
The 1β-Hydroxy-Deoxycholic Acid to Deoxycholic Acid Urinary Metabolic Ratio: Toward a Phenotyping of CYP3A Using an Endogenous Marker?

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
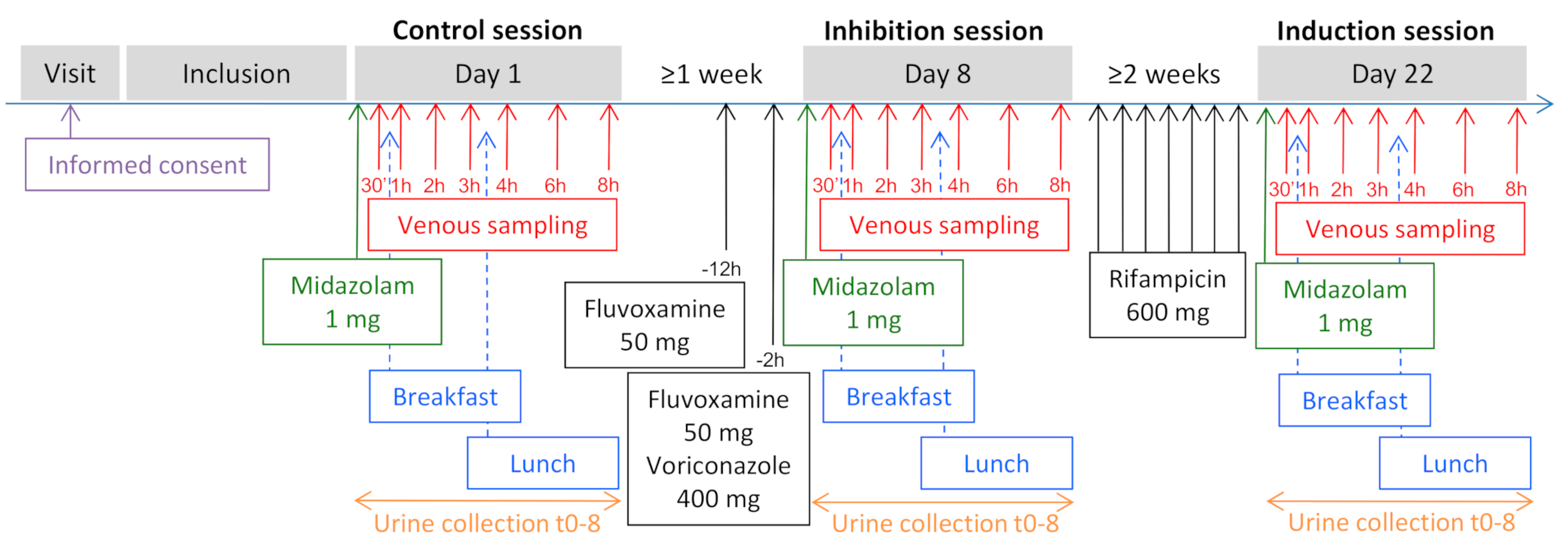
2.1. Study Design and Population
2.2. Quantification of Exogenous and Endogenous Probes
2.3. Statistical Analysis
- Oral midazolam clearance (oral MDZ CL)
- 1′-hydroxy-midazolam AUC0–8 to midazolam AUC0–8 ratio (AUC8 OH-MDZ/AUC8 MDZ)
- 1′-hydroxy-midazolam to midazolam metabolic ratio at 1 and 2 h (OH-MDZ/MDZ MR)
- Midazolam maximum blood concentration (MDZ Cmax).
3. Results
3.1. Subjects
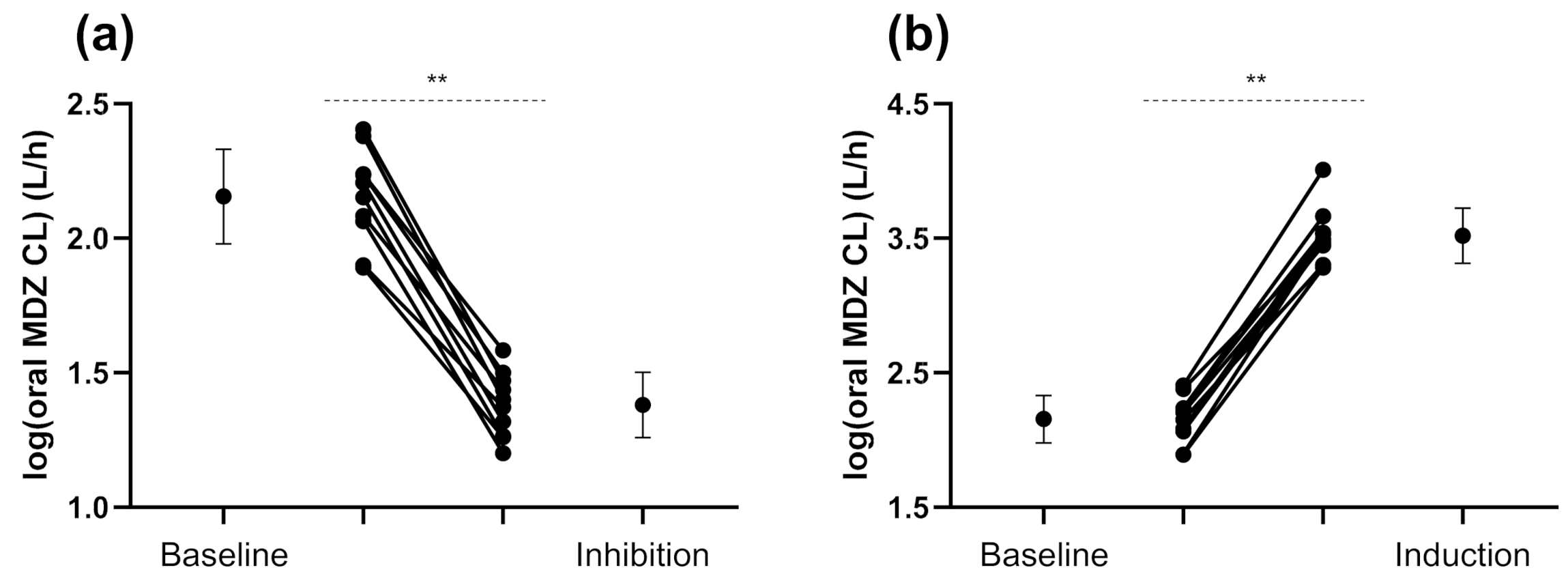
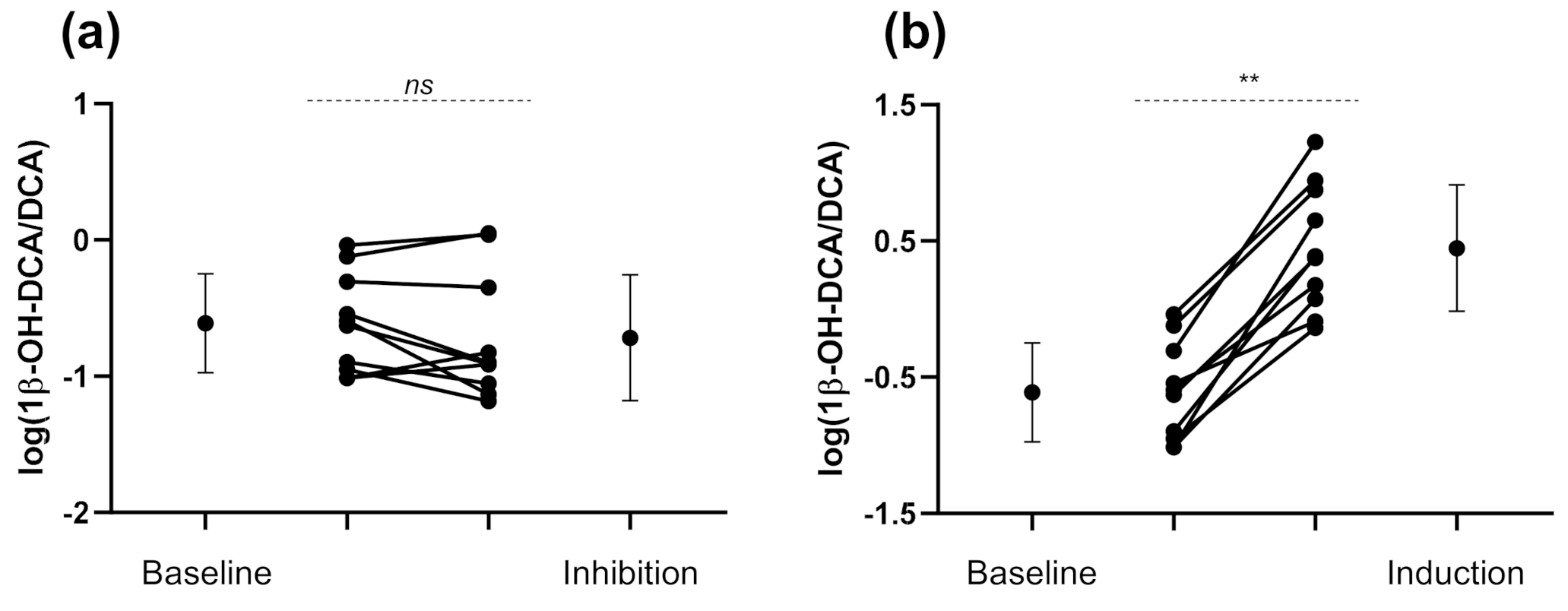
3.2. Exogenous and Endogenous CYP3A Phenotyping Metrics
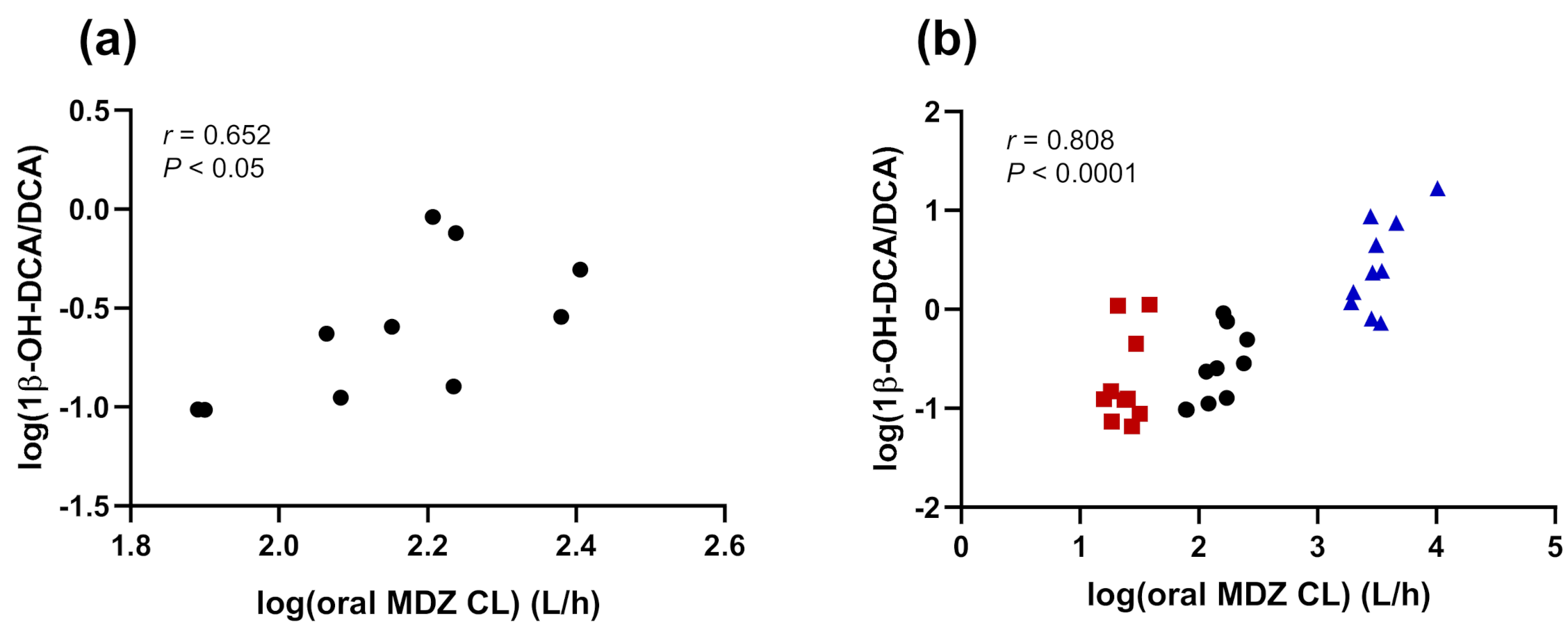
3.3. Correlations
4. Discussion
4.1. CYP3A Induction
4.2. CYP3A Inhibition
4.3. Correlations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zanger, U.M.; Schwab, M. Cytochrome P450 Enzymes in Drug Metabolism: Regulation of Gene Expression, Enzyme Activities, and Impact of Genetic Variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Tracy, T.S.; Chaudhry, A.S.; Prasad, B.; Thummel, K.E.; Schuetz, E.G.; Zhong, X.; Tien, Y.-C.; Jeong, H.; Pan, X.; Shireman, L.M.; et al. Interindividual Variability in Cytochrome P450–Mediated Drug Metabolism. Drug Metab. Dispos. 2016, 44, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Sadee, W. The Making of a CYP3A Biomarker Panel for Guiding Drug Therapy. J. Pers. Med. 2012, 2, 175–191. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Price, E.T.; Chang, C.-W.; Li, Y.; Huang, Y.; Guo, L.-W.; Guo, Y.; Kaput, J.; Shi, L.; Ning, B. Gene Expression Variability in Human Hepatic Drug Metabolizing Enzymes and Transporters. PLoS ONE 2013, 8, e60368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samer, C.F.; Lorenzini, K.I.; Rollason, V.; Daali, Y.; Desmeules, J.A. Applications of CYP450 Testing in the Clinical Setting. Mol. Diagn. Ther. 2013, 17, 165–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.; Mak, V.W.L.; Tomlinson, B. Simvastatin-Induced Myopathy, the Role of Interaction with Diltiazem and Genetic Predisposition. J. Clin. Pharm. 2011, 36, 419–425. [Google Scholar] [CrossRef]
- Lee, A.J.; Maddix, D.S. Rhabdomyolysis Secondary to a Drug Interaction between Simvastatin and Clarithromycin. Ann. Pharm. 2001, 35, 26–31. [Google Scholar] [CrossRef]
- Marot, A.; Morelle, J.; Chouinard, V.A.; Jadoul, M.; Lambert, M.; Demoulin, N. Concomitant Use of Simvastatin and Amiodarone Resulting in Severe Rhabdomyolysis: A Case Report and Review of the Literature. Acta Clin. Belg. 2011, 66, 134–136. [Google Scholar] [CrossRef]
- McGraw, J.; Gerhardt, A.; Morris, T.C. Opportunities and Obstacles in Genotypic Prediction of Cytochrome P450 Phenotypes. Expert Opin. Drug Metab. Toxicol. 2018, 14, 659–661. [Google Scholar] [CrossRef]
- Miao, J.; Jin, Y.; Marunde, R.L.; Kim, S.; Quinney, S.; Radovich, M.; Li, L.; Hall, S.D. Association of Genotypes of the CYP3A Cluster with Midazolam Disposition In Vivo. Pharm. J. 2009, 9, 319. [Google Scholar] [CrossRef] [Green Version]
- Kurnik, D.; Wood, A.J.J.; Wilkinson, G.R. The Erythromycin Breath Test Reflects P-Glycoprotein Function Independently of Cytochrome P450 3A Activity. Clin. Pharmacol. Ther. 2006, 80, 228–234. [Google Scholar] [CrossRef]
- Sugiyama, E.; Kikuchi, A.; Inada, M.; Sato, H. The Use of 13C–Erythromycin as an in Vivo Probe to Evaluate CYP3A-Mediated Drug Interactions in Rats. J. Pharm. Sci. 2011, 100, 3995–4005. [Google Scholar] [CrossRef] [PubMed]
- Christians, U.; Schmitz, V.; Haschke, M. Functional Interactions between P-Glycoprotein and CYP3A in Drug Metabolism. Expert Opin. Drug Metab. Toxicol. 2005, 1, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Rollason, V.; Mouterde, M.; Daali, Y.; Čížková, M.; Priehodová, E.; Kulichová, I.; Posová, H.; Petanová, J.; Mulugeta, A.; Makonnen, E.; et al. Safety of the Geneva Cocktail, a Cytochrome P450 and P-Glycoprotein Phenotyping Cocktail, in Healthy Volunteers from Three Different Geographic Origins. Drug Saf. 2020, 43, 1181–1189. [Google Scholar] [CrossRef]
- Magliocco, G.; Thomas, A.; Desmeules, J.; Daali, Y. Phenotyping of Human CYP450 Enzymes by Endobiotics: Current Knowledge and Methodological Approaches. Clin. Pharm. 2019, 58, 1373–1391. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Martin, I.; McLeod, J.; Nolan, G.; van Horn, R.; Vourvahis, M.; Lin, Y.S. Perspective: 4β-Hydroxycholesterol as an Emerging Endogenous Biomarker of Hepatic CYP3A. Drug Metab. Rev. 2017, 49, 18–34. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Lee, J.; Shin, K.-H.; Lee, S.; Yu, K.-S.; Jang, I.-J.; Cho, J.-Y. Identification of ω- or (ω-1)-Hydroxylated Medium-Chain Acylcarnitines as Novel Urinary Biomarkers for CYP3A Activity. Clin. Pharmacol. Ther. 2018, 103, 879–887. [Google Scholar] [CrossRef]
- Lee, J.; Kim, A.H.; Yi, S.; Lee, S.; Yoon, S.H.; Yu, K.-S.; Jang, I.-J.; Cho, J.-Y. Distribution of Exogenous and Endogenous CYP3A Markers and Related Factors in Healthy Males and Females. AAPS J 2017, 19, 1196–1204. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, K.-N.; Chen, C. The Role of CYP3A4 in the Biotransformation of Bile Acids and Therapeutic Implication for Cholestasis. Ann. Transl. Med. 2014, 2. [Google Scholar] [CrossRef]
- Bodin, K.; Lindbom, U.; Diczfalusy, U. Novel Pathways of Bile Acid Metabolism Involving CYP3A4. Biochim. Biophys. Acta 2005, 1687, 84–93. [Google Scholar] [CrossRef]
- Hayes, M.A.; Li, X.-Q.; Grönberg, G.; Diczfalusy, U.; Andersson, T.B. CYP3A Specifically Catalyzes 1β-Hydroxylation of Deoxycholic Acid: Characterization and Enzymatic Synthesis of a Potential Novel Urinary Biomarker for CYP3A Activity. Drug Metab. Dispos. 2016, 44, 1480–1489. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-J.; Zhang, J.; Zhu, P.-P.; Tan, X.-W.; Lin, Q.-H.; Wang, W.-X.; Yin, S.-S.; Gao, L.-Z.; Su, M.-M.; Liu, C.-X.; et al. Stereoselective Oxidation Kinetics of Deoxycholate in Recombinant and Microsomal CYP3A Enzymes: Deoxycholate 19-Hydroxylation Is an In Vitro Marker of CYP3A7 Activity. Drug Metab. Dispos. 2019, 47, 574–581. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, L.-Z.; Chen, Y.-J.; Zhu, P.-P.; Yin, S.-S.; Su, M.-M.; Ni, Y.; Miao, J.; Wu, W.-L.; Chen, H.; et al. Continuum of Host-Gut Microbial Co-Metabolism: Host CYP3A4/3A7 Are Responsible for Tertiary Oxidations of Deoxycholate Species. Drug Metab. Dispos. 2019, 47, 283–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosilkovska, M.; Samer, C.F.; Déglon, J.; Rebsamen, M.; Staub, C.; Dayer, P.; Walder, B.; Desmeules, J.A.; Daali, Y. Geneva Cocktail for Cytochrome P450 and P-Glycoprotein Activity Assessment Using Dried Blood Spots. Clin. Pharmacol. Ther. 2014, 96, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Bosilkovska, M.; Déglon, J.; Samer, C.; Walder, B.; Desmeules, J.; Staub, C.; Daali, Y. Simultaneous LC-MS/MS Quantification of P-Glycoprotein and Cytochrome P450 Probe Substrates and Their Metabolites in DBS and Plasma. Bioanalysis 2014, 6, 151–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, J.D.; Strauss, K.A.; Caplan, Y.H.; LoDico, C.P.; Bush, D.M. Urine PH: The Effects of Time and Temperature after Collection. J. Anal. Toxicol. 2007, 31, 486–496. [Google Scholar] [CrossRef] [Green Version]
- Thakare, R.; Chhonker, Y.S.; Gautam, N.; Alamoudi, J.A.; Alnouti, Y. Quantitative Analysis of Endogenous Compounds. J. Pharm. Biomed. Anal. 2016, 128, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Sarafian, M.H.; Lewis, M.R.; Pechlivanis, A.; Ralphs, S.; McPhail, M.J.W.; Patel, V.C.; Dumas, M.-E.; Holmes, E.; Nicholson, J.K. Bile Acid Profiling and Quantification in Biofluids Using Ultra-Performance Liquid Chromatography Tandem Mass Spectrometry. Anal. Chem. 2015, 87, 9662–9670. [Google Scholar] [CrossRef] [Green Version]
- Scherer, M.; Gnewuch, C.; Schmitz, G.; Liebisch, G. Rapid Quantification of Bile Acids and Their Conjugates in Serum by Liquid Chromatography-Tandem Mass Spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2009, 877, 3920–3925. [Google Scholar] [CrossRef]
- Ulaszewska, M.M.; Mancini, A.; Garcia-Aloy, M.; Del Bubba, M.; Tuohy, K.M.; Vrhovsek, U. Isotopic Dilution Method for Bile Acid Profiling Reveals New Sulfate Glycine-Conjugated Dihydroxy Bile Acids and Glucuronide Bile Acids in Serum. J. Pharm. Biomed. Anal. 2019, 173, 1–17. [Google Scholar] [CrossRef]
- Kanebratt, K.P.; Diczfalusy, U.; Bäckström, T.; Sparve, E.; Bredberg, E.; Böttiger, Y.; Andersson, T.B.; Bertilsson, L. Cytochrome P450 Induction by Rifampicin in Healthy Subjects: Determination Using the Karolinska Cocktail and the Endogenous CYP3A4 Marker 4β-Hydroxycholesterol. Clin. Pharmacol. Ther. 2008, 84, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, J. Bile Acid Metabolism and Circadian Rhythms. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G549–G563. [Google Scholar] [CrossRef]
- Li, T.; Apte, U. Bile Acid Metabolism and Signaling in Cholestasis, Inflammation and Cancer. Adv. Pharm. 2015, 74, 263–302. [Google Scholar] [CrossRef] [Green Version]
- Setchell, K.D.; Lawson, A.M.; Blackstock, E.J.; Murphy, G.M. Diurnal Changes in Serum Unconjugated Bile Acids in Normal Man. Gut 1982, 23, 637–642. [Google Scholar] [CrossRef] [Green Version]
- Galteau, M.M.; Shamsa, F. Urinary 6beta-Hydroxycortisol: A Validated Test for Evaluating Drug Induction or Drug Inhibition Mediated through CYP3A in Humans and in Animals. Eur. J. Clin. Pharmacol. 2003, 59, 713–733. [Google Scholar] [CrossRef]
- Shibasaki, H.; Hosoda, K.; Goto, M.; Suzuki, A.; Yokokawa, A.; Ishii, K.; Furuta, T. Intraindividual and Interindividual Variabilities in Endogenous Cortisol 6β-Hydroxylation Clearance as an Index for in Vivo CYP3A Phenotyping in Humans. Drug Metab. Dispos. 2013, 41, 475–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, K.-H.; Choi, M.H.; Lim, K.S.; Yu, K.-S.; Jang, I.-J.; Cho, J.-Y. Evaluation of Endogenous Metabolic Markers of Hepatic CYP3A Activity Using Metabolic Profiling and Midazolam Clearance. Clin. Pharmacol. Ther. 2013, 94, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Kasichayanula, S.; Boulton, D.W.; Luo, W.-L.; Rodrigues, A.D.; Yang, Z.; Goodenough, A.; Lee, M.; Jemal, M.; LaCreta, F. Validation of 4β-Hydroxycholesterol and Evaluation of Other Endogenous Biomarkers for the Assessment of CYP3A Activity in Healthy Subjects. Br. J. Clin. Pharm. 2014, 78, 1122–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemeryck, A.; Belpaire, F.M. Selective Serotonin Reuptake Inhibitors and Cytochrome P-450 Mediated Drug-Drug Interactions: An Update. Curr. Drug Metab. 2002, 3, 13–37. [Google Scholar] [CrossRef]
- Saari, T.I.; Laine, K.; Leino, K.; Valtonen, M.; Neuvonen, P.J.; Olkkola, K.T. Effect of Voriconazole on the Pharmacokinetics and Pharmacodynamics of Intravenous and Oral Midazolam. Clin. Pharm. Ther. 2006, 79, 362–370. [Google Scholar] [CrossRef]
- Greenblatt, D.J. Elimination Half-Life of Drugs: Value and Limitations. Annu. Rev. Med. 1985, 36, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Taves, M.D.; Gomez-Sanchez, C.E.; Soma, K.K. Extra-Adrenal Glucocorticoids and Mineralocorticoids: Evidence for Local Synthesis, Regulation, and Function. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E11–E24. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Li, X.; Hu, Z.; Cheng, Z. Evaluation of CYP3A Activity in Humans Using Three Different Parameters Based on Endogenous Cortisol Metabolism. Acta Pharmacol. Sin. 2009, 30, 1323–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penzak, S.R.; Rojas-Fernandez, C. 4β-Hydroxycholesterol as an Endogenous Biomarker for CYP3A Activity: Literature Review and Critical Evaluation. J. Clin. Pharmacol. 2019, 59, 611–624. [Google Scholar] [CrossRef] [Green Version]
- Tomalik-Scharte, D.; Lütjohann, D.; Doroshyenko, O.; Frank, D.; Jetter, A.; Fuhr, U. Plasma 4beta-Hydroxycholesterol: An Endogenous CYP3A Metric? Clin. Pharmacol. Ther. 2009, 86, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gravel, S.; Chiasson, J.-L.; Gaudette, F.; Turgeon, J.; Michaud, V. Use of 4β-Hydroxycholesterol Plasma Concentrations as an Endogenous Biomarker of CYP3A Activity: Clinical Validation in Individuals With Type 2 Diabetes. Clin. Pharmacol. Ther. 2019, 106, 831–840. [Google Scholar] [CrossRef]
- Ticho, A.L.; Malhotra, P.; Dudeja, P.K.; Gill, R.K.; Alrefai, W.A. Intestinal Absorption of Bile Acids in Health and Disease. Compr. Physiol. 2019, 10, 21–56. [Google Scholar] [CrossRef]
- de Wildt, S.N.; Ito, S.; Koren, G. Challenges for Drug Studies in Children: CYP3A Phenotyping as Example. Drug Discov. Today 2009, 14, 6–15. [Google Scholar] [CrossRef]
- Elens, L.; van Gelder, T.; Hesselink, D.A.; Haufroid, V.; van Schaik, R.H.N. CYP3A4*22: Promising Newly Identified CYP3A4 Variant Allele for Personalizing Pharmacotherapy. Pharmacogenomics 2013, 14, 47–62. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | MRM | Collision Energy (V) | Declustering Potential (V) | Cell Exit Potential (V) |
|---|---|---|---|---|
| DCA | 391.2 → 391.2 | −20 | −160 | −17 |
| DCA-D4 | 395.2 → 395.2 | −20 | −160 | −17 |
| 1β-OH-DCA | 407.2 → 407. 2 | −18 | −200 | −29 |
| 1β-OH-DCA-D4 | 411.3 → 411.3 | −18 | −200 | −29 |
| Baseline | Inhibition | Inhibition Ratio | Induction | Induction Ratio | |
|---|---|---|---|---|---|
| UMR1β-OH-DCA/DCA | 0.25 (0.13–0.45) | 0.19 (0.09–0.41) | 0.78 (0.52–1.2) | 2.8 (1.3–6.0) | 11.4 ** (6.3–20.7) |
| Urinary DCA excretion (µg/8 h) | 63.3 (27.6–145.1) | 50.2 (22.7–110.8) | 0.79 (0.40–1.6) | 22.1 (9.2–52.8) | 0.35 * (0.13–0.92) |
| Urinary1β-OH-DCA excretion (µg/8 h) | 15.5 (5.8–41.3) | 9.6 (4.1–22.5) | 0.62 (0.26–1.5) | 61.8 (30.9–123.3) | 4.0 * (1.4–11.0) |
| Oral MDZ CL (L/h) | 143.0 (107.1–191.0) | 24.0 (19.7–29.3) | 0.17 ** (0.13–0.22) | 3296 (2351–4622) | 23.1 ** (17.3–30.7) |
| AUC8 OH-MDZ/AUC8 MDZ | 0.45 (0.36–0.58) | 0.17 (0.14–0.22) | 0.38 ** (0.30–0.47) | 2.4 (2.0–2.9) | 5.4 ** (4.2–6.8) |
| OH-MDZ/MDZ MR at 1 h | 0.47 (0.37–0.58) | 0.21 (0.17–0.27) | 0.45 ** (0.36–0.56) | 2.4 (2.0–2.8) | 5.1 ** (4.2–6.3) |
| OH-MDZ/MDZ MR at 2 h | 0.44 (0.33–0.59) | 0.22 (0.17–0.28) | 0.49 ** (0.38–0.65) | 2.6 (2.2–3.0) | 6.0 ** (4.3–8.3) |
| MDZ Cmax (ng/mL) | 3.1 (2.3–4.2) | 11.7 (9.2–14.9) | 3.7 ** (2.7–5.2) | 0.15 (0.10–0.23) | 0.05 ** (0.03–0.07) |
| Baseline | Inhibition | Induction | All Sessions | |||||
|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | |
| Oral MDZ CL (L/h) | 0.652 | 0.041 | 0.336 | 0.343 | 0.678 | 0.031 | 0.808 | <0.0001 |
| AUC8 OH-MDZ/AUC8 MDZ | 0.275 | 0.442 | 0.172 | 0.635 | 0.567 | 0.088 | 0.781 | <0.0001 |
| OH-MDZ/MDZ MR at 1 h | 0.331 | 0.351 | −0.040 | 0.914 | 0.720 | 0.019 | 0.786 | <0.0001 |
| OH-MDZ/MDZ MR at 2 h | 0.242 | 0.500 | 0.215 | 0.550 | 0.500 | 0.141 | 0.790 | <0.0001 |
| MDZ Cmax (ng/mL) | −0.652 | 0.041 | −0.491 | 0.150 | −0.456 | 0.185 | −0.821 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magliocco, G.; Desmeules, J.; Bosilkovska, M.; Thomas, A.; Daali, Y. The 1β-Hydroxy-Deoxycholic Acid to Deoxycholic Acid Urinary Metabolic Ratio: Toward a Phenotyping of CYP3A Using an Endogenous Marker? J. Pers. Med. 2021, 11, 150. https://doi.org/10.3390/jpm11020150
Magliocco G, Desmeules J, Bosilkovska M, Thomas A, Daali Y. The 1β-Hydroxy-Deoxycholic Acid to Deoxycholic Acid Urinary Metabolic Ratio: Toward a Phenotyping of CYP3A Using an Endogenous Marker? Journal of Personalized Medicine. 2021; 11(2):150. https://doi.org/10.3390/jpm11020150
Chicago/Turabian StyleMagliocco, Gaëlle, Jules Desmeules, Marija Bosilkovska, Aurélien Thomas, and Youssef Daali. 2021. "The 1β-Hydroxy-Deoxycholic Acid to Deoxycholic Acid Urinary Metabolic Ratio: Toward a Phenotyping of CYP3A Using an Endogenous Marker?" Journal of Personalized Medicine 11, no. 2: 150. https://doi.org/10.3390/jpm11020150
APA StyleMagliocco, G., Desmeules, J., Bosilkovska, M., Thomas, A., & Daali, Y. (2021). The 1β-Hydroxy-Deoxycholic Acid to Deoxycholic Acid Urinary Metabolic Ratio: Toward a Phenotyping of CYP3A Using an Endogenous Marker? Journal of Personalized Medicine, 11(2), 150. https://doi.org/10.3390/jpm11020150