
Pressure as a Limiting Factor for Life

{kind=link}
{kind=link}
Abstract
:1. Introduction
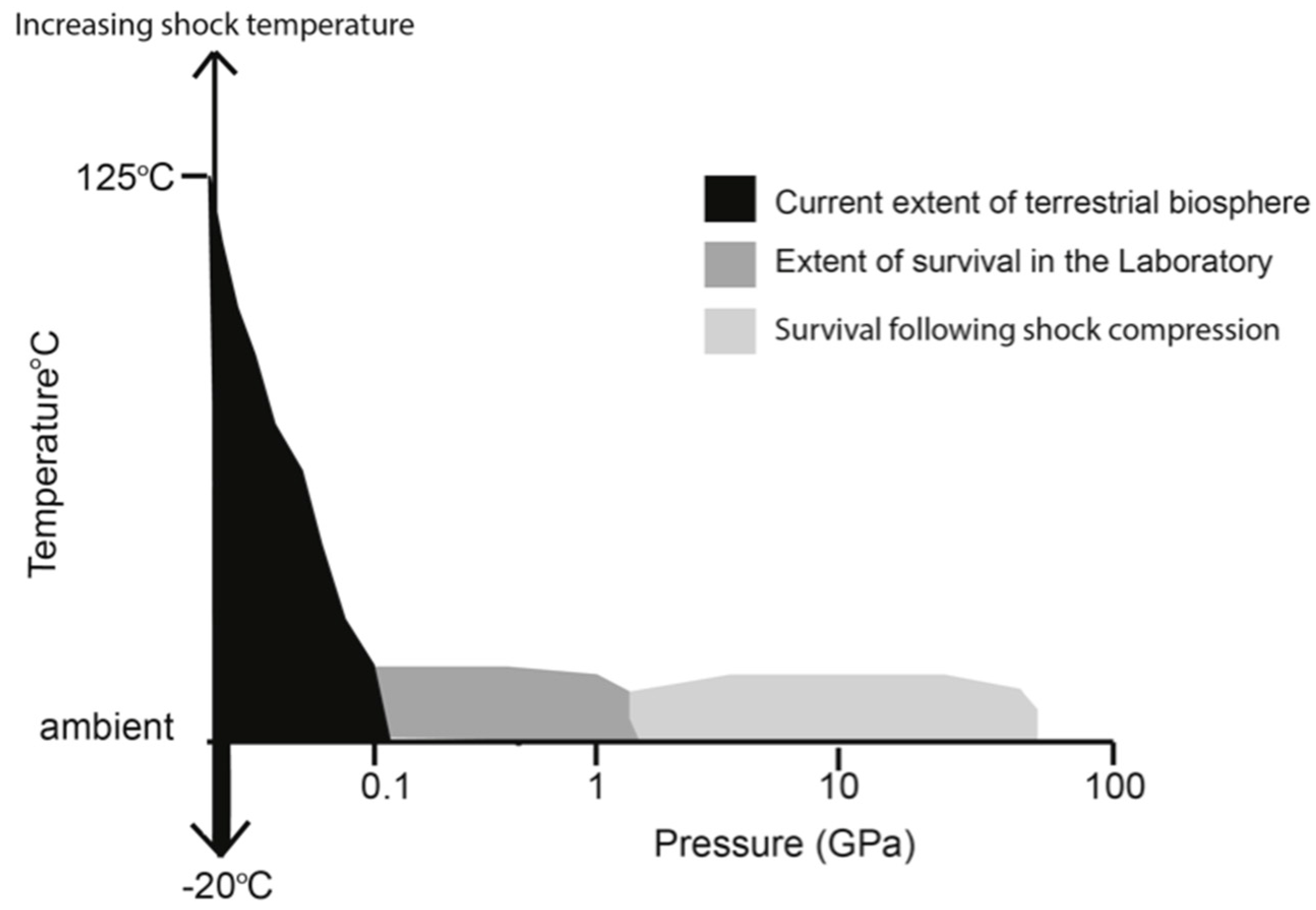
2. Pressure Stability Limits for Biological Macromolecules and Their Complexes
3. Actual Results for Bacterial Survival into the GPa Range
4. Water in Cells
5. Timescales of Biological Processes
6. Complex Lifeforms
7. Compression on a Rapid Timescale
8. Conclusions
Supplementary Materials
Acknowledgments
Author contributions
Conflicts of Interest
References
- Melosh, H.J. The rocky road to panspermia. Nature 1988, 332, 687–688. [Google Scholar] [CrossRef] [PubMed]
- Yayanos, A.A. Microbiology to 10,500 Meters in the Deep-Sea. Ann. Rev. Microbiol. 1995, 49, 777–805. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.J.; Becker, K.; Colwell, F. The Deep, Dark Energy Biosphere: Intraterrestrial Life on Earth. Ann. Rev. Earth Planet. Sci. 2012, 40, 551–568. [Google Scholar] [CrossRef]
- Gold, T. The deep, hot biosphere. Proc. Nat. Acad. Sci. USA 1992, 89, 6045–6049. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, L.J.; Mancinelli, R.L. Life in extreme environments. Nature 2001, 409, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Whitman, W.B.; Coleman, D.C.; Wiebe, W.J. Prokaryotes: The unseen majority. Proc. Nat. Acad. Sci. USA 1998, 95, 6578–6583. [Google Scholar] [CrossRef] [PubMed]
- Huber, J.A. Making methane down deep. Science 2015, 349, 376–377. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, F.; Hinrichs, K.-U.; Kubo, Y.; Bowles, M.W.; Heuer, V.B.; Hong, W.-L.; Hoshino, T.; Ijiri, A.; Imachi, H.; Ito, M.; et al. Exploring deep microbial life in coal-bearing sediment down to ~2.5 km below the ocean floor. Science 2015, 349, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Paulino-Lima, I.G.; Pilling, S.; Janot-Pacheco, E.; de Brito, A.N.; Barbosa, J.A.R.G.; Leitão, A.C.; Lage, C.D.A.S. Laboratory simulation of interplanetary ultraviolet radiation (broad spectrum) and its effects on Deinococcus radiodurans. Planet. Space Sci. 2010, 58, 1180–1187. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Dopson, M. Life in acid: pH homeostasis in acidophiles. Trend. Microbiol. 2007, 15, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Padan, E.; Bibi, E.; Ito, M.; Krulwich, T.A. Alkaline pH Homeostasis in Bacteria: New Insights. BBA Biomembr. 2005, 1717, 67–88. [Google Scholar] [CrossRef] [PubMed]
- Borgonie, G.; Garcia-Moyano, A.; Litthauer, D.; Bert, W.; Bester, A.; van Heerden, E.; Moller, C.; Erasmus, M.; Onstott, T.C. Nematoda from the terrestrial deep subsurface of South Africa. Nature 2011, 474, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Schiffries, C.M.; Hazen, R.M. Why Deep Carbon? Rev. Miner. Geochem. 2013, 74, 1–6. [Google Scholar]
- Kargel, J.S.; Kaye, J.Z.; Head, J.W.; Marion, G.M.; Sassen, R.; Crowley, J.K.; Ballesteros, O.P.; Grant, S.A.; Hogenboom, D.L. Europa’s Crust and Ocean: Origin, Composition, and the Prospects for Life. Icarus 2000, 148, 226–265. [Google Scholar] [CrossRef]
- Lorenz, R.D. Thermodynamics of Geysers: Application to Titan. Icarus 2002, 156, 176–183. [Google Scholar] [CrossRef]
- Gaidos, E.J.; Nealson, K.H.; Kirschvink, J.L. Life in Ice-Covered Oceans. Science 1999, 284, 1631–1633. [Google Scholar] [CrossRef] [PubMed]
- Bridgman, P.W. The coagulation of albumen by pressure. J. Biol. Chem. 1914, 19, 511–512. [Google Scholar]
- Meersman, F.; Daniel, I.; Bartlett, D.H.; Winter, R.; Hazael, R.; McMillan, P.F. High pressure biochemistry and biophysics. Rev. Miner. Geochem. 2013, 75, 607–648. [Google Scholar] [CrossRef]
- Winter, R.; Jeworrek, C. Effect of pressure on membranes. Soft Matter 2009, 5, 3157–3173. [Google Scholar] [CrossRef]
- Bernsdorff, C.; Wolf, A.; Winter, R. The Effect of Temperature and Pressure on Structural and Dynamic Properties of Phospholipid/Sterol Mixtures- A Steady-State and Time-Resolved Fluorescence Anisotropy Study. Z. Phys. Chem. 1996, 193, 151–173. [Google Scholar] [CrossRef]
- Hauben, K.J.; Bartlett, D.H.; Soontjens, C.C.; Cornelis, K.; Wuytack, E.Y.; Michiels, C.W. Escherichia coli mutants resistant to inactivation by high hydrostatic pressure. Appl. Environ. Microbiol. 1997, 63, 945–950. [Google Scholar] [PubMed]
- Sharma, A.; Scott, J.H.; Cody, G.D.; Fogel, M.L.; Hazen, R.M.; Hemley, R.J.; Huntress, W.T. Microbial activity at gigapascal pressures. Science 2002, 295, 1514–1516. [Google Scholar] [CrossRef] [PubMed]
- Vanlint, D.; Mitchell, R.; Bailey, E.; Meersman, F.; McMillan, P.F.; Michiels, C.W.; Aertsen, A. Rapid acquisition of gigapascal-high-pressure resistance by Escherichia coli. Mbiology 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Hazael, R.; Foglia, F.; Kardzhaliyska, L.; Daniel, I.; Meersman, F.; McMillan, P.F. Laboratory Investigation of High Pressure Survival in Shewanella oneidensis MR-1 into the GigaPascal Pressure Range. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Ball, P. Water as an Active Constituent in Cell Biology. Chem. Rev. 2008, 108, 74–108. [Google Scholar] [CrossRef] [PubMed]
- Bove, L.E.; Klotz, S.; Strassle, Th.; Koza, M.; Teixeira, J.; Saitta, A.M. Translational and Rotational Diffusion in Water in the Gigapascal Range. Phys. Rev. Lett. 2013, 111, 185901. [Google Scholar] [CrossRef] [PubMed]
- Tehei, M.; Jasnin, M.; Stadler, A.; Zaccai, G. Specific cellular water dynamics observed in vivo by neutron scattering and NMR. Phys. Chem. Chem. Phys. 2010, 12, 10154–10160. [Google Scholar]
- Le Bihan, D. Looking into the functional architecture of the brain with diffusion MRI. Nat. Rev. Neurosci. 2003, 4, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Le Bihan, D. The “wet mind”: Water and functional neuroimaging. Phys. Med. Biol. 2007, 52, R57. [Google Scholar] [CrossRef] [PubMed]
- Picard, A.; Daniel, I.; Testemale, D.; Kieffer, I.; Bleuet, P.; Cardon, H.; Oger, P.M. Monitoring microbial redox transformations of metal and metalloid elements inder high pressure using in situ X-ray absorption spectroscopy. Geobiology 2011, 9, 196–204. [Google Scholar] [PubMed]
- Foglia, F.; Hazael, R.; Simeoni, G.G.; Appavou, M.-S.; Moulin, M.; Haertlein, M.; Forsyth, V.T.; Seydel, T.; Daniel, I.; Meersman, F.; et al. Water Dynamics in Shewanella oneidensis at Ambient and High Pressure using Quasi-Elastic Neutron Scattering. Sci. Rep. 2016, 6, 18862. [Google Scholar] [CrossRef] [PubMed]
- Colwell, F.S.; D’Hondt, S. Nature and Extent of the Deep Biosphere. Rev. Miner. Geochem. 2013, 75, 547–574. [Google Scholar] [CrossRef]
- Zeng, X.; Birren, J.-L.; Fouquet, Y.; Cherkashov, G.; Jebbar, M.; Querellou, J.; Oger, P.; Cambon-Bonavita, M.-A.; Xiao, X.; Prieur, D. Pyrococcus CH1, an obligate piezophilic hyperthermophile: Extending the upper pressure-temperature limits for life. Isme J. 2009, 3, 873–876. [Google Scholar] [CrossRef] [PubMed]
- Clegg, J.S. Cryptobiosis—A peculiar state of biological organization. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2001, 128, 613–624. [Google Scholar] [CrossRef]
- Keilin, D. The Leeuwenhoek Lecture: The Problem of Anabiosis or Latent Life: History and Current Concept. Proc. R. Soc. Lond. B Biol. Sci. 1959, 150, 149–191. [Google Scholar] [CrossRef] [PubMed]
- Fox-Skelly, J. The Strange Beasts that Live in Solid Rock Deep Underground. 2015. Available online: http://www.bbc.co.uk/earth/story/20151124-meet-the-strange-creatures-that-live-in-solid-rock-deep-underground (accessed on 20 May 2016).
- O’Callaghan, J. World’s Deepest Fish Found: Ghostly Snailfish Is Found Lurking 27,000 ft below at the Bottom of the Pacific’s Mariana Trench. 2014. Available online: http://www.dailymail.co.uk/sciencetech/article-2880612/World-s-deepest-fish-Ghostly-snailfish-27-000ft-deep-bottom-Pacific-s-Mariana-Trench.html (accessed on 20 May 2016).
- Lee, J.J. Weird Animal World Discovered in Deepest Pacific Ocean Vents. 2015. Available online: http://news.nationalgeographic.com/2015/06/150610-hydrothermal-vent-gulf-california-ocean-earth-science/ (accessed on 20 May 2016).
- Frazer, J. What Lives at the Bottom of the Mariana Trench? More Than You Might Think. 2013. Available online: http://blogs.scientificamerican.com/artful-amoeba/what-lives-at-the-bottom-of-the-mariana-trench-more-than-you-might-think/ (accessed on 20 May 2016).
- Nishihira, N.; Shindo, A.; Saigusa, M.; Ono, F.; Matsushima, Y.; Mori, Y.; Takarabe, K.; Saini, N.L.; Yamashita, M. Preserving life of moss Ptychomitrium under very high pressure. J. Phys. Chem. Solids 2010, 71, 1123–1126. [Google Scholar] [CrossRef]
- Ono, F.; Saigusa, M.; Uozumi, T.; Matsushima, Y.; Ikeda, H.; Saini, N.L.; Yamashita, M. Effect of high hydrostatic pressure on to life of the tiny animal tardigrade. J. Phys. Chem. Solid. 2008, 69, 2297–2300. [Google Scholar] [CrossRef]
- Ono, F.; Mori, Y.; Sougawa, M.; Takarabe, K.; Hada, Y.; Nishihira, N.; Motose, H.; Saigusa, M.; Matsushima, Y.; Yamazaki, D.; et al. Effects of very high pressure on life of plants and animals. J. Phys. Conf. Ser. 2011, 377, 012053. [Google Scholar] [CrossRef]
- Gupta, R.K.; Das, S.K. Fracture resistance of sunflower seed and kernal to compressive loading. J. Food Eng. 2000, 46, 1–8. [Google Scholar] [CrossRef]
- Ono, F.; Minami, K.; Saigusa, M.; Matsushima, Y.; Mori, Y.; Takarabe, K.; Saini, N.L.; Yamashita, M. Life of Artemia under very high pressure. J. Phys. Chem. Solids 2010, 71, 1127–1130. [Google Scholar] [CrossRef]
- Govers, S.K.; Aertsen, A. Impact of high hydrostatic pressure processing on individual cellular resuscitation times and protein aggregates in Escherichia coli. Int. J. Food Microbiol. 2015, 213, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Hazael, R.; Fitzmaurice, B.C.; Foglia, F.; Appleby-Thomas, G.J.; McMillan, P.F. Bacterial survival following shock compression in the GigaPascal range. Icarus 2016. submitted. [Google Scholar]
- Burchell, M.J.; Mann, J.R.; Bunch, A.W. Survival of bacteria and spores under extreme shock pressures. Mon. Not. R. Astron. Soc. 2004, 1278, 1273–1278. [Google Scholar] [CrossRef]
- Vadillo-Rodriguez, V.; Beveridge, T.J.; Dutcher, J.R. Surface Viscoelasticity of Individual Gram-Negative Bacterial Cells Measured Using Atomic Force Microscopy. J. Bacteriol. 2008, 190. [Google Scholar] [CrossRef] [PubMed]
- Mustey, A.; Leighs, J.A.; Appleby-Thomas, G.J.; Wood, D.C.; Hazael, R.; McMillan, P.F.; Hazell, P.F. The effect of hydrostatic vs. shock pressure treatment of plant seeds. J. Phys. Conf. Ser. 2014, 500, 182025. [Google Scholar] [CrossRef]
- Burchell, M.J.; Mann, J.; Bunch, A.W.; Brand, P.F.B. Survivability of Bacteria in Hypervelocity Impact. Icarus 2001, 547, 545–547. [Google Scholar] [CrossRef]
- Price, M.C.; Solscheid, C.; Burchell, M.J.; Josse, L.; Adamek, N.; Cole, M.J. Survival of yeast spores in hypervelocity impact events up to velocities of 7.4 km·s−1. Icarus 2013, 222, 263–272. [Google Scholar] [CrossRef]
- Beck, P.; Gillet, P.; El Goresy, A.; Mostefaoui, S. Timescales of shock processes in chondritic and martian meteorites. Nature 2005, 435, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Horneck, G.; Stöffler, D.; Ott, S.; Hornemann, U.; Cockell, C.S.; Moeller, R.; Meyer, C.; de Vera, J.P.; Fritz, J.; Schade, S.; et al. Microbial Rock Inhabitants Survive Hypervelocity Impacts on Mars-Like Host Planets: First Phase of Lithopanspermia Experimentally Tested. Astrobiology 2008, 8, 17–44. [Google Scholar] [CrossRef] [PubMed]
- Fajardo-Cavazos, P.; Langenhorst, F.; Melosh, H.J.; Nicholson, W.L. Bacterial Spores in Granite Survive Hypervelocity Launch by Spallation: Implications for Lithopanspermia. Astrobiology 2009, 9, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Martins, Z.; Price, M.C.; Goldman, N.; Sephton, M.A.; Burchell, M.J. Shock synthesis of amino acids from impacting cometary and icy planet surface analogues. Nat. Geosci. 2013, 6, 1045–1049. [Google Scholar] [CrossRef]


© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hazael, R.; Meersman, F.; Ono, F.; McMillan, P.F. Pressure as a Limiting Factor for Life. Life 2016, 6, 34. https://doi.org/10.3390/life6030034
Hazael R, Meersman F, Ono F, McMillan PF. Pressure as a Limiting Factor for Life. Life. 2016; 6(3):34. https://doi.org/10.3390/life6030034
Chicago/Turabian StyleHazael, Rachael, Filip Meersman, Fumihisa Ono, and Paul F. McMillan. 2016. "Pressure as a Limiting Factor for Life" Life 6, no. 3: 34. https://doi.org/10.3390/life6030034
APA StyleHazael, R., Meersman, F., Ono, F., & McMillan, P. F. (2016). Pressure as a Limiting Factor for Life. Life, 6(3), 34. https://doi.org/10.3390/life6030034