B.; Böhm, M.; Nickenig, G. Interleukin-6 Induces Oxidative Stress and Endothehal Dysfunction by Overexpression of the Angiotensin II Type 1 Receptor. Circ. Res. 2004, 94, 534–541. [Google Scholar] [CrossRef]
- Cárdeno, A.; Aparicio-Soto, M.; Montserrat-de la Paz, S.; Bermudez, B.; Muriana, F.J.G.; Alarcón-de-la-Lastra, C. Squalene Targets Pro- and Anti-Inflammatory Mediators and Pathways to Modulate over-Activation of Neutrophils, Monocytes and Macrophages. J. Funct. Foods 2015, 14, 779–790. [Google Scholar] [CrossRef] [Green Version]
- Krishnamoorthy, K.; Subramaniam, P. Phytochemical Profiling of Leaf, Stem, and Tuber Parts of Solena amplexicaulis (Lam.) Gandhi Using GC-MS. Int. Sch. Res. Not. 2014, 2014, 67409. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| № | Standards | Extracts | |||
|---|---|---|---|---|---|
| EtOH-AP | EtOH-R | DCM-AP | DCM-R | ||
| 1 | Kaempferol | + | - | - | - |
| 2 | Quercetin-3-O-glucopyranoside | + | - | - | - |
| 3 | Coumarin | - | - | - | - |
| 4 | Luteolin | - | - | - | - |
| 5 | Rutin | - | - | - | - |
| 6 | Morin | - | - | - | - |
| 7 | Riboflavin | - | - | - | - |
| Peak Number | Retention Time (min) | Identified Chemical Compounds | Molecular Formula | Molecular Weight | Peak Area (%) |
|---|---|---|---|---|---|
| 1 | 17.235 | 1-Hexadecanol | C16H34O | 242.4 | 1.1 |
| 2 | 18.034 | Palmitic Acid | C16H32O2 | 256.4 | 2.9 |
| 3 | 19.541 | 9-Octadecenoic acid | C18H34O2 | 282.5 | 31.8 |
| 4 | 24.013 | Squalene | C30H50 | 410.7 | 1.9 |
| 5 | 27.166 | Stigmast-5-ene, 3β-(trimethylsiloxy) | C32H58OSi | 486.9 | 5.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selenge, T.; Vieira, S.F.; Gendaram, O.; Reis, R.L.; Tsolmon, S.; Tsendeekhuu, E.; Ferreira, H.; Neves, N.M. Antioxidant and Anti-Inflammatory Activities of Stellera chamaejasme L. Roots and Aerial Parts Extracts. Life 2023, 13, 1654. https://doi.org/10.3390/life13081654
Selenge T, Vieira SF, Gendaram O, Reis RL, Tsolmon S, Tsendeekhuu E, Ferreira H, Neves NM. Antioxidant and Anti-Inflammatory Activities of Stellera chamaejasme L. Roots and Aerial Parts Extracts. Life. 2023; 13(8):1654. https://doi.org/10.3390/life13081654
Chicago/Turabian StyleSelenge, Temuulen, Sara F. Vieira, Odontuya Gendaram, Rui L. Reis, Soninkhishig Tsolmon, Enkhtuul Tsendeekhuu, Helena Ferreira, and Nuno M. Neves. 2023. "Antioxidant and Anti-Inflammatory Activities of Stellera chamaejasme L. Roots and Aerial Parts Extracts" Life 13, no. 8: 1654. https://doi.org/10.3390/life13081654
APA StyleSelenge, T., Vieira, S. F., Gendaram, O., Reis, R. L., Tsolmon, S., Tsendeekhuu, E., Ferreira, H., & Neves, N. M. (2023). Antioxidant and Anti-Inflammatory Activities of Stellera chamaejasme L. Roots and Aerial Parts Extracts. Life, 13(8), 1654. https://doi.org/10.3390/life13081654