
Traditional Therapeutics and Potential Epidrugs for CVD: Why Not Both?


 and
and
Abstract
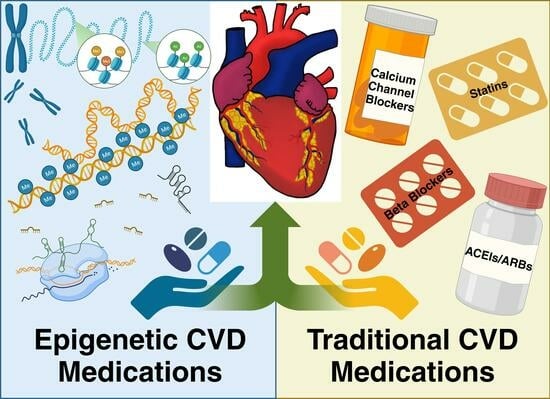
:
1. Introduction
2. CVD Risk Factors
2.1. Obesity
2.2. Type 2 Diabetes Mellitus (T2DM)
2.3. Smoking
2.4. Hypertension
2.5. Hyperlipidemia/Atherosclerosis
3. Epigenetics
4. Traditional CVD Medications
4.1. Statins
4.2. Calcium Channel Blockers
4.3. Beta Blockers
4.4. ACEIs/ARBs
5. Epigenetic CVD Medications
5.1. HDAC Inhibitors (HDACIs)
5.1.1. Valproic Acid
5.1.2. Sodium Phenylbutyrate
5.1.3. Vorinostat
5.1.4. Trichostatin A
5.2. SIRT Family + Sirtuin-Activating Compounds (STACs)
Resveratrol
5.3. HAT Inhibitors (HATIs)
Curcumin
5.4. Interventional Clinical Trial Updates of Epigenetic Drugs
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vaduganathan, M.; Mensah George, A.; Turco Justine, V.; Fuster, V.; Roth Gregory, A. The Global Burden of Cardiovascular Diseases and Risk. J. Am. Coll. Cardiol. 2022, 80, 2361–2371. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Heart Disease Facts. Available online: https://www.cdc.gov/heartdisease/facts.htm (accessed on 27 September 2023).
- American Heart Association Statistics, Committee. What Is Cardiovascular Disease. Available online: https://www.heart.org/en/health-topics/consumer-healthcare/what-is-cardiovascular-disease (accessed on 15 January 2023).
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990–2019: Update From the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef] [PubMed]
- Tsao, C.W.; Aday, A.W.; Almarzooq, Z.I.; Anderson, C.A.M.; Arora, P.; Avery, C.L.; Baker-Smith, C.M.; Beaton, A.Z.; Boehme, A.K.; Buxton, A.E.; et al. Heart Disease and Stroke Statistics—2023 Update: A Report From the American Heart Association. Circulation 2023, 147, e93–e621. [Google Scholar] [CrossRef] [PubMed]
- Powell-Wiley, T.M.; Poirier, P.; Burke, L.E.; Després, J.-P.; Gordon-Larsen, P.; Lavie, C.J.; Lear, S.A.; Ndumele, C.E.; Neeland, I.J.; Sanders, P.; et al. Obesity and Cardiovascular Disease: A Scientific Statement From the American Heart Association. Circulation 2021, 143, e984–e1010. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.J.; Deedwania, P.; Acharya, T.; Aguilar, D.; Bhatt, D.L.; Chyun, D.A.; Di Palo, K.E.; Golden, S.H.; Sperling, L.S.; on behalf of the American Heart Association Diabetes Committee of the Council on Lifestyle and Cardiometabolic Health; et al. Comprehensive Management of Cardiovascular Risk Factors for Adults With Type 2 Diabetes: A Scientific Statement From the American Heart Association. Circulation 2022, 145, e722–e759. [Google Scholar] [CrossRef]
- Ng, A.C.T.; Delgado, V.; Borlaug, B.A.; Bax, J.J. Diabesity: The combined burden of obesity and diabetes on heart disease and the role of imaging. Nat. Rev. Cardiol. 2021, 18, 291–304. [Google Scholar] [CrossRef]
- Amini, M.; Zayeri, F.; Salehi, M. Trend analysis of cardiovascular disease mortality, incidence, and mortality-to-incidence ratio: Results from global burden of disease study 2017. BMC Public Health 2021, 21, 401. [Google Scholar] [CrossRef]
- Sidney, S.; Quesenberry, C.P., Jr.; Jaffe, M.G.; Sorel, M.; Nguyen-Huynh, M.N.; Kushi, L.H.; Go, A.S.; Rana, J.S. Recent Trends in Cardiovascular Mortality in the United States and Public Health Goals. JAMA Cardiol. 2016, 1, 594–599. [Google Scholar] [CrossRef]
- Lloyd-Jones, D.M.; Allen, N.B.; Anderson, C.A.M.; Black, T.; Brewer, L.C.; Foraker, R.E.; Grandner, M.A.; Lavretsky, H.; Perak, A.M.; Sharma, G.; et al. Life’s Essential 8: Updating and Enhancing the American Heart Association’s Construct of Cardiovascular Health: A Presidential Advisory From the American Heart Association. Circulation 2022, 146, e18–e43. [Google Scholar] [CrossRef]
- Diaz, C.L.; Shah, N.S.; Lloyd-Jones, D.M.; Khan, S.S. State of the Nation’s Cardiovascular Health and Targeting Health Equity in the United States: A Narrative Review. JAMA Cardiol. 2021, 6, 963–970. [Google Scholar] [CrossRef]
- Javed, Z.; Haisum Maqsood, M.; Yahya, T.; Amin, Z.; Acquah, I.; Valero-Elizondo, J.; Andrieni, J.; Dubey, P.; Jackson, R.K.; Daffin, M.A.; et al. Race, Racism, and Cardiovascular Health: Applying a Social Determinants of Health Framework to Racial/Ethnic Disparities in Cardiovascular Disease. Circ. Cardiovasc. Qual. Outcomes 2022, 15, e007917. [Google Scholar] [CrossRef] [PubMed]
- Ordovas, J.M.; Smith, C.E. Epigenetics and cardiovascular disease. Nat. Rev. Cardiol. 2010, 7, 510–519. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, S.J.; Tellam, R.L.; Morrison, J.L.; Muhlhausler, B.S.; Molloy, P.L. Recent developments on the role of epigenetics in obesity and metabolic disease. Clin. Epigenet. 2015, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Kazmi, N.; Elliott, H.R.; Burrows, K.; Tillin, T.; Hughes, A.D.; Chaturvedi, N.; Gaunt, T.R.; Relton, C.L. Associations between high blood pressure and DNA methylation. PLoS ONE 2020, 15, e0227728. [Google Scholar] [CrossRef] [PubMed]
- Kou, M.; Li, X.; Shao, X.; Grundberg, E.; Wang, X.; Ma, H.; Heianza, Y.; Martinez, J.A.; Bray, G.A.; Sacks, F.M.; et al. DNA Methylation of Birthweight–Blood Pressure Genes and Changes of Blood Pressure in Response to Weight-Loss Diets in the POUNDS Lost Trial. Hypertension 2023, 80, 1223–1230. [Google Scholar] [CrossRef]
- Lehmann, L.H.; Worst, B.C.; Stanmore, D.A.; Backs, J. Histone deacetylase signaling in cardioprotection. Cell. Mol. Life Sci. 2014, 71, 1673–1690. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, E.A.; Khavjou, O.A.; Thompson, H.; Trogdon, J.G.; Pan, L.; Sherry, B.; Dietz, W. Obesity and severe obesity forecasts through 2030. Am. J. Prev. Med. 2012, 42, 563–570. [Google Scholar] [CrossRef]
- Ward, Z.J.; Bleich, S.N.; Cradock, A.L.; Barrett, J.L.; Giles, C.M.; Flax, C.; Long, M.W.; Gortmaker, S.L. Projected U.S. State-Level Prevalence of Adult Obesity and Severe Obesity. N. Engl. J. Med. 2019, 381, 2440–2450. [Google Scholar] [CrossRef]
- Fryar, C.D.; Carroll, M.D.; Afful, J. Prevalence of Overweight, Obesity, and Severe Obesity among Adults Aged 20 and Over: United States, 1960–1962 through 2017–2018. Available online: https://www.cdc.gov/nchs/data/hestat/obesity_adult_15_16/obesity_adult_15_16.htm (accessed on 16 February 2023).
- Fuster, J.J.; Ouchi, N.; Gokce, N.; Walsh, K. Obesity-Induced Changes in Adipose Tissue Microenvironment and Their Impact on Cardiovascular Disease. Circ. Res. 2016, 118, 1786–1807. [Google Scholar] [CrossRef]
- Koenen, M.; Hill, M.A.; Cohen, P.; Sowers, J.R. Obesity, Adipose Tissue and Vascular Dysfunction. Circ. Res. 2021, 128, 951–968. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Strategies to Prevent Obesity. Available online: https://www.cdc.gov/obesity/strategies/index.html (accessed on 14 October 2019).
- Sadashiv; Modi, A.; Khokhar, M.; Sharma, P.; Joshi, R.; Mishra, S.S.; Bharshankar, R.N.; Tiwari, S.; Singh, P.K.; Bhosale, V.V.; et al. Leptin DNA Methylation and Its Association with Metabolic Risk Factors in a Northwest Indian Obese Population. J. Obes. Metab. Syndr. 2021, 30, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Du, T.; Li, C.; Yang, G. STAT3 phosphorylation in central leptin resistance. Nutr. Metab. 2021, 18, 39. [Google Scholar] [CrossRef] [PubMed]
- Mikula, M.; Majewska, A.; Ledwon, J.K.; Dzwonek, A.; Ostrowski, J. Obesity increases histone H3 lysine 9 and 18 acetylation at Tnfa and Ccl2 genes in mouse liver. Int. J. Mol. Med. 2014, 34, 1647–1654. [Google Scholar] [CrossRef] [PubMed]
- Castillo, E.C.; Morales, J.A.; Chapoy-Villanueva, H.; Silva-Platas, C.; Treviño-Saldaña, N.; Guerrero-Beltrán, C.E.; Bernal-Ramírez, J.; Torres-Quintanilla, A.; García, N.; Youker, K.; et al. Mitochondrial Hyperacetylation in the Failing Hearts of Obese Patients Mediated Partly by a Reduction in SIRT3: The Involvement of the Mitochondrial Permeability Transition Pore. Cell. Physiol. Biochem. 2019, 53, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Romanick, S.S.; Ulrich, C.; Schlauch, K.; Hostler, A.; Payne, J.; Woolsey, R.; Quilici, D.; Feng, Y.; Ferguson, B.S. Obesity-mediated regulation of cardiac protein acetylation: Parallel analysis of total and acetylated proteins via TMT-tagged mass spectrometry. Biosci. Rep. 2018, 38, BSR20180721. [Google Scholar] [CrossRef] [PubMed]
- International Diabete Federation. IDF Diabetes Atlas; IDF: Brussels, Belgium, 2021. [Google Scholar]
- Association, A.D. Economic Costs of Diabetes in the U.S. in 2017. Diabetes Care 2018, 41, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Dal Canto, E.; Ceriello, A.; Rydén, L.; Ferrini, M.; Hansen, T.B.; Schnell, O.; Standl, E.; Beulens, J.W.J. Diabetes as a cardiovascular risk factor: An overview of global trends of macro and micro vascular complications. Eur. J. Prev. Cardiol. 2019, 26, 25–32. [Google Scholar] [CrossRef]
- Nahmias, A.; Stahel, P.; Xiao, C.; Lewis, G.F. Glycemia and Atherosclerotic Cardiovascular Disease: Exploring the Gap Between Risk Marker and Risk Factor. Front. Cardiovasc. Med. 2020, 7, 100. [Google Scholar] [CrossRef]
- Bunyavanich, S.; Do, A.; Vicencio, A. Nasal Gene Expression of Angiotensin-Converting Enzyme 2 in Children and Adults. JAMA 2020, 323, 2427–2429. [Google Scholar] [CrossRef]
- Biswas, S.; Chakrabarti, S. Increased Extracellular Matrix Protein Production in Chronic Diabetic Complications: Implications of Non-Coding RNAs. Noncoding RNA 2019, 5, 30. [Google Scholar] [CrossRef]
- Ma, C.-X.; Ma, X.-N.; Guan, C.-H.; Li, Y.-D.; Mauricio, D.; Fu, S.-B. Cardiovascular disease in type 2 diabetes mellitus: Progress toward personalized management. Cardiovasc. Diabetol. 2022, 21, 74. [Google Scholar] [CrossRef]
- Kyriakos, G.; Quiles-Sanchez, L.V.; Garmpi, A.; Farmaki, P.; Kyre, K.; Savvanis, S.; Antoniou, V.K.; Memi, E. SGLT2 Inhibitors and Cardiovascular Outcomes: Do they Differ or there is a Class Effect? New Insights from the EMPA-REG OUTCOME trial and the CVD-REAL Study. Curr. Cardiol. Rev. 2020, 16, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Raciti, G.A.; Desiderio, A.; Longo, M.; Leone, A.; Zatterale, F.; Prevenzano, I.; Miele, C.; Napoli, R.; Beguinot, F. DNA Methylation and Type 2 Diabetes: Novel Biomarkers for Risk Assessment? Int. J. Mol. Sci. 2021, 22, 11652. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, S.; Natarajan, R. Epigenetics and Type 2 Diabetes Risk. Curr. Diabetes Rep. 2019, 19, 47. [Google Scholar] [CrossRef] [PubMed]
- Baccarelli, A.; Ghosh, S. Environmental exposures, epigenetics and cardiovascular disease. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Khan, A.W.; Akhmedov, A.; Suades, R.; Costantino, S.; Paneni, F.; Caidahl, K.; Mohammed, S.A.; Hage, C.; Gkolfos, C.; et al. Hyperglycemia Induces Myocardial Dysfunction via Epigenetic Regulation of JunD. Circ. Res. 2020, 127, 1261–1273. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Liu, Y.; Luo, M.; Wu, J.; Ma, R.; Wan, Q.; Wu, J. Circulating miRNA-24 and its target YKL-40 as potential biomarkers in patients with coronary heart disease and type 2 diabetes mellitus. Oncotarget 2017, 8, 63038–63046. [Google Scholar] [CrossRef]
- Zhao, T.; Su, Z.; Li, Y.; Zhang, X.; You, Q. Chitinase-3 like-protein-1 function and its role in diseases. Signal Transduct. Target. Ther. 2020, 5, 201. [Google Scholar] [CrossRef]
- Kato, M.; Natarajan, R. Epigenetics and epigenomics in diabetic kidney disease and metabolic memory. Nat. Rev. Nephrol. 2019, 15, 327–345. [Google Scholar] [CrossRef]
- De Rosa, S.; Arcidiacono, B.; Chiefari, E.; Brunetti, A.; Indolfi, C.; Foti, D.P. Type 2 Diabetes Mellitus and Cardiovascular Disease: Genetic and Epigenetic Links. Front. Endocrinol. 2018, 9, 2. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Burden of Cigarette Use in the U.S. Available online: https://www.cdc.gov/tobacco/campaign/tips/resources/data/cigarette-smoking-in-united-states.html (accessed on 17 July 2020).
- Gallucci, G.; Tartarone, A.; Lerose, R.; Lalinga, A.V.; Capobianco, A.M. Cardiovascular risk of smoking and benefits of smoking cessation. J. Thorac. Dis. 2020, 12, 3866–3876. [Google Scholar] [CrossRef] [PubMed]
- Borgerding, M.; Klus, H. Analysis of complex mixtures—Cigarette smoke. Exp. Toxicol. Pathol. 2005, 57 (Suppl. 1), 43–73. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Rawal, I.; Jabbour, S.; Prabhakaran, D. Tobacco and Cardiovascular Disease: A Summary of Evidence. In Cardiovascular, Respiratory, and Related Disorders; Prabhakaran, D., Anand, S., Gaziano, T.A., Mbanya, J.C., Wu, Y., Nugent, R., Eds.; The World Bank: Washington, DC, USA, 2017. [Google Scholar]
- Messner, B.; Bernhard, D. Smoking and cardiovascular disease: Mechanisms of endothelial dysfunction and early atherogenesis. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.D.; Lindson, N.; Hartmann-Boyce, J.; Wahedi, A.; Hajizadeh, A.; Theodoulou, A.; Thomas, E.T.; Lee, C.; Aveyard, P. Smoking cessation for secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2022, 2022, CD014936. [Google Scholar] [CrossRef]
- Power, C.; Atherton, K.; Thomas, C. Maternal smoking in pregnancy, adult adiposity and other risk factors for cardiovascular disease. Atherosclerosis 2010, 211, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Bakker, H.; Jaddoe, V.W. Cardiovascular and metabolic influences of fetal smoke exposure. Eur. J. Epidemiol. 2011, 26, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Ivashkiv, L.B. IFNgamma: Signalling, epigenetics and roles in immunity, metabolism, disease and cancer immunotherapy. Nat. Rev. Immunol. 2018, 18, 545–558. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Zhou, Y.; Shi, H.; Xu, C.; He, H.; Wang, S.; Xiong, X.; Zhang, Y.; Du, Z.; et al. Downregulation of miR-133 via MAPK/ERK signaling pathway involved in nicotine-induced cardiomyocyte apoptosis. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2014, 387, 197–206. [Google Scholar] [CrossRef]
- Fang, F.; Andersen, A.M.; Philibert, R.; Hancock, D.B. Epigenetic biomarkers for smoking cessation. Addict. Neurosci. 2023, 6, 197–206. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Facts About Hypertension. Available online: https://www.cdc.gov/bloodpressure/facts.htm (accessed on 8 September 2020).
- Kirkland, E.B.; Heincelman, M.; Bishu, K.G.; Schumann, S.O.; Schreiner, A.; Axon, R.N.; Mauldin, P.D.; Moran, W.P. Trends in Healthcare Expenditures Among US Adults With Hypertension: National Estimates, 2003–2014. J. Am. Heart Assoc. 2018, 7, e008731. [Google Scholar] [CrossRef]
- Schutte, A.E.; Jafar, T.H.; Poulter, N.R.; Damasceno, A.; Khan, N.A.; Nilsson, P.M.; Alsaid, J.; Neupane, D.; Kario, K.; Beheiry, H.; et al. Addressing global disparities in blood pressure control: Perspectives of the International Society of Hypertension. Cardiovasc. Res. 2023, 119, 381–409. [Google Scholar] [CrossRef] [PubMed]
- Ferdinand, K.C.; Brown, A.L. Will the 2021 USPSTF Hypertension Screening Recommendation Decrease or Worsen Racial/Ethnic Disparities in Blood Pressure Control? JAMA Netw. Open 2021, 4, e213718. [Google Scholar] [CrossRef] [PubMed]
- Stoll, S.; Wang, C.; Qiu, H. DNA Methylation and Histone Modification in Hypertension. Int. J. Mol. Sci. 2018, 19, 1174. [Google Scholar] [CrossRef] [PubMed]
- Helmer, A.; Slater, N.; Smithgall, S. A Review of ACE Inhibitors and ARBs in Black Patients With Hypertension. Ann. Pharmacother. 2018, 52, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E., Jr.; Collins, K.J.; Dennison Himmelfarb, C.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Hypertension 2018, 71, e13–e115. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.I.; Freedman, B.I.; Ellison, D.H.; Rodriguez, C.J. Salt sensitivity: A review with a focus on non-Hispanic blacks and Hispanics. J. Am. Soc. Hypertens. 2013, 7, 170–179. [Google Scholar] [CrossRef] [PubMed]
- van der Linden, E.L.; Halley, A.; Meeks, K.A.C.; Chilunga, F.; Hayfron-Benjamin, C.; Venema, A.; Garrelds, I.M.; Danser, A.H.J.; van den Born, B.-J.; Henneman, P.; et al. An explorative epigenome-wide association study of plasma renin and aldosterone concentration in a Ghanaian population: The RODAM study. Clin. Epigenet. 2022, 14, 159. [Google Scholar] [CrossRef]
- Agha, G.; Mendelson, M.M.; Ward-Caviness, C.K.; Joehanes, R.; Huan, T.; Gondalia, R.; Salfati, E.; Brody, J.A.; Fiorito, G.; Bressler, J.; et al. Blood Leukocyte DNA Methylation Predicts Risk of Future Myocardial Infarction and Coronary Heart Disease. Circulation 2019, 140, 645–657. [Google Scholar] [CrossRef]
- Chitrala, K.N.; Hernandez, D.G.; Nalls, M.A.; Mode, N.A.; Zonderman, A.B.; Ezike, N.; Evans, M.K. Race-specific alterations in DNA methylation among middle-aged African Americans and Whites with metabolic syndrome. Epigenetics 2020, 15, 462–482. [Google Scholar] [CrossRef]
- Surendran, P.; Drenos, F.; Young, R.; Warren, H.; Cook, J.P.; Manning, A.K.; Grarup, N.; Sim, X.; Barnes, D.R.; Witkowska, K.; et al. Trans-ancestry meta-analyses identify rare and common variants associated with blood pressure and hypertension. Nat. Genet. 2016, 48, 1151–1161. [Google Scholar] [CrossRef]
- Liang, M. Epigenetic Mechanisms and Hypertension. Hypertension 2018, 72, 1244–1254. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Yamada, H.; Fujii, R.; Munetsuna, E.; Ando, Y.; Ohashi, K.; Ishikawa, H.; Yamazaki, M.; Maeda, K.; Hashimoto, S.; et al. Association between circulating vascular-related microRNAs and an increase in blood pressure: A 5-year longitudinal population-based study. J. Hypertens. 2021, 39, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, M. Methods of Predicting Preeclampsia Using Biomarkers. U.S. Patent No. 11,344,121, 31 May 2022. [Google Scholar]
- Wu, P.; Haththotuwa, R.; Kwok, C.S.; Babu, A.; Kotronias, R.A.; Rushton, C.; Zaman, A.; Fryer, A.A.; Kadam, U.; Chew-Graham, C.A.; et al. Preeclampsia and Future Cardiovascular Health. Circ. Cardiovasc. Qual. Outcomes 2017, 10, e003497. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.F.; Bordoni, B. Hyperlipidemia. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- CDC. National Hospital Ambulatory Medical Care Survey: 2019 National Summary Tables; CDC: Atlanta, GA, USA, 2023.
- U.S. Preventive Services Task Force. Statin Use for the Primary Prevention of Cardiovascular Disease in Adults: US Preventive Services Task Force Recommendation Statement. JAMA 2022, 328, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Bays, H.E.; Agarwala, A.; German, C.; Satish, P.; Iluyomade, A.; Dudum, R.; Thakkar, A.; Rifai, M.A.; Mehta, A.; Thobani, A.; et al. Ten things to know about ten cardiovascular disease risk factors–2022. Am. J. Prev. Cardiol. 2022, 10, 100342. [Google Scholar] [CrossRef]
- Navar-Boggan, A.M.; Peterson, E.D.; D’Agostino, R.B., Sr.; Neely, B.; Sniderman, A.D.; Pencina, M.J. Hyperlipidemia in early adulthood increases long-term risk of coronary heart disease. Circulation 2015, 131, 451–458. [Google Scholar] [CrossRef]
- Nelson, R.H. Hyperlipidemia as a risk factor for cardiovascular disease. Prim. Care 2013, 40, 195–211. [Google Scholar] [CrossRef]
- Rafieian-Kopaei, M.; Setorki, M.; Doudi, M.; Baradaran, A.; Nasri, H. Atherosclerosis: Process, indicators, risk factors and new hopes. Int. J. Prev. Med. 2014, 5, 927–946. [Google Scholar]
- Greißel, A.; Culmes, M.; Burgkart, R.; Zimmermann, A.; Eckstein, H.-H.; Zernecke, A.; Pelisek, J. Histone acetylation and methylation significantly change with severity of atherosclerosis in human carotid plaques. Cardiovasc. Pathol. 2016, 25, 79–86. [Google Scholar] [CrossRef]
- Zhu, L.; Jia, L.; Liu, N.; Wu, R.; Guan, G.; Hui, R.; Xing, Y.; Zhang, Y.; Wang, J. DNA Methyltransferase 3b Accelerates the Process of Atherosclerosis. Oxid. Med. Cell. Longev. 2022, 2022, 5249367. [Google Scholar] [CrossRef]
- Kuan, R.; Agrawal, D.K.; Thankam, F.G. Treg cells in atherosclerosis. Mol. Biol. Rep. 2021, 48, 4897–4910. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Atkinson, C.; Bhalla, K.; Birbeck, G.; Burstein, R.; Chou, D.; Dellavalle, R.; Danaei, G.; Ezzati, M.; Fahimi, A.; et al. The state of US health, 1990-2010: Burden of diseases, injuries, and risk factors. JAMA 2013, 310, 591–608. [Google Scholar] [CrossRef] [PubMed]
- Pahwa, R.; Jialal, I. Atherosclerosis. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- Wong, N.D.; Budoff, M.J.; Ferdinand, K.; Graham, I.M.; Michos, E.D.; Reddy, T.; Shapiro, M.D.; Toth, P.P. Atherosclerotic cardiovascular disease risk assessment: An American Society for Preventive Cardiology clinical practice statement. Am. J. Prev. Cardiol. 2022, 10, 100335. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.M.; Wilson, P.W.; Odell, P.M.; Kannel, W.B. An updated coronary risk profile. A statement for health professionals. Circulation 1991, 83, 356–362. [Google Scholar] [CrossRef]
- Okunrintemi, V.; Valero-Elizondo, J.; Patrick, B.; Salami, J.; Tibuakuu, M.; Ahmad, S.; Ogunmoroti, O.; Mahajan, S.; Khan, S.U.; Gulati, M.; et al. Gender Differences in Patient-Reported Outcomes Among Adults With Atherosclerotic Cardiovascular Disease. J. Am. Heart Assoc. 2018, 7, e010498. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.T.; Mahtta, D.; Ramsey, D.J.; Liu, J.; Misra, A.; Nasir, K.; Samad, Z.; Itchhaporia, D.; Khan, S.U.; Schofield, R.S.; et al. Sex-Related Disparities in Cardiovascular Health Care Among Patients With Premature Atherosclerotic Cardiovascular Disease. JAMA Cardiol. 2021, 6, 782–790. [Google Scholar] [CrossRef]
- Freaney, P.M.; Khan, S.S.; Lloyd-Jones, D.M.; Stone, N.J. The Role of Sex-Specific Risk Factors in the Risk Assessment of Atherosclerotic Cardiovascular Disease for Primary Prevention in Women. Curr. Atheroscler. Rep. 2020, 22, 46. [Google Scholar] [CrossRef]
- Rämö, J.T.; Ripatti, P.; Tabassum, R.; Söderlund, S.; Matikainen, N.; Gerl, M.J.; Klose, C.; Surma, M.A.; Stitziel, N.O.; Havulinna, A.S.; et al. Coronary Artery Disease Risk and Lipidomic Profiles Are Similar in Hyperlipidemias With Family History and Population-Ascertained Hyperlipidemias. J. Am. Heart Assoc. 2019, 8, e012415. [Google Scholar] [CrossRef]
- Mihăilă, R.G. Pragmatic Analysis of Dyslipidemia Involvement in Coronary Artery Disease: A Narrative Review. Curr. Cardiol. Rev. 2020, 16, 36–47. [Google Scholar] [CrossRef]
- Khera, A.V.; Kathiresan, S. Genetics of coronary artery disease: Discovery, biology and clinical translation. Nat. Rev. Genet. 2017, 18, 331–344. [Google Scholar] [CrossRef]
- Kessler, T.; Schunkert, H. Coronary Artery Disease Genetics Enlightened by Genome-Wide Association Studies. JACC Basic Transl. Sci. 2021, 6, 610–623. [Google Scholar] [CrossRef]
- Poznyak, A.V.; Grechko, A.V.; Wetzker, R.; Orekhov, A.N. In Search for Genes Related to Atherosclerosis and Dyslipidemia Using Animal Models. Int. J. Mol. Sci. 2020, 21, 2097. [Google Scholar] [CrossRef] [PubMed]
- Maneerat, Y.; Prasongsukarn, K.; Benjathummarak, S.; Dechkhajorn, W. PPBP and DEFA1/DEFA3 genes in hyperlipidaemia as feasible synergistic inflammatory biomarkers for coronary heart disease. Lipids Health Dis. 2017, 16, 80. [Google Scholar] [CrossRef] [PubMed]
- Eberharter, A.; Becker, P.B. Histone acetylation: A switch between repressive and permissive chromatin. Second in review series on chromatin dynamics. EMBO Rep. 2002, 3, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.M.; Johnson, A.; Tarapore, P.; Janakiram, V.; Zhang, X.; Leung, Y.K. Environmental epigenetics and its implication on disease risk and health outcomes. Ilar J. 2012, 53, 289–305. [Google Scholar] [CrossRef]
- Mazzio, E.A.; Soliman, K.F. Basic concepts of epigenetics: Impact of environmental signals on gene expression. Epigenetics 2012, 7, 119–130. [Google Scholar] [CrossRef]
- Rendina González, A.P.; Preite, V.; Verhoeven, K.J.F.; Latzel, V. Transgenerational Effects and Epigenetic Memory in the Clonal Plant Trifolium repens. Front. Plant Sci. 2018, 9, 1677. [Google Scholar] [CrossRef]
- de Castro Barbosa, T.; Ingerslev, L.R.; Alm, P.S.; Versteyhe, S.; Massart, J.; Rasmussen, M.; Donkin, I.; Sjögren, R.; Mudry, J.M.; Vetterli, L.; et al. High-fat diet reprograms the epigenome of rat spermatozoa and transgenerationally affects metabolism of the offspring. Mol. Metab. 2016, 5, 184–197. [Google Scholar] [CrossRef]
- Heijmans, B.T.; Tobi, E.W.; Stein, A.D.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.E.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar] [CrossRef]
- Bleker, L.S.; de Rooij, S.R.; Painter, R.C.; Ravelli, A.C.; Roseboom, T.J. Cohort profile: The Dutch famine birth cohort (DFBC)- a prospective birth cohort study in the Netherlands. BMJ Open 2021, 11, e042078. [Google Scholar] [CrossRef]
- Abraham, M.J.; El Sherbini, A.; El-Diasty, M.; Askari, S.; Szewczuk, M.R. Restoring Epigenetic Reprogramming with Diet and Exercise to Improve Health-Related Metabolic Diseases. Biomolecules 2023, 13, 318. [Google Scholar] [CrossRef] [PubMed]
- Almodóvar, A.S.; Nahata, M.C. Associations Between Chronic Disease, Polypharmacy, and Medication-Related Problems Among Medicare Beneficiaries. J. Manag. Care Spec. Pharm. 2019, 25, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Sheikh-Taha, M.; Asmar, M. Polypharmacy and severe potential drug-drug interactions among older adults with cardiovascular disease in the United States. BMC Geriatr. 2021, 21, 233. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Liu, C.F.; Feng, Y.F.; Chen, H. Potential drug-drug interactions in drug therapy for older adults with chronic coronary syndrome at hospital discharge: A real-world study. Front. Pharmacol. 2022, 13, 946415. [Google Scholar] [CrossRef] [PubMed]
- Laslett, D.B.; Cooper, J.M.; Greenberg, R.M.; Yesenosky, G.A.; Basil, A.; Gangireddy, C.; Whitman, I.R. Electrolyte Abnormalities in Patients Presenting With Ventricular Arrhythmia (from the LYTE-VT Study). Am. J. Cardiol. 2020, 129, 36–41. [Google Scholar] [CrossRef]
- Skogestad, J.; Aronsen, J.M. Hypokalemia-Induced Arrhythmias and Heart Failure: New Insights and Implications for Therapy. Front. Physiol. 2018, 9, 1500. [Google Scholar] [CrossRef] [PubMed]
- van der Laan, D.M.; Elders, P.J.M.; Boons, C.; Nijpels, G.; Krska, J.; Hugtenburg, J.G. The impact of cardiovascular medication use on patients’ daily lives: A cross-sectional study. Int. J. Clin. Pharm. 2018, 40, 412–420. [Google Scholar] [CrossRef]
- Leslie, K.H.; McCowan, C.; Pell, J.P. Adherence to cardiovascular medication: A review of systematic reviews. J. Public Health 2019, 41, e84–e94. [Google Scholar] [CrossRef]
- Bansal, A.B.; Cassagnol, M. HMG-CoA Reductase Inhibitors. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- Diamantis, E.; Kyriakos, G.; Quiles-Sanchez, L.V.; Farmaki, P.; Troupis, T. The Anti-Inflammatory Effects of Statins on Coronary Artery Disease: An Updated Review of the Literature. Curr. Cardiol. Rev. 2017, 13, 209–216. [Google Scholar] [CrossRef]
- Arnett, D.K.; Blumenthal, R.S.; Albert, M.A.; Buroker, A.B.; Goldberger, Z.D.; Hahn, E.J.; Himmelfarb, C.D.; Khera, A.; Lloyd-Jones, D.; McEvoy, J.W.; et al. 2019 ACC/AHA Guideline on the Primary Prevention of Cardiovascular Disease: Executive Summary: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2019, 140, e563–e595. [Google Scholar] [CrossRef]
- Tarn, D.M.; Pletcher, M.J.; Tosqui, R.; Fernandez, A.; Tseng, C.-h.; Moriconi, R.; Bell, D.S.; Barrientos, M.; Turner, J.A.; Schwartz, J.B. Primary nonadherence to statin medications: Survey of patient perspectives. Prev. Med. Rep. 2021, 22, 101357. [Google Scholar] [CrossRef] [PubMed]
- Lavie, G.; Hoshen, M.; Leibowitz, M.; Benis, A.; Akriv, A.; Balicer, R.; Reges, O. Statin Therapy for Primary Prevention in the Elderly and Its Association with New-Onset Diabetes, Cardiovascular Events, and All-Cause Mortality. Am. J. Med. 2021, 134, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Tangelloju, S.; Little, B.B.; Esterhay, R.J.; Brock, G.; LaJoie, S. Statins are associated with new onset type 2 diabetes mellitus (T2DM) in Medicare patients ≥65 years. Diabetes Metab. Res. Rev. 2020, 36, e3310. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Sikarwar, P.; Khurana, S.; Sharma, J. Assessing the Incidence of New-onset Diabetes Mellitus with Statin Use: A Systematic Review of the Systematic Reviews and Meta-analyses. Touchrev Endocrinol. 2022, 18, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Mollazadeh, H.; Tavana, E.; Fanni, G.; Bo, S.; Banach, M.; Pirro, M.; von Haehling, S.; Jamialahmadi, T.; Sahebkar, A. Effects of statins on mitochondrial pathways. J. Cachexia Sarcopenia Muscle 2021, 12, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Tricarico, P.M.; Crovella, S.; Celsi, F. Mevalonate Pathway Blockade, Mitochondrial Dysfunction and Autophagy: A Possible Link. Int. J. Mol. Sci. 2015, 16, 16067–16084. [Google Scholar] [CrossRef]
- Galicia-Garcia, U.; Jebari, S.; Larrea-Sebal, A.; Uribe, K.B.; Siddiqi, H.; Ostolaza, H.; Benito-Vicente, A.; Martín, C. Statin Treatment-Induced Development of Type 2 Diabetes: From Clinical Evidence to Mechanistic Insights. Int. J. Mol. Sci. 2020, 21, 4725. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Lin, J.-H.; Chou, C.-W.; Chang, Y.-F.; Yeh, S.-H.; Chen, C.-C. Statins Increase p21 through Inhibition of Histone Deacetylase Activity and Release of Promoter-Associated HDAC1/2. Cancer Res. 2008, 68, 2375–2383. [Google Scholar] [CrossRef]
- Yoon, S.; Eom, G.H. HDAC and HDAC Inhibitor: From Cancer to Cardiovascular Diseases. Chonnam Med. J. 2016, 52, 1–11. [Google Scholar] [CrossRef]
- Gillette, T.G. HDAC Inhibition in the Heart. Circulation 2021, 143, 1891–1893. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, Y.T.; Zhao, T.C. Histone deacetylases in modulating cardiac disease and their clinical translational and therapeutic implications. Exp. Biol. Med. 2021, 246, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Andolina, D.; Savi, M.; Ielpo, D.; Barbetti, M.; Bocchi, L.; Stilli, D.; Ventura, R.; Lo Iacono, L.; Sgoifo, A.; Carnevali, L. Elevated miR-34a expression and altered transcriptional profile are associated with adverse electromechanical remodeling in the heart of male rats exposed to social stress. Stress 2021, 24, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Qu, G.; Han, C.; Wang, Y.; Sun, T.; Li, F.; Wang, J.; Luo, S. MiR-34a, miR-21 and miR-23a as potential biomarkers for coronary artery disease: A pilot microarray study and confirmation in a 32 patient cohort. Exp. Mol. Med. 2015, 47, e138. [Google Scholar] [CrossRef] [PubMed]
- Gatsiou, A.; Georgiopoulos, G.; Vlachogiannis, N.I.; Pfisterer, L.; Fischer, A.; Sachse, M.; Laina, A.; Bonini, F.; Delialis, D.; Tual-Chalot, S.; et al. Additive contribution of microRNA-34a/b/c to human arterial ageing and atherosclerosis. Atherosclerosis 2021, 327, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Hua, C.C.; Liu, X.M.; Liang, L.R.; Wang, L.F.; Zhong, J.C. Targeting the microRNA-34a as a Novel Therapeutic Strategy for Cardiovascular Diseases. Front. Cardiovasc. Med. 2021, 8, 784044. [Google Scholar] [CrossRef] [PubMed]
- Raucci, A.; Macrì, F.; Castiglione, S.; Badi, I.; Vinci, M.C.; Zuccolo, E. MicroRNA-34a: The bad guy in age-related vascular diseases. Cell. Mol. Life Sci. 2021, 78, 7355–7378. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, T.; Satoh, M.; Itoh, T.; Nakamura, M. MicroRNA-34a regulates the longevity-associated protein SIRT1 in coronary artery disease: Effect of statins on SIRT1 and microRNA-34a expression. Clin. Sci. 2012, 123, 161–171. [Google Scholar] [CrossRef]
- Karlic, H.; Thaler, R.; Gerner, C.; Grunt, T.; Proestling, K.; Haider, F.; Varga, F. Inhibition of the mevalonate pathway affects epigenetic regulation in cancer cells. Cancer Genet. 2015, 208, 241–252. [Google Scholar] [CrossRef]
- Weber, M.A.; Schiffrin, E.L.; White, W.B.; Mann, S.; Lindholm, L.H.; Kenerson, J.G.; Flack, J.M.; Carter, B.L.; Materson, B.J.; Ram, C.V.; et al. Clinical practice guidelines for the management of hypertension in the community: A statement by the American Society of Hypertension and the International Society of Hypertension. J. Clin. Hypertens. 2014, 16, 14–26. [Google Scholar] [CrossRef]
- Basile, J. The role of existing and newer calcium channel blockers in the treatment of hypertension. J. Clin. Hypertens. 2004, 6, 621–629, quiz 630–621. [Google Scholar] [CrossRef]
- McKeever, R.G.; Hamilton, R.J. Calcium Channel Blockers. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- Godfraind, T. Calcium Channel Blockers in Cardiovascular Pharmacotherapy. J. Cardiovasc. Pharmacol. Ther. 2014, 19, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Sueta, D.; Tabata, N.; Hokimoto, S. Clinical roles of calcium channel blockers in ischemic heart diseases. Hypertens. Res. 2017, 40, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Aronow, W.S. Medical management after myocardial infarction. Future Cardiol. 2019, 15, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, P.A.; Bozkurt, B.; Aguilar, D.; Allen, L.A.; Byun, J.J.; Colvin, M.M.; Deswal, A.; Drazner, M.H.; Dunlay, S.M.; Evers, L.R.; et al. 2022 AHA/ACC/HFSA Guideline for the Management of Heart Failure: A Report of the American College of Cardiology/American Heart Association Joint Committee on Clinical Practice Guidelines. Circulation 2022, 145, e895–e1032. [Google Scholar] [CrossRef] [PubMed]
- Palandri, C.; Santini, L.; Argirò, A.; Margara, F.; Doste, R.; Bueno-Orovio, A.; Olivotto, I.; Coppini, R. Pharmacological Management of Hypertrophic Cardiomyopathy: From Bench to Bedside. Drugs 2022, 82, 889–912. [Google Scholar] [CrossRef] [PubMed]
- Leitch, J.W.; McElduff, P.; Dobson, A.; Heller, R. Outcome With Calcium Channel Antagonists After Myocardial Infarction: A Community-Based Study. J. Am. Coll. Cardiol. 1998, 31, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.A.; Wang, X.; Garcia-Betancourt, M.M.; Forse, R.A. Calcium channel blockers and the incidence of breast and prostate cancer: A meta-analysis. J. Clin. Pharm. Ther. 2018, 43, 519–529. [Google Scholar] [CrossRef]
- Saltzman, B.S.; Weiss, N.S.; Sieh, W.; Fitzpatrick, A.L.; McTiernan, A.; Daling, J.R.; Li, C.I. Use of antihypertensive medications and breast cancer risk. Cancer Causes Control 2013, 24, 365–371. [Google Scholar] [CrossRef]
- Rotshild, V.; Hirsh Raccah, B.; Gazawe, M.; Matok, I. Calcium Channel Blocker Use and the Risk for Breast Cancer: A Population-Based Nested Case-Control Study. Cancers 2022, 14, 2344. [Google Scholar] [CrossRef]
- Kizer, J.R.; Kimmel, S.E. Epidemiologic review of the calcium channel blocker drugs. An up-to-date perspective on the proposed hazards. Arch. Intern. Med. 2001, 161, 1145–1158. [Google Scholar] [CrossRef]
- Elliott, W.J.; Ram, C.V.S. Calcium Channel Blockers. J. Clin. Hypertens. 2011, 13, 687–689. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Li, X.; Wang, Y.; Sjölander, A.; Johnell, K.; Thambisetty, M.; Ferrucci, L.; Reynolds, C.A.; Finkel, D.; Jylhävä, J.; et al. Longitudinal associations between use of antihypertensive, antidiabetic, and lipid-lowering medications and biological aging. GeroScience 2023, 45, 2065–2078. [Google Scholar] [CrossRef] [PubMed]
- Kho, M.; Wang, Y.Z.; Chaar, D.; Zhao, W.; Ratliff, S.M.; Mosley, T.H.; Peyser, P.A.; Kardia, S.L.R.; Smith, J.A. Accelerated DNA methylation age and medication use among African Americans. Aging 2021, 13, 14604–14629. [Google Scholar] [CrossRef]
- Dai, H.; Sinclair, D.A.; Ellis, J.L.; Steegborn, C. Sirtuin activators and inhibitors: Promises, achievements, and challenges. Pharmacol. Ther. 2018, 188, 140–154. [Google Scholar] [CrossRef] [PubMed]
- Manna, D.; Bhuyan, R.; Ghosh, R. Probing the mechanism of SIRT1 activation by a 1,4-dihydropyridine. J. Mol. Model. 2018, 24, 340. [Google Scholar] [CrossRef] [PubMed]
- Zwergel, C.; Aventaggiato, M.; Garbo, S.; Di Bello, E.; Fassari, B.; Noce, B.; Castiello, C.; Lambona, C.; Barreca, F.; Rotili, D.; et al. Novel 1,4-Dihydropyridines as Specific Binders and Activators of SIRT3 Impair Cell Viability and Clonogenicity and Downregulate Hypoxia-Induced Targets in Cancer Cells. J. Med. Chem. 2023, 66, 9622–9641. [Google Scholar] [CrossRef]
- Holley, A.K.; Bakthavatchalu, V.; Velez-Roman, J.M.; St Clair, D.K. Manganese superoxide dismutase: Guardian of the powerhouse. Int. J. Mol. Sci. 2011, 12, 7114–7162. [Google Scholar] [CrossRef]
- Kane, A.E.; Sinclair, D.A. Sirtuins and NAD+ in the Development and Treatment of Metabolic and Cardiovascular Diseases. Circ. Res. 2018, 123, 868–885. [Google Scholar] [CrossRef]
- Matsushima, S.; Sadoshima, J. The role of sirtuins in cardiac disease. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1375–H1389. [Google Scholar] [CrossRef]
- Farzam, K.; Jan, A. Beta Blockers. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- Gorre, F.; Vandekerckhove, H. Beta-blockers: Focus on mechanism of action Which beta-blocker, when and why? Acta Cardiol. 2010, 65, 565–570. [Google Scholar] [CrossRef]
- Schena, G.; Caplan, M.J. Everything You Always Wanted to Know about β(3)-AR * (* But Were Afraid to Ask). Cells 2019, 8, 357. [Google Scholar] [CrossRef] [PubMed]
- Divan, I.; Suleman, A.; Lars, L.; Joakim, A.; Tatendashe Bernadette, D.; Claes, H.; Tomas, J.; Troels, Y.; Chris, P.G.; Gorav, B. Association of beta-blockers beyond 1 year after myocardial infarction and cardiovascular outcomes. Heart 2023, 109, 1159. [Google Scholar] [CrossRef]
- Godoy, L.C.; Farkouh, M.E.; Austin, P.C.; Shah, B.R.; Qiu, F.; Jackevicius, C.A.; Wijeysundera, H.C.; Krumholz, H.M.; Ko, D.T. Association of Beta-Blocker Therapy With Cardiovascular Outcomes in Patients With Stable Ischemic Heart Disease. J. Am. Coll. Cardiol. 2023, 81, 2299–2311. [Google Scholar] [CrossRef] [PubMed]
- Bangalore, S.; Steg, G.; Deedwania, P.; Crowley, K.; Eagle, K.A.; Goto, S.; Ohman, E.M.; Cannon, C.P.; Smith, S.C.; Zeymer, U.; et al. β-Blocker Use and Clinical Outcomes in Stable Outpatients With and Without Coronary Artery Disease. JAMA 2012, 308, 1340–1349. [Google Scholar] [CrossRef] [PubMed]
- Ziff, O.J.; Samra, M.; Howard, J.P.; Bromage, D.I.; Ruschitzka, F.; Francis, D.P.; Kotecha, D. Beta-blocker efficacy across different cardiovascular indications: An umbrella review and meta-analytic assessment. BMC Med. 2020, 18, 103. [Google Scholar] [CrossRef]
- Dézsi, C.A.; Szentes, V. The Real Role of β-Blockers in Daily Cardiovascular Therapy. Am. J. Cardiovasc. Drugs 2017, 17, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Barron, A.J.; Zaman, N.; Cole, G.D.; Wensel, R.; Okonko, D.O.; Francis, D.P. Systematic review of genuine versus spurious side-effects of beta-blockers in heart failure using placebo control: Recommendations for patient information. Int. J. Cardiol. 2013, 168, 3572–3579. [Google Scholar] [CrossRef]
- Lin, W.; Huang, Z.; Ping, S.; Zhang, S.; Wen, X.; He, Y.; Ren, Y. Toxicological effects of atenolol and venlafaxine on zebrafish tissues: Bioaccumulation, DNA hypomethylation, and molecular mechanism. Environ. Pollut. 2022, 299, 118898. [Google Scholar] [CrossRef]
- Napoli, C.; Grimaldi, V.; De Pascale, M.R.; Sommese, L.; Infante, T.; Soricelli, A. Novel epigenetic-based therapies useful in cardiovascular medicine. World J. Cardiol. 2016, 8, 211–219. [Google Scholar] [CrossRef]
- Zhong, J.; Agha, G.; Baccarelli, A.A. The Role of DNA Methylation in Cardiovascular Risk and Disease: Methodological Aspects, Study Design, and Data Analysis for Epidemiological Studies. Circ. Res. 2016, 118, 119–131. [Google Scholar] [CrossRef]
- Solayman, M.H.; Langaee, T.Y.; Gong, Y.; Shahin, M.H.; Turner, S.T.; Chapman, A.B.; Gums, J.G.; Boerwinkle, E.; Beitelshees, A.L.; El-Hamamsy, M.; et al. Effect of plasma MicroRNA on antihypertensive response to beta blockers in the Pharmacogenomic Evaluation of Antihypertensive Responses (PEAR) studies. Eur. J. Pharm. Sci. 2019, 131, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Kovell, L.C.; Ahmed, H.M.; Misra, S.; Whelton, S.P.; Prokopowicz, G.P.; Blumenthal, R.S.; McEvoy, J.W. US Hypertension Management Guidelines: A Review of the Recent Past and Recommendations for the Future. J. Am. Heart Assoc. 2015, 4, e002315. [Google Scholar] [CrossRef] [PubMed]
- Tai, C.; Gan, T.; Zou, L.; Sun, Y.; Zhang, Y.; Chen, W.; Li, J.; Zhang, J.; Xu, Y.; Lu, H.; et al. Effect of angiotensin-converting enzyme inhibitors and angiotensin II receptor blockers on cardiovascular events in patients with heart failure: A meta-analysis of randomized controlled trials. BMC Cardiovasc. Disord. 2017, 17, 257. [Google Scholar] [CrossRef] [PubMed]
- Messerli Franz, H.; Bangalore, S.; Bavishi, C.; Rimoldi Stefano, F. Angiotensin-Converting Enzyme Inhibitors in Hypertension. J. Am. Coll. Cardiol. 2018, 71, 1474–1482. [Google Scholar] [CrossRef] [PubMed]
- Awad, K.; Zaki, M.M.; Mohammed, M.; Lewek, J.; Lavie, C.J.; Banach, M. Effect of the Renin-Angiotensin System Inhibitors on Inflammatory Markers: A Systematic Review and Meta-analysis of Randomized Controlled Trials. Mayo Clin. Proc. 2022, 97, 1808–1823. [Google Scholar] [CrossRef] [PubMed]
- Herman, L.L.; Padala, S.A.; Ahmed, I.; Bashir, K. Angiotensin-Converting Enzyme Inhibitors (ACEI). In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- Yılmaz, İ. Angiotensin-Converting Enzyme Inhibitors Induce Cough. Turk. Thorac. J. 2019, 20, 36–42. [Google Scholar] [CrossRef]
- Pinto, B.; Jadhav, U.; Singhai, P.; Sadhanandham, S.; Shah, N. ACEI-induced cough: A review of current evidence and its practical implications for optimal CV risk reduction. Indian Heart J. 2020, 72, 345–350. [Google Scholar] [CrossRef]
- Mirabito Colafella, K.M.; Uijl, E.; Jan Danser, A.H. Interference with the Renin–Angiotensin System (RAS): Classical Inhibitors and Novel Approaches. In Encyclopedia of Endocrine Diseases, 2nd ed.; Huhtaniemi, I., Martini, L., Eds.; Academic Press: Oxford, UK, 2019; pp. 523–530. [Google Scholar]
- Burnier, M. Angiotensin II Type 1 Receptor Blockers. Circulation 2001, 103, 904–912. [Google Scholar] [CrossRef]
- Barreras, A.; Gurk-Turner, C. Angiotensin II receptor blockers. Proc. (Bayl. Univ. Med. Cent.) 2003, 16, 123–126. [Google Scholar] [CrossRef]
- Reddy, M.A.; Sumanth, P.; Lanting, L.; Yuan, H.; Wang, M.; Mar, D.; Alpers, C.E.; Bomsztyk, K.; Natarajan, R. Losartan reverses permissive epigenetic changes in renal glomeruli of diabetic db/db mice. Kidney Int. 2014, 85, 362–373. [Google Scholar] [CrossRef]
- Wang, T.; Lian, G.; Cai, X.; Lin, Z.; Xie, L. Effect of prehypertensive losartan therapy on AT1R and ATRAP methylation of adipose tissue in the later life of high-fat-fed spontaneously hypertensive rats. Mol. Med. Rep. 2018, 17, 1753–1761. [Google Scholar] [CrossRef]
- Seto, E.; Yoshida, M. Erasers of histone acetylation: The histone deacetylase enzymes. Cold Spring Harb. Perspect. Biol. 2014, 6, a018713. [Google Scholar] [CrossRef]
- Park, S.-Y.; Kim, J.-S. A short guide to histone deacetylases including recent progress on class II enzymes. Exp. Mol. Med. 2020, 52, 204–212. [Google Scholar] [CrossRef]
- Li, G.; Tian, Y.; Zhu, W.-G. The Roles of Histone Deacetylases and Their Inhibitors in Cancer Therapy. Front. Cell Dev. Biol. 2020, 8, 576946. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, R.A.; Weeks, K.L. Histone deacetylases in cardiovascular and metabolic diseases. J. Mol. Cell. Cardiol. 2019, 130, 151–159. [Google Scholar] [CrossRef]
- Lee, H.T.; Oh, S.; Ro, D.H.; Yoo, H.; Kwon, Y.W. The Key Role of DNA Methylation and Histone Acetylation in Epigenetics of Atherosclerosis. J. Lipid Atheroscler. 2020, 9, 419–434. [Google Scholar] [CrossRef] [PubMed]
- Kronlage, M.; Katus, H.A.; Backs, J. HDAC Signaling Networks in Heart Failure. In Epigenetics in Cardiac Disease; Backs, J., McKinsey, T.A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 191–208. [Google Scholar]
- Kabra, D.G.; Pfuhlmann, K.; García-Cáceres, C.; Schriever, S.C.; Casquero García, V.; Kebede, A.F.; Fuente-Martin, E.; Trivedi, C.; Heppner, K.; Uhlenhaut, N.H.; et al. Hypothalamic leptin action is mediated by histone deacetylase 5. Nat. Commun. 2016, 7, 10782. [Google Scholar] [CrossRef] [PubMed]
- Çakır, I.; Hadley, C.K.; Pan, P.L.; Bagchi, R.A.; Ghamari-Langroudi, M.; Porter, D.T.; Wang, Q.; Litt, M.J.; Jana, S.; Hagen, S.; et al. Histone deacetylase 6 inhibition restores leptin sensitivity and reduces obesity. Nat. Metab. 2022, 4, 44–59. [Google Scholar] [CrossRef]
- Chen, Y.; Du, J.; Zhao, Y.T.; Zhang, L.; Lv, G.; Zhuang, S.; Qin, G.; Zhao, T.C. Histone deacetylase (HDAC) inhibition improves myocardial function and prevents cardiac remodeling in diabetic mice. Cardiovasc. Diabetol. 2015, 14, 99. [Google Scholar] [CrossRef]
- Jung, J.K.; Yoon, G.-E.; Jang, G.; Park, K.M.; Kim, I.; Kim, J.I. Inhibition of HDACs (Histone Deacetylases) Ameliorates High-Fat Diet–Induced Hypertension Through Restoration of the MsrA (Methionine Sulfoxide Reductase A)/Hydrogen Sulfide Axis. Hypertension 2021, 78, 1103–1115. [Google Scholar] [CrossRef]
- Rahman, M.; Awosika, A.O.; Nguyen, H. Valproic Acid. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- Göttlicher, M.; Minucci, S.; Zhu, P.; Krämer, O.H.; Schimpf, A.; Giavara, S.; Sleeman, J.P.; Lo Coco, F.; Nervi, C.; Pelicci, P.G.; et al. Valproic acid defines a novel class of HDAC inhibitors inducing differentiation of transformed cells. Embo J. 2001, 20, 6969–6978. [Google Scholar] [CrossRef] [PubMed]
- Sixto-López, Y.; Bello, M.; Correa-Basurto, J. Exploring the inhibitory activity of valproic acid against the HDAC family using an MMGBSA approach. J. Comput.-Aided Mol. Des. 2020, 34, 857–878. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-A.; Lee, D.-Y.; Cho, H.-M.; Kim, S.-Y.; Iwasaki, Y.; Kim, I.K. Histone Deacetylase Inhibition Attenuates Transcriptional Activity of Mineralocorticoid Receptor Through Its Acetylation and Prevents Development of Hypertension. Circ. Res. 2013, 112, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Terker, A.S.; Ellison, D.H. Renal mineralocorticoid receptor and electrolyte homeostasis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R1068–R1070. [Google Scholar] [CrossRef]
- Scholz, B.; Schulte, J.S.; Hamer, S.; Himmler, K.; Pluteanu, F.; Seidl, M.D.; Stein, J.; Wardelmann, E.; Hammer, E.; Völker, U.; et al. HDAC (Histone Deacetylase) Inhibitor Valproic Acid Attenuates Atrial Remodeling and Delays the Onset of Atrial Fibrillation in Mice. Circ. Arrhythmia Electrophysiol. 2019, 12, e007071. [Google Scholar] [CrossRef] [PubMed]
- Odutayo, A.; Wong, C.X.; Hsiao, A.J.; Hopewell, S.; Altman, D.G.; Emdin, C.A. Atrial fibrillation and risks of cardiovascular disease, renal disease, and death: Systematic review and meta-analysis. BMJ 2016, 354, i4482. [Google Scholar] [CrossRef]
- Tian, S.; Lei, I.; Gao, W.; Liu, L.; Guo, Y.; Creech, J.; Herron, T.J.; Xian, S.; Ma, P.X.; Eugene Chen, Y.; et al. HDAC inhibitor valproic acid protects heart function through Foxm1 pathway after acute myocardial infarction. EBioMedicine 2019, 39, 83–94. [Google Scholar] [CrossRef]
- English, J.D.; Tian, S.; Wang, Z.; Luzum, J.A. Association of Valproic Acid Use With Post-Myocardial Infarction Heart Failure Development: A Meta-Analysis of Two Retrospective Case-Control Studies. J. Cardiovasc. Pharmacol. Ther. 2022, 27, 10742484221140303. [Google Scholar] [CrossRef]
- Kusaczuk, M.; Krętowski, R.; Bartoszewicz, M.; Cechowska-Pasko, M. Phenylbutyrate-a pan-HDAC inhibitor-suppresses proliferation of glioblastoma LN-229 cell line. Tumour Biol. 2016, 37, 931–942. [Google Scholar] [CrossRef]
- Daosukho, C.; Chen, Y.; Noel, T.; Sompol, P.; Nithipongvanitch, R.; Velez, J.M.; Oberley, T.D.; St Clair, D.K. Phenylbutyrate, a histone deacetylase inhibitor, protects against Adriamycin-induced cardiac injury. Free Radic. Biol. Med. 2007, 42, 1818–1825. [Google Scholar] [CrossRef]
- Takatori, O.; Usui, S.; Okajima, M.; Kaneko, S.; Ootsuji, H.; Takashima, S.-i.; Kobayashi, D.; Murai, H.; Furusho, H.; Takamura, M. Sodium 4-Phenylbutyrate Attenuates Myocardial Reperfusion Injury by Reducing the Unfolded Protein Response. J. Cardiovasc. Pharmacol. Ther. 2016, 22, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Adi, D.; Long, M.; Wang, J.; Liu, F.; Gai, M.T.; Aierken, A.; Li, M.Y.; Li, Q.; Wu, L.Q.; et al. 4-Phenylbutyric Acid Induces Protection against Pulmonary Arterial Hypertension in Rats. PLoS ONE 2016, 11, e0157538. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Luo, T.; Zeng, Z.; Fu, H.; Asano, Y.; Liao, Y.; Minamino, T.; Kitakaze, M. Histone Deacetylase Inhibitor Phenylbutyrate Exaggerates Heart Failure in Pressure Overloaded Mice independently of HDAC inhibition. Sci. Rep. 2016, 6, 34036. [Google Scholar] [CrossRef] [PubMed]
- Bubna, A.K. Vorinostat-An Overview. Indian J. Dermatol. 2015, 60, 419. [Google Scholar] [CrossRef] [PubMed]
- Pedro Ferreira, J.; Pitt, B.; Zannad, F. Histone deacetylase inhibitors for cardiovascular conditions and healthy longevity. Lancet Healthy Longev. 2021, 2, e371–e379. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Kong, Y.; Tan, W.; May, H.; Battiprolu, P.K.; Pedrozo, Z.; Wang, Z.V.; Morales, C.; Luo, X.; Cho, G.; et al. Histone deacetylase inhibition blunts ischemia/reperfusion injury by inducing cardiomyocyte autophagy. Circulation 2014, 129, 1139–1151. [Google Scholar] [CrossRef]
- Nagata, S.; Marunouchi, T.; Tanonaka, K. Histone Deacetylase Inhibitor SAHA Treatment Prevents the Development of Heart Failure after Myocardial Infarction via an Induction of Heat-Shock Proteins in Rats. Biol. Pharm. Bull. 2019, 42, 453–461. [Google Scholar] [CrossRef]
- Patnaik, S.; Nathan, S.; Kar, B.; Gregoric, I.D.; Li, Y.-P. The Role of Extracellular Heat Shock Proteins in Cardiovascular Diseases. Biomedicines 2023, 11, 1557. [Google Scholar] [CrossRef]
- Chelladurai, P.; Dabral, S.; Basineni, S.R.; Chen, C.-N.; Schmoranzer, M.; Bender, N.; Feld, C.; Nötzold, R.R.; Dobreva, G.; Wilhelm, J.; et al. Isoform-specific characterization of class I histone deacetylases and their therapeutic modulation in pulmonary hypertension. Sci. Rep. 2020, 10, 12864. [Google Scholar] [CrossRef]
- Ma, J.; Guo, X.; Zhang, S.; Liu, H.; Lu, J.; Dong, Z.; Liu, K.; Ming, L. Trichostatin A, a histone deacetylase inhibitor, suppresses proliferation and promotes apoptosis of esophageal squamous cell lines. Mol. Med. Rep. 2015, 11, 4525–4531. [Google Scholar] [CrossRef]
- Vigushin, D.M.; Ali, S.; Pace, P.E.; Mirsaidi, N.; Ito, K.; Adcock, I.; Coombes, R.C. Trichostatin A Is a Histone Deacetylase Inhibitor with Potent Antitumor Activity against Breast Cancer in Vivo1. Clin. Cancer Res. 2001, 7, 971–976. [Google Scholar] [PubMed]
- Muslin, A.J. MAPK signalling in cardiovascular health and disease: Molecular mechanisms and therapeutic targets. Clin. Sci. 2008, 115, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Elimban, V.; Nijjar, M.S.; Gupta, S.K.; Dhalla, N.S. Role of mitogen-activated protein kinase in cardiac hypertrophy and heart failure. Exp. Clin. Cardiol. 2003, 8, 173–183. [Google Scholar] [PubMed]
- Yang, D.; Xie, P.; Liu, Z. Ischemia/reperfusion-induced MKP-3 impairs endothelial NO formation via inactivation of ERK1/2 pathway. PLoS ONE 2012, 7, e42076. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.; Garcia, T.; Aniqa, M.; Ali, S.; Ally, A.; Nauli, S.M. Endothelial Nitric Oxide Synthase (eNOS) and the Cardiovascular System: In Physiology and in Disease States. Am. J. Biomed. Sci. Res. 2022, 15, 153–177. [Google Scholar] [PubMed]
- Turner, N.A.; Blythe, N.M. Cardiac Fibroblast p38 MAPK: A Critical Regulator of Myocardial Remodeling. J. Cardiovasc. Dev. Dis. 2019, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Somanna, N.K.; Valente, A.J.; Krenz, M.; McDonald, K.S.; Higashi, Y.; Noda, M.; Chandrasekar, B. Histone deacetyltransferase inhibitors Trichostatin A and Mocetinostat differentially regulate MMP9, IL-18 and RECK expression, and attenuate Angiotensin II-induced cardiac fibroblast migration and proliferation. Hypertens. Res. 2016, 39, 709–716. [Google Scholar] [CrossRef]
- Houtkooper, R.H.; Pirinen, E.; Auwerx, J. Sirtuins as regulators of metabolism and healthspan. Nat. Rev. Mol. Cell Biol. 2012, 13, 225–238. [Google Scholar] [CrossRef]
- Vassilopoulos, A.; Fritz, K.S.; Petersen, D.R.; Gius, D. The human sirtuin family: Evolutionary divergences and functions. Hum. Genom. 2011, 5, 485. [Google Scholar] [CrossRef]
- Bindu, S.; Pillai, V.B.; Gupta, M.P. Role of Sirtuins in Regulating Pathophysiology of the Heart. Trends Endocrinol. Metab. 2016, 27, 563–573. [Google Scholar] [CrossRef]
- Carafa, V.; Rotili, D.; Forgione, M.; Cuomo, F.; Serretiello, E.; Hailu, G.S.; Jarho, E.; Lahtela-Kakkonen, M.; Mai, A.; Altucci, L. Sirtuin functions and modulation: From chemistry to the clinic. Clin. Epigenet. 2016, 8, 61. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Michno, A.; Grużewska, K.; Ronowska, A.; Gul-Hinc, S.; Zyśk, M.; Jankowska-Kulawy, A. Resveratrol Inhibits Metabolism and Affects Blood Platelet Function in Type 2 Diabetes. Nutrients 2022, 14, 1633. [Google Scholar] [CrossRef]
- Zheng, X.; Hai, J.; Yang, Y.; Zhang, C.; Ma, X.; Kong, B.; Zhao, Y.; Hu, Y.; Bu, P.; Ti, Y. Effects of resveratrol supplementation on cardiac remodeling in hypertensive patients: A randomized controlled clinical trial. Hypertens. Res. 2023, 46, 1493–1503. [Google Scholar] [CrossRef] [PubMed]
- Kuro-o, M. The Klotho proteins in health and disease. Nat. Rev. Nephrol. 2019, 15, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xiang, H.; Liu, J.; Chen, Y.; He, R.R.; Liu, B. Mitochondrial Sirtuin 3: New emerging biological function and therapeutic target. Theranostics 2020, 10, 8315–8342. [Google Scholar] [CrossRef] [PubMed]
- Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Health Sci. 2018, 12, 88–93. [Google Scholar]
- Mathieu, L.; Lopes Costa, A.; Le Bachelier, C.; Slama, A.; Lebre, A.-S.; Taylor, R.W.; Bastin, J.; Djouadi, F. Resveratrol attenuates oxidative stress in mitochondrial Complex I deficiency: Involvement of SIRT3. Free Radic. Biol. Med. 2016, 96, 190–198. [Google Scholar] [CrossRef]
- Dyck, G.J.B.; Raj, P.; Zieroth, S.; Dyck, J.R.B.; Ezekowitz, J.A. The Effects of Resveratrol in Patients with Cardiovascular Disease and Heart Failure: A Narrative Review. Int. J. Mol. Sci. 2019, 20, 904. [Google Scholar] [CrossRef]
- Mao, Q.; Liang, X.; Wu, Y.; Lu, Y. Resveratrol Attenuates Cardiomyocyte Apoptosis in Rats Induced by Coronary Microembolization Through SIRT1-Mediated Deacetylation of p53. J. Cardiovasc. Pharmacol. Ther. 2019, 24, 551–558. [Google Scholar] [CrossRef]
- Ma, S.; Feng, J.; Zhang, R.; Chen, J.; Han, D.; Li, X.; Yang, B.; Li, X.; Fan, M.; Li, C.; et al. SIRT1 Activation by Resveratrol Alleviates Cardiac Dysfunction via Mitochondrial Regulation in Diabetic Cardiomyopathy Mice. Oxid. Med. Cell. Longev. 2017, 2017, 4602715. [Google Scholar] [CrossRef] [PubMed]
- Marmorstein, R. Structure and function of histone acetyltransferases. Cell. Mol. Life Sci. 2001, 58, 693–703. [Google Scholar] [CrossRef]
- Lee, K.K.; Workman, J.L. Histone acetyltransferase complexes: One size doesn’t fit all. Nat. Rev. Mol. Cell Biol. 2007, 8, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Costantino, S.; Libby, P.; Kishore, R.; Tardif, J.-C.; El-Osta, A.; Paneni, F. Epigenetics and precision medicine in cardiovascular patients: From basic concepts to the clinical arena. Eur. Heart J. 2018, 39, 4150–4158. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Miao, X.; Liu, Y.; Li, F.; Liu, Q.; Sun, J.; Cai, L. Dysregulation of histone acetyltransferases and deacetylases in cardiovascular diseases. Oxid. Med. Cell. Longev. 2014, 2014, 641979. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, Y.; Ren, J. Acetylation in cardiovascular diseases: Molecular mechanisms and clinical implications. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2020, 1866, 165836. [Google Scholar] [CrossRef]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The Essential Medicinal Chemistry of Curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef]
- Balasubramanyam, K.; Varier, R.A.; Altaf, M.; Swaminathan, V.; Siddappa, N.B.; Ranga, U.; Kundu, T.K. Curcumin, a Novel p300/CREB-binding Protein-specific Inhibitor of Acetyltransferase, Represses the Acetylation of Histone/Nonhistone Proteins and Histone Acetyltransferase-dependent Chromatin Transcription*. J. Biol. Chem. 2004, 279, 51163–51171. [Google Scholar] [CrossRef]
- Marcu, M.G.; Jung, Y.J.; Lee, S.; Chung, E.J.; Lee, M.J.; Trepel, J.; Neckers, L. Curcumin is an inhibitor of p300 histone acetylatransferase. Med. Chem. 2006, 2, 169–174. [Google Scholar] [CrossRef]
- Hassan, F.-u.; Rehman, M.S.-u.; Khan, M.S.; Ali, M.A.; Javed, A.; Nawaz, A.; Yang, C. Curcumin as an Alternative Epigenetic Modulator: Mechanism of Action and Potential Effects. Front. Genet. 2019, 10, 514. [Google Scholar] [CrossRef]
- Morimoto, T.; Sunagawa, Y.; Kawamura, T.; Takaya, T.; Wada, H.; Nagasawa, A.; Komeda, M.; Fujita, M.; Shimatsu, A.; Kita, T.; et al. The dietary compound curcumin inhibits p300 histone acetyltransferase activity and prevents heart failure in rats. J. Clin. Investig. 2008, 118, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Sunagawa, Y.; Funamoto, M.; Shimizu, K.; Shimizu, S.; Sari, N.; Katanasaka, Y.; Miyazaki, Y.; Kakeya, H.; Hasegawa, K.; Morimoto, T. Curcumin, an Inhibitor of p300-HAT Activity, Suppresses the Development of Hypertension-Induced Left Ventricular Hypertrophy with Preserved Ejection Fraction in Dahl Rats. Nutrients 2021, 13, 2608. [Google Scholar] [CrossRef] [PubMed]
- Välimäki, M.J.; Ruskoaho, H.J. Targeting GATA4 for cardiac repair. IUBMB Life 2020, 72, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Chen, Y.; Chen, Q.; Hong, T.; Zhong, Z.; He, J.; Ni, C. Curcumin Ameliorates Cardiac Fibrosis by Regulating Macrophage-Fibroblast Crosstalk via IL18-P-SMAD2/3 Signaling Pathway Inhibition. Front. Pharmacol. 2022, 12, 784041. [Google Scholar] [CrossRef]
- Li, C.; Meng, X.; Wang, L.; Dai, X. Mechanism of action of non-coding RNAs and traditional Chinese medicine in myocardial fibrosis: Focus on the TGF-β/Smad signaling pathway. Front. Pharmacol. 2023, 14, 1092148. [Google Scholar] [CrossRef]
- Qin, S.; Huang, L.; Gong, J.; Shen, S.; Huang, J.; Ren, H.; Hu, H. Efficacy and safety of turmeric and curcumin in lowering blood lipid levels in patients with cardiovascular risk factors: A meta-analysis of randomized controlled trials. Nutr. J. 2017, 16, 68. [Google Scholar] [CrossRef]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; García-Almagro, F.J.; Avilés-Plaza, F.; Parra, S.; Yáñez-Gascón, M.J.; Ruiz-Ros, J.A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; et al. Consumption of a grape extract supplement containing resveratrol decreases oxidized LDL and ApoB in patients undergoing primary prevention of cardiovascular disease: A triple-blind, 6-month follow-up, placebo-controlled, randomized trial. Mol. Nutr. Food Res. 2012, 56, 810–821. [Google Scholar] [CrossRef]
- Tomé-Carneiro, J.; Larrosa, M.; Yáñez-Gascón, M.J.; Dávalos, A.; Gil-Zamorano, J.; Gonzálvez, M.; García-Almagro, F.J.; Ruiz Ros, J.A.; Tomás-Barberán, F.A.; Espín, J.C.; et al. One-year supplementation with a grape extract containing resveratrol modulates inflammatory-related microRNAs and cytokines expression in peripheral blood mononuclear cells of type 2 diabetes and hypertensive patients with coronary artery disease. Pharmacol. Res. 2013, 72, 69–82. [Google Scholar] [CrossRef]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; Yáñez-Gascón, M.J.; García-Almagro, F.J.; Ruiz-Ros, J.A.; Tomás-Barberán, F.A.; García-Conesa, M.T.; Espín, J.C. Grape Resveratrol Increases Serum Adiponectin and Downregulates Inflammatory Genes in Peripheral Blood Mononuclear Cells: A Triple-Blind, Placebo-Controlled, One-Year Clinical Trial in Patients with Stable Coronary Artery Disease. Cardiovasc. Drugs Ther. 2013, 27, 37–48. [Google Scholar] [CrossRef]
- Garg, A.X.; Devereaux, P.J.; Hill, A.; Sood, M.; Aggarwal, B.; Dubois, L.; Hiremath, S.; Guzman, R.; Iyer, V.; James, M.; et al. Oral curcumin in elective abdominal aortic aneurysm repair: A multicentre randomized controlled trial. Can. Med. Assoc. J. 2018, 190, E1273–E1280. [Google Scholar] [CrossRef]
- Wongcharoen, W.; Jai-Aue, S.; Phrommintikul, A.; Nawarawong, W.; Woragidpoonpol, S.; Tepsuwan, T.; Sukonthasarn, A.; Apaijai, N.; Chattipakorn, N. Effects of curcuminoids on frequency of acute myocardial infarction after coronary artery bypass grafting. Am. J. Cardiol. 2012, 110, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhang, H.; Huang, S.; Yin, L.; Wang, F.; Luo, P.; Huang, H. Epigenetic regulation in cardiovascular disease: Mechanisms and advances in clinical trials. Signal Transduct. Target. Ther. 2022, 7, 200. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-L.; Lin, Z.-J.; Li, C.-C.; Lin, X.; Shan, S.-K.; Guo, B.; Zheng, M.-H.; Li, F.; Yuan, L.-Q.; Li, Z.-h. Epigenetic regulation in metabolic diseases: Mechanisms and advances in clinical study. Signal Transduct. Target. Ther. 2023, 8, 98. [Google Scholar] [CrossRef] [PubMed]
- Sarno, F.; Benincasa, G.; List, M.; Barabasi, A.-L.; Baumbach, J.; Ciardiello, F.; Filetti, S.; Glass, K.; Loscalzo, J.; Marchese, C.; et al. Clinical epigenetics settings for cancer and cardiovascular diseases: Real-life applications of network medicine at the bedside. Clin. Epigenet. 2021, 13, 66. [Google Scholar] [CrossRef]

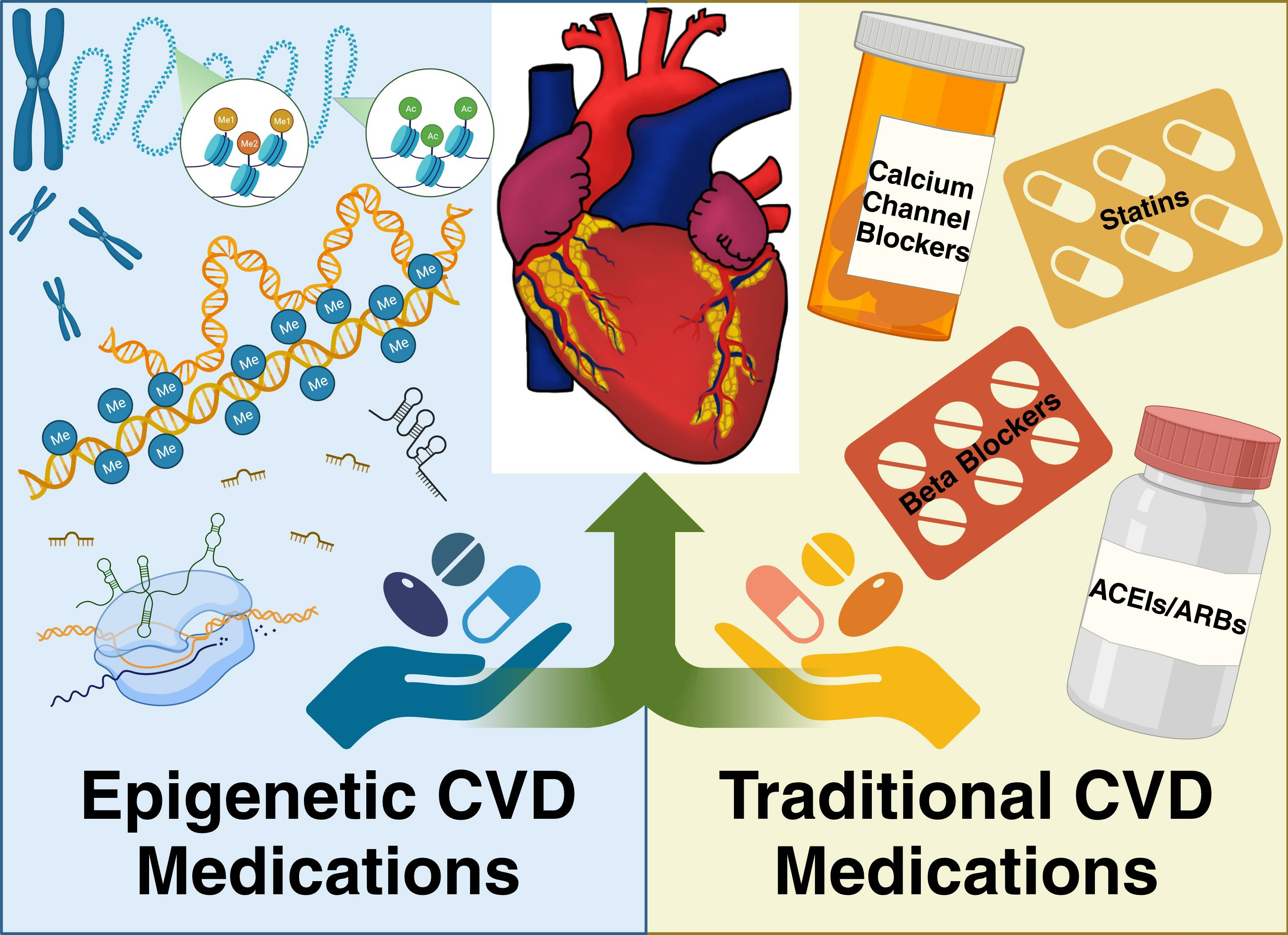{kind=link}
| Epigenetic Drugs | Mechanism of Action | Similar Traditional Drug |
|---|---|---|
| Valproic Acid | Decreases the transcription of the mineralocorticoid receptor to reduce sodium and water retention, inhibits cardiac remodeling, and attenuates atrial fibrillation | Statins, ARBs, ACEIs, Beta Blockers |
| Sodium Phenylbutyrate | Increases MnSOD activity, mitigates reperfusion therapy post-MI, and reduces ER stress | Calcium Channel Blockers |
| Vorinostat | Anti-inflammatory, prevents cardiac remodeling, attenuates cardiac dysfunction, lowers systolic and pulmonary arterial pressure, inhibits cardiac hypertrophy | ACEIs, Statins, ARBs |
| Trichostatin A | Decreases cardiac hypertrophy, inhibits MAPK pathway activation, decreases inflammatory marker expression, and improves eNOS expression | Statins, ARBs |
| Resveratrol | Anti-inflammatory, inhibits cardiomyocyte apoptosis, inhibits platelet adhesion and aggregation, increases SOD expression, inhibits oxidative stress, and improves mitochondrial function | Statins, Calcium Channel Blockers, Beta Blockers |
| Curcumin (Turmeric) | Inhibits miRNA expression, mitigates cardiac hypertrophy, inhibits IL-6, IL-1β, IL-18, and TNF-α, reduces serum lipid levels, and reduces CVD risk factors | Statins, ACEIs, ARBs, |
| Drug | Condition/Disease | Status | NCT Number | Phase |
|---|---|---|---|---|
| HDACi | ||||
| Valproic Acid | Thrombosis | Completed | NCT03903302 | Phase 1 |
| Subarachnoid Hemorrhage | Terminated | NCT00774306 | N/A | |
| Anoxic Encephalopathy; Cardiac Arrest; Status Epilepticus; | Completed | NCT02056236 | N/A | |
| Brain Injuries, Acute; Brain Injuries, Traumatic; Brain Ischemia; Brain Hypoxia; Hypoxia-Ischemia, Brain; Heart Arrest; Stroke; Intracranial Hemorrhages; Coma; Persistent Vegetative State; | Not Yet Recruiting | NCT06081283 | Phase 4 | |
| Acute Ischemic Stroke | Recruiting | NCT06020898 | Phase 2 | |
| Cardiac Valve disease; Coronary Brain Artery Disease; Organ Failure, Multiple; | Unknown Status | NCT03825250 | Phase 1, Phase 2 | |
| Cerebral Aneurysm | Unknown Status | NCT01460563 | N/A | |
| Acute Kidney Injury; Ischemia Reperfusion Injury | Withdrawn | NCT04531579 | Phase 2 | |
| Acute Kidney Injury; Ischemia Reperfusion Injury | Withdrawn | NCT04531592 | Phase 2 | |
| Post Cerebral Hemorrhage | Completed | NCT01115959 | Phase 4 | |
| Cardiac Arrest | Unknown Status | NCT01083784 | Phase 4 | |
| Cardiac Arrest; Status Epilepticus | Recruiting | NCT05756621 | ||
| Pulmonary Arterial Hypertension | Recruiting | NCT05224531 | Phase 2 | |
| Sodium Phenylbutyrate | No Recorded Studies | |||
| Vorinostat | No Recorded Studies | |||
| Trichostatin A | No Recorded Studies | |||
| STAC | ||||
| Resveratrol | Cardiovascular Diseases | Completed | NCT01449110 | Phase 2 |
| Cardiovascular Disease | Completed | NCT01564381 | Phase 1, Phase 2 | |
| Hypercholesterolemia | Completed | NCT02409537 | Phase 2 | |
| Vascular System Injuries, Lipid Metabolism Disorders, Endothelial Dysfunction; | Completed | NCT01668836 | N/A | |
| Peripheral Artery Disease | Completed | NCT02246660 | N/A | |
| Metabolic Syndrome, Coronary Artery Disease | Completed | NCT02137421 | N/A | |
| Cardiovascular Diseases; Autonomic Nervous System; Polyphenols; | Recruiting | NCT06020313 | N/A | |
| Coronary Artery Disease; Diabetes Mellitus, Type 2; | Active, Not Recruiting | NCT03762096 | Phase 1, Phase 2 | |
| Pulmonary Disease, Chronic Obstructive; | Completed | NCT02245932 | N/A | |
| Chronic Obstructive Pulmonary Disease | Completed | NCT03819517 | N/A | |
| Peripheral Artery Disease | Completed | NCT03743636 | Phase 3 | |
| Hypertension; Vascular Resistance; | Terminated | NCT01842399 | Phase 1, Phase 2 | |
| Coronary Artery Disease; Endothelial Dysfunction; Menopause | Recruiting | NCT05808387 | N/A | |
| Cardiovascular Disease; Atherosclerosis; Inflammation; | Unknown Status | NCT02998918 | Phase 2 | |
| Autonomic Nervous System Disease; Blood Pressure; Coronary Artery Disease; Heart Rate; | Completed | NCT06095635 | N/A | |
| Congestive Heart Failure Chronic | Completed | NCT03525379 | Phase 2 | |
| Dilated Cardiomyopathy | Unknown Status | NCT01914081 | Phase 3 | |
| Coronary Artery Disease; Metabolic Syndrome | Completed | NCT02137421 | N/A | |
| Coronary Artery Restenosis; In-stent Coronary Artery Restenosis; In-stent Restenosis; | Withdrawn | NCT05093244 | N/A | |
| Diastolic Heart Failure; Heart Failure with Preserved Ejection Fraction; Hypertension; Hypertensive Heart Disease; Oxidative Stress; | Completed | NCT01185067 | Phase 1 | |
| Hypertension in Pregnancy; Intrauterine Growth Restriction; Pre-Eclampsia; Pre-Term; | Completed | NCT04633551 | N/A | |
| HATi | ||||
| Curcumin | Cardiovascular Disease | Completed | NCT02088307 | N/A |
| Abdominal Aortic Aneurysm; Acute Kidney Injury; | Completed | NCT01225094 | Phase 2, Phase 3 | |
| Coronary Artery Disease; Inflammation; Oxidative Stress; | Unknown Status | NCT04458116 | N/A | |
| Cardiovascular Disease; Atherosclerosis; Inflammation; | Unknown Status | NCT02998918 | Phase 2 | |
| Cardiovascular Abnormalities; Type 2 Diabetes Mellitus; | Unknown Status | NCT01052597 | Phase 4 | |
| Diabetes Mellitus, Type 2; Dyslipidemias; Hypertension | Recruiting | NCT05753436 | Phase 2 | |
| Cardiovascular Risk; Insulin Resistance; Pre-diabetes; Type 2 Diabetes; | Unknown Status | NCT01052025 | Phase 4 | |
| Heart Diseases; High Blood Pressure; High Cholesterol; Obesity; Type2 Diabetes; | Completed | NCT03542240 | N/A | |
| Blood Pressure; Chronic Kidney Diseases; Hyperemia; Vasoconstriction; | Withdrawn | NCT04132648 | Phase 2 | |
| Hematoma, Subdural, Chronic; | Withdrawn | NCT03845322 | Early Phase 1 | |
| Hypertension in Pregnancy; Intrauterine Growth Restriction; Pre-Eclampsia; Pre-Term; | Completed | NCT04633551 | N/A | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gladwell, L.R.; Ahiarah, C.; Rasheed, S.; Rahman, S.M.; Choudhury, M. Traditional Therapeutics and Potential Epidrugs for CVD: Why Not Both? Life 2024, 14, 23. https://doi.org/10.3390/life14010023
Gladwell LR, Ahiarah C, Rasheed S, Rahman SM, Choudhury M. Traditional Therapeutics and Potential Epidrugs for CVD: Why Not Both? Life. 2024; 14(1):23. https://doi.org/10.3390/life14010023
Chicago/Turabian StyleGladwell, Lauren Rae, Chidinma Ahiarah, Shireen Rasheed, Shaikh Mizanoor Rahman, and Mahua Choudhury. 2024. "Traditional Therapeutics and Potential Epidrugs for CVD: Why Not Both?" Life 14, no. 1: 23. https://doi.org/10.3390/life14010023
APA StyleGladwell, L. R., Ahiarah, C., Rasheed, S., Rahman, S. M., & Choudhury, M. (2024). Traditional Therapeutics and Potential Epidrugs for CVD: Why Not Both? Life, 14(1), 23. https://doi.org/10.3390/life14010023