
Gestational Diabetes—Placental Expression of Human Equilibrative Nucleoside Transporter 1 (hENT1): Is Delayed Villous Maturation an Adaptive Pattern?

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Statistical Methods
2.1.1. Pre-Processing
2.1.2. Analysis
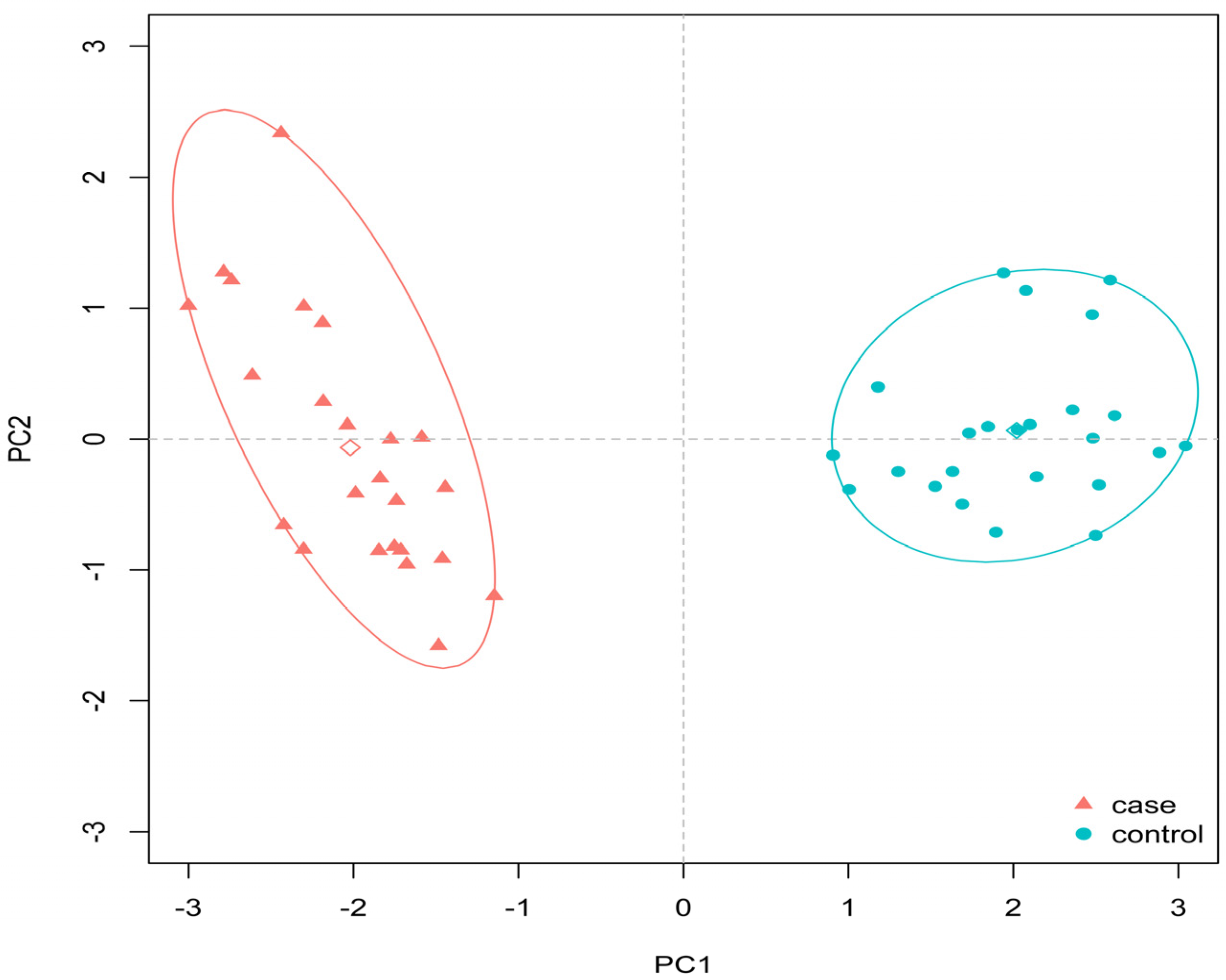
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2013, 36 (Suppl. 1), S67–S74. [Google Scholar] [CrossRef] [Green Version]
- Sivaraman, S.C.; Vinnamala, S.; Jenkins, D. Gestational diabetes and future risk of diabetes. J. Clin. Med. Res. 2013, 5, 92–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, T.A.; Xiang, A.H.; Page, K.A. Gestational diabetes mellitus: Risks and management during and after pregnancy. Nat. Rev. Endocrinol. 2012, 8, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Metzger, B.E.; Coustan, D.R.; Trimble, E.R. Hyperglycemia and Adverse Pregnancy Outcomes. Clin. Chem. 2019, 65, 937–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landon, M.B.; Mele, L.; Varner, M.W.; Casey, B.M.; Reddy, U.M.; Wapner, R.J.; Rouse, D.J.; Tita, A.T.N.; Thorp, J.M.; Chien, E.K.; et al. The relationship of maternal glycemia to childhood obesity and metabolic dysfunction. J. Matern. Fetal Neonatal Med. 2020, 33, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Cao, S.; Li, N.; Liu, J.; Lyu, Y.; Li, J.; Yang, X. Risks of overweight in the offspring of women with gestational diabetes at different developmental stages: A meta-analysis with more than half a million offspring. Obes. Rev. 2022, 23, e13395. [Google Scholar] [CrossRef]
- Leiva, A.; Pardo, F.; Ramirez, M.A.; Farias, M.; Casanello, P.; Sobrevia, L. Fetoplacental vascular endothelial dysfunction as an early phenomenon in the programming of human adult diseases in subjects born from gestational diabetes mellitus or obesity in pregnancy. Exp. Diabetes Res. 2011, 2011, 349286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman-Gutierrez, E.; Abarzua, F.; Belmar, C.; Nien, J.K.; Ramirez, M.A.; Arroyo, P.; Salomon, C.; Westermeier, F.; Puebla, C.; Leiva, A.; et al. Functional link between adenosine and insulin: A hypothesis for fetoplacental vascular endothelial dysfunction in gestational diabetes. Curr. Vasc. Pharmacol. 2011, 9, 750–762. [Google Scholar] [CrossRef]
- San Martin, R.; Sobrevia, L. Gestational diabetes and the adenosine/L-arginine/nitric oxide (ALANO) pathway in human umbilical vein endothelium. Placenta 2006, 27, 1–10. [Google Scholar] [CrossRef]
- Baldwin, S.A.; Beal, P.R.; Yao, S.Y.; King, A.E.; Cass, C.E.; Young, J.D. The equilibrative nucleoside transporter family, SLC29. Pflug. Arch. 2004, 447, 735–743. [Google Scholar] [CrossRef]
- Guzman-Gutierrez, E.; Sandoval, C.; Nova, E.; Castillo, J.L.; Vera, J.C.; Lamperti, L.; Krause, B.; Salomon, C.; Sepulveda, C.; Aguayo, C.; et al. Differential expression of functional nucleoside transporters in non-differentiated and differentiated human endothelial progenitor cells. Placenta 2010, 31, 928–936. [Google Scholar] [CrossRef]
- Sobrevia, L.; Abarzua, F.; Nien, J.K.; Salomon, C.; Westermeier, F.; Puebla, C.; Cifuentes, F.; Guzman-Gutierrez, E.; Leiva, A.; Casanello, P. Review: Differential placental macrovascular and microvascular endothelial dysfunction in gestational diabetes. Placenta 2011, 32 (Suppl. 2), S159–S164. [Google Scholar] [CrossRef]
- Treacy, A.; Higgins, M.; Kearney, J.M.; McAuliffe, F.; Mooney, E.E. Delayed villous maturation of the placenta: Quantitative assessment in different cohorts. Pediatr. Dev. Pathol. 2013, 16, 63–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacometti, C.; Cassaro, M. Clinicopathologic Correlations in Histopathologic Report of Placenta: Can We Do Something More? Arch. Pathol. Lab. Med. 2017, 141, 184–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.; Biswas, S.; Mitra, D.; Adhikari, A.; Saha, C. Histologic and morphometric study of human placenta in gestational diabetes mellitus. Ital. J. Anat. Embryol. 2014, 119, 1–9. [Google Scholar] [PubMed]
- Sala, M.A.; Matheus, M. Regional variation of the vasculo-syncytial membranes in the human full-term placenta. Gegenbaurs Morphol. Jahrb. 1986, 132, 285–289. [Google Scholar]
- Khong, T.Y.; Mooney, E.E.; Ariel, I.; Balmus, N.C.; Boyd, T.K.; Brundler, M.A.; Derricott, H.; Evans, M.J.; Faye-Petersen, O.M.; Gillan, J.E.; et al. Sampling and Definitions of Placental Lesions: Amsterdam Placental Workshop Group Consensus Statement. Arch. Pathol. Lab. Med. 2016, 140, 698–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, M.; McAuliffe, F.M.; Mooney, E.E. Clinical associations with a placental diagnosis of delayed villous maturation: A retrospective study. Pediatr. Dev. Pathol. 2011, 14, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Unek, G.; Ozmen, A.; Mendilcioglu, I.; Simsek, M.; Korgun, E.T. Immunohistochemical distribution of cell cycle proteins p27, p57, cyclin D3, PCNA and Ki67 in normal and diabetic human placentas. J. Mol. Histol. 2014, 45, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.H.; Huang, S.Y.; Chen, S.F.; Wu, C.P.; Hsieh, T.T. Decreased placental apoptosis and autophagy in pregnancies complicated by gestational diabetes with large-for-gestational age fetuses. Placenta 2020, 90, 27–36. [Google Scholar] [CrossRef]
- Tetzlaff, M.T.; Curry, J.L.; Ivan, D.; Wang, W.L.; Torres-Cabala, C.A.; Bassett, R.L.; Valencia, K.M.; McLemore, M.S.; Ross, M.I.; Prieto, V.G. Immunodetection of phosphohistone H3 as a surrogate of mitotic figure count and clinical outcome in cutaneous melanoma. Mod. Pathol. 2013, 26, 1153–1160. [Google Scholar] [CrossRef] [Green Version]
- Rossi, M.N.; Antonangeli, F. Cellular Response upon Stress: p57 Contribution to the Final Outcome. Mediat. Inflamm. 2015, 2015, 259325. [Google Scholar] [CrossRef] [Green Version]
- McCarty, K.S., Jr.; Miller, L.S.; Cox, E.B.; Konrath, J.; McCarty, K.S., Sr. Estrogen receptor analyses. Correlation of biochemical and immunohistochemical methods using monoclonal antireceptor antibodies. Arch. Pathol. Lab. Med. 1985, 109, 716–721. [Google Scholar] [PubMed]
- Langston, C.; Kaplan, C.; Macpherson, T.; Manci, E.; Peevy, K.; Clark, B.; Murtagh, C.; Cox, S.; Glenn, G. Practice guideline for examination of the placenta: Developed by the Placental Pathology Practice Guideline Development Task Force of the College of American Pathologists. Arch. Pathol. Lab. Med. 1997, 121, 449–476. [Google Scholar]
- Benhalima, K.; Van Crombrugge, P.; Moyson, C.; Verhaeghe, J.; Vandeginste, S.; Verlaenen, H.; Vercammen, C.; Maes, T.; Dufraimont, E.; De Block, C.; et al. Risk factor screening for gestational diabetes mellitus based on the 2013 WHO criteria. Eur. J. Endocrinol. 2019, 180, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Subiabre, M.; Silva, L.; Villalobos-Labra, R.; Toledo, F.; Paublo, M.; Lopez, M.A.; Salsoso, R.; Pardo, F.; Leiva, A.; Sobrevia, L. Maternal insulin therapy does not restore foetoplacental endothelial dysfunction in gestational diabetes mellitus. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2987–2998. [Google Scholar] [CrossRef]
- Farias, M.; San Martin, R.; Puebla, C.; Pearson, J.D.; Casado, J.F.; Pastor-Anglada, M.; Casanello, P.; Sobrevia, L. Nitric oxide reduces adenosine transporter ENT1 gene (SLC29A1) promoter activity in human fetal endothelium from gestational diabetes. J. Cell. Physiol. 2006, 208, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Casanello, P.; Escudero, C.; Sobrevia, L. Equilibrative nucleoside (ENTs) and cationic amino acid (CATs) transporters: Implications in foetal endothelial dysfunction in human pregnancy diseases. Curr. Vasc. Pharmacol. 2007, 5, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Westermeier, F.; Puebla, C.; Vega, J.L.; Farias, M.; Escudero, C.; Casanello, P.; Sobrevia, L. Equilibrative nucleoside transporters in fetal endothelial dysfunction in diabetes mellitus and hyperglycaemia. Curr. Vasc. Pharmacol. 2009, 7, 435–449. [Google Scholar] [CrossRef]
- da Silva, C.G.; Jarzyna, R.; Specht, A.; Kaczmarek, E. Extracellular nucleotides and adenosine independently activate AMP-activated protein kinase in endothelial cells: Involvement of P2 receptors and adenosine transporters. Circ. Res. 2006, 98, e39–e47. [Google Scholar] [CrossRef] [Green Version]
- Coade, S.B.; Pearson, J.D. Metabolism of adenine nucleotides in human blood. Circ. Res. 1989, 65, 531–537. [Google Scholar] [CrossRef] [Green Version]
- Yoneyama, Y.; Sawa, R.; Suzuki, S.; Ishino, H.; Miura, A.; Kuwabara, Y.; Kuwajima, T.; Ito, N.; Kiyokawa, Y.; Otsubo, Y.; et al. Regulation of plasma adenosine levels in normal pregnancy. Gynecol. Obs. Investig. 2002, 53, 71–74. [Google Scholar] [CrossRef]
- Pardo, F.; Arroyo, P.; Salomon, C.; Westermeier, F.; Salsoso, R.; Saez, T.; Guzman-Gutierrez, E.; Leiva, A.; Sobrevia, L. Role of equilibrative adenosine transporters and adenosine receptors as modulators of the human placental endothelium in gestational diabetes mellitus. Placenta 2013, 34, 1121–1127. [Google Scholar] [CrossRef] [PubMed]
- Salomon, C.; Westermeier, F.; Puebla, C.; Arroyo, P.; Guzman-Gutierrez, E.; Pardo, F.; Leiva, A.; Casanello, P.; Sobrevia, L. Gestational diabetes reduces adenosine transport in human placental microvascular endothelium, an effect reversed by insulin. PLoS ONE 2012, 7, e40578. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, G.; Sanhueza, F.; Vasquez, R.; Gonzalez, M.; San Martin, R.; Casanello, P.; Sobrevia, L. Role of adenosine transport in gestational diabetes-induced L-arginine transport and nitric oxide synthesis in human umbilical vein endothelium. J. Physiol. 2004, 560, 111–122. [Google Scholar] [CrossRef]
- Higgins, M.F.; Russell, N.M.; Mooney, E.E.; McAuliffe, F.M. Clinical and ultrasound features of placental maturation in pre-gestational diabetic pregnancy. Early Hum. Dev. 2012, 88, 817–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, J.; Dawson, D.; Roberts, D.; Bentley-Lewis, R. A systematic review of placental pathology in maternal diabetes mellitus. Placenta 2015, 36, 101–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evers, I.M.; Nikkels, P.G.; Sikkema, J.M.; Visser, G.H. Placental pathology in women with type 1 diabetes and in a control group with normal and large-for-gestational-age infants. Placenta 2003, 24, 819–825. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variable | GDMd (Mean ± SD) | Non-GDM (Mean ± SD) | |
|---|---|---|---|
| Mothers | p | ||
| Age (yrs) | 34.62 ± 4.28 | 33.67 ± 4.69 | 0.48 |
| BMI (kg/m2) | |||
| Pre-pregnancy | 24.19 ± 4.85 | 23.98 ± 4.23 | 0.87 |
| At term | 27.77 ± 4.32 | 28.145 ± 4.55 | 0.78 |
| Systolic blood pressure (mmHg) | 118.75 ± 11.07 | 123.75 ± 18.14 | 0.25 |
| Diastolic blood pressure (mmHg) | 73.08 ± 9.32 | 74.67 ± 10.98 | 0.60 |
| OGTT (mg/dL) | Not performed | ||
| Basal glycemia | 86.04 ± 9.34 | - | |
| 1-h glycemia after glucose load | 162.08 ± 29.03 | - | |
| 2-h glycemia after glucose load | 153.87 ± 25.98 | - | |
| Newborns | |||
| Sex (female/male) | 12/12 | 13/11 | |
| Gestational age (weeks) | 39.05 ± 1.11 | 37.95 ± 2.49 | 0.056 |
| Birth weight (grams) | 3248.54 ± 481.66 | 2626.12 ± 568.67 | 0.0002 |
| Placenta/fetus ratio | 0.14 ± 0.02 | 0.14 ± 0.02 | 0.87 |
| APGAR score (5 min) | 9.83 ± 0.38 | 9.21 ± 0.97 | 0.007 |
| Deliveries | |||
| Type of delivery | |||
| Vaginal, spontaneous, n (%) | 10/24 (41.67%) | 11/24 (45.83%) | |
| Vaginal, induced, n (%) | 13/24 (54.17%) | 1/24 (4.16%) | |
| Caesarian, n (%) | 1/24 (4.16%) | 10/24 (41.67%) | |
| Operative, n (%) | 0/24 (0%) | 2/24 (8.34%) |
| Variable | GDMd (Mean ± SD) | Non-GDM (Mean ± SD) | p |
|---|---|---|---|
| Morphological findings | |||
| Weight (grams) | 477.54 ± 75.96 | 394.91 ± 120.96 | 0.007 |
| Diameter (cm) | |||
| Maximum | 17.29 ± 1.89 | 16.26 ± 1.86 | 0.07 |
| Minimum | 15.21 ± 1.61 | 14.10 ± 2.02 | 0.04 |
| Thickness (cm) | |||
| Maximum | 3.48 ± 0.59 | 3.41 ± 0.82 | 0.76 |
| Minimum | 1.73 ± 0.80 | 1.49 ± 0.78 | 0.30 |
| Cord length (cm) | 27.08 ± 7.38 | 30.85 ± 11.97 | 0.10 |
| Umbilical Cord Index (UCI, number of umbilical veins twists/cm) | 0.32 ± 0.12 | 0.32 ± 0.22 | 0.94 |
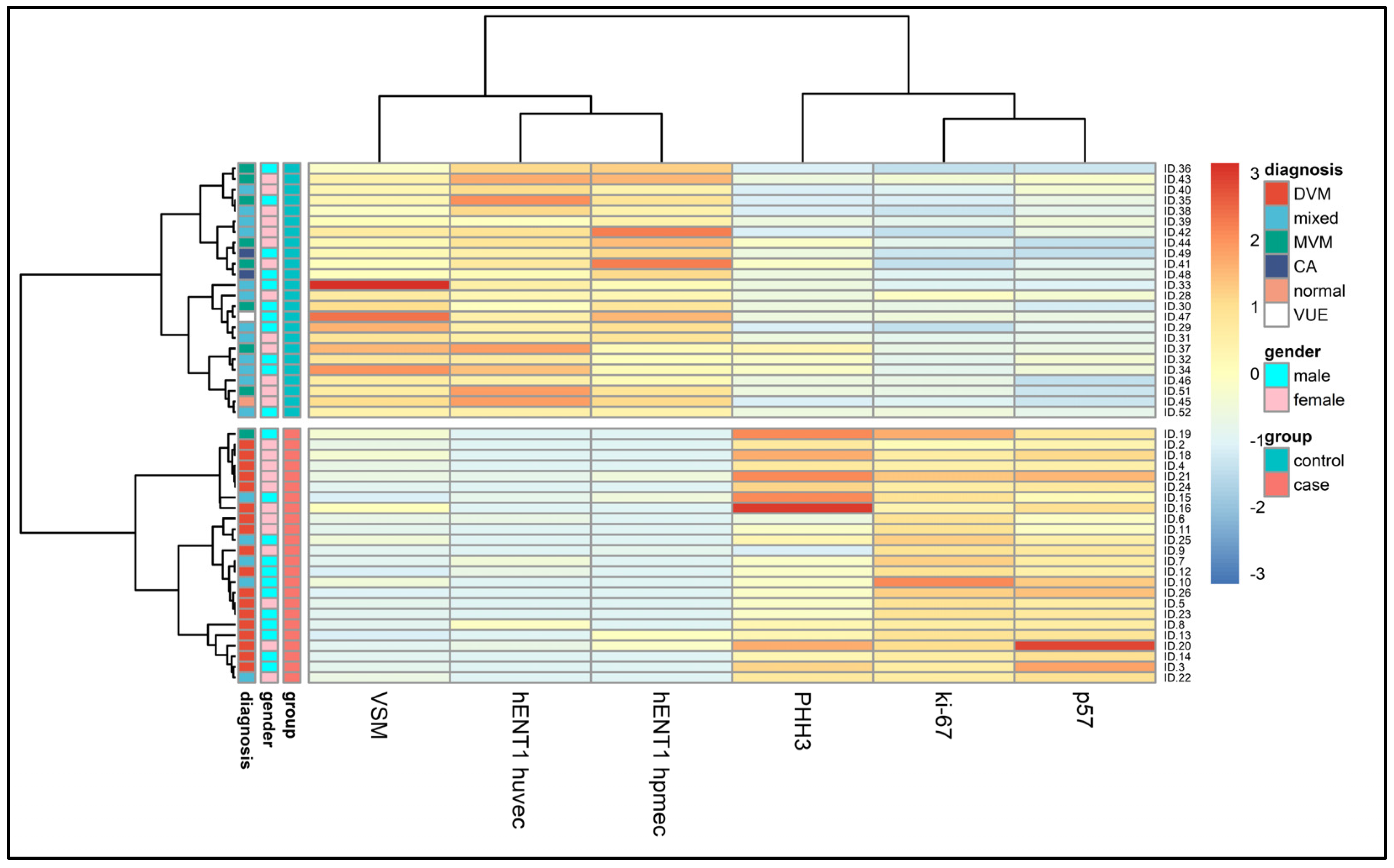
| Histological diagnosis | GDMd | Non-GDM | |
| Number of cases, % | DVM 20 (83.3%) | MVM 12 (50%) | |
| DVM + MVM 4 (16.7%) | MVM + DVM 8 (33%) | ||
| CA 2 (8.3%) | |||
| VUE 1 (4.2%) | |||
| Normal 1 (4.2%) | |||
| Immunohistochemical and morphological study | GDMd (mean ± SD) | Non-GDM (mean ± SD) | p |
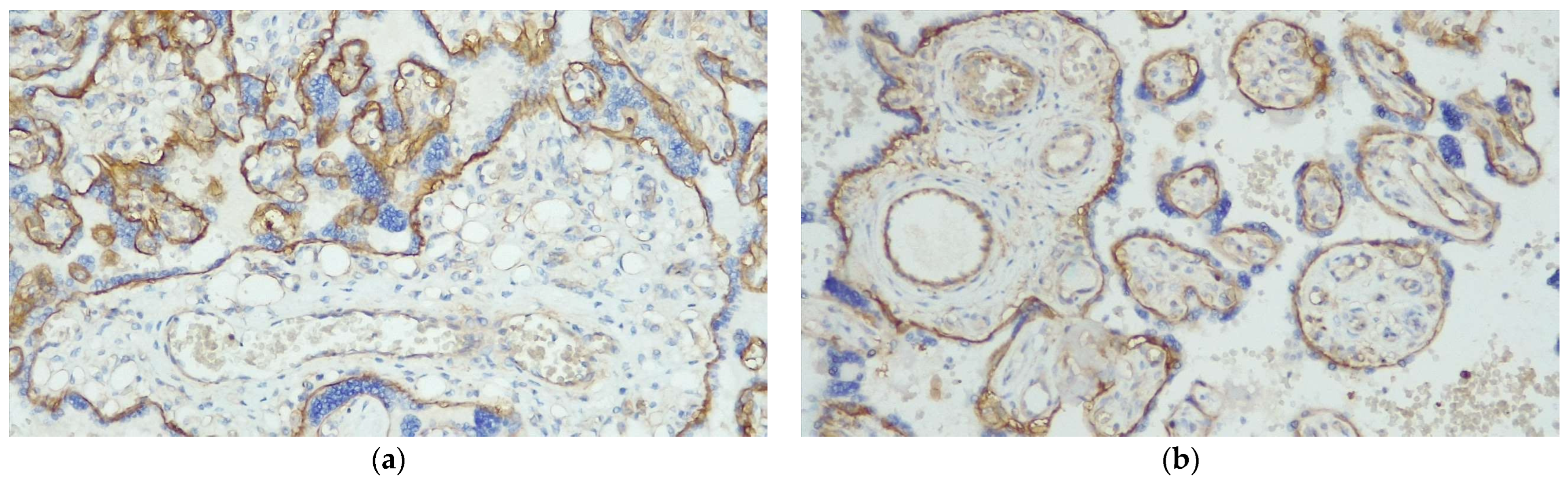
| hENT1 HUVEC | 11.25 ± 21.93 | 194.17 ± 66.52 | <0.0001 |
| hENT1 hPMEC | 10.00 ± 23.03 | 159.17 ± 54.76 | <0.0001 |
| Ki-67 | 19.96 ± 2.83 | 7.50 ± 2.37 | <0.0001 |
| PHH3 | 3.79 ± 2.26 | 0.96 ± 0.80 | <0.0001 |
| p57 | 20.87 ± 4.72 | 8.45 ± 2.84 | <0.0001 |
| VSM | 3.67 ± 2.96 | 20.04 ± 8.77 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giacometti, C.; Ludwig, K.; Guidi, M.; Colantuono, E.; Coracina, A.; Rigano, M.; Cassaro, M.; Ambrosi, A. Gestational Diabetes—Placental Expression of Human Equilibrative Nucleoside Transporter 1 (hENT1): Is Delayed Villous Maturation an Adaptive Pattern? Diagnostics 2023, 13, 2034. https://doi.org/10.3390/diagnostics13122034
Giacometti C, Ludwig K, Guidi M, Colantuono E, Coracina A, Rigano M, Cassaro M, Ambrosi A. Gestational Diabetes—Placental Expression of Human Equilibrative Nucleoside Transporter 1 (hENT1): Is Delayed Villous Maturation an Adaptive Pattern? Diagnostics. 2023; 13(12):2034. https://doi.org/10.3390/diagnostics13122034
Chicago/Turabian StyleGiacometti, Cinzia, Kathrin Ludwig, Monica Guidi, Elvira Colantuono, Anna Coracina, Marcello Rigano, Mauro Cassaro, and Alessandro Ambrosi. 2023. "Gestational Diabetes—Placental Expression of Human Equilibrative Nucleoside Transporter 1 (hENT1): Is Delayed Villous Maturation an Adaptive Pattern?" Diagnostics 13, no. 12: 2034. https://doi.org/10.3390/diagnostics13122034
APA StyleGiacometti, C., Ludwig, K., Guidi, M., Colantuono, E., Coracina, A., Rigano, M., Cassaro, M., & Ambrosi, A. (2023). Gestational Diabetes—Placental Expression of Human Equilibrative Nucleoside Transporter 1 (hENT1): Is Delayed Villous Maturation an Adaptive Pattern? Diagnostics, 13(12), 2034. https://doi.org/10.3390/diagnostics13122034