
Circulating Free DNA and Its Emerging Role in Autoimmune Diseases

 ,
, 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Circulating Free DNA Characteristics and Clinical Interest
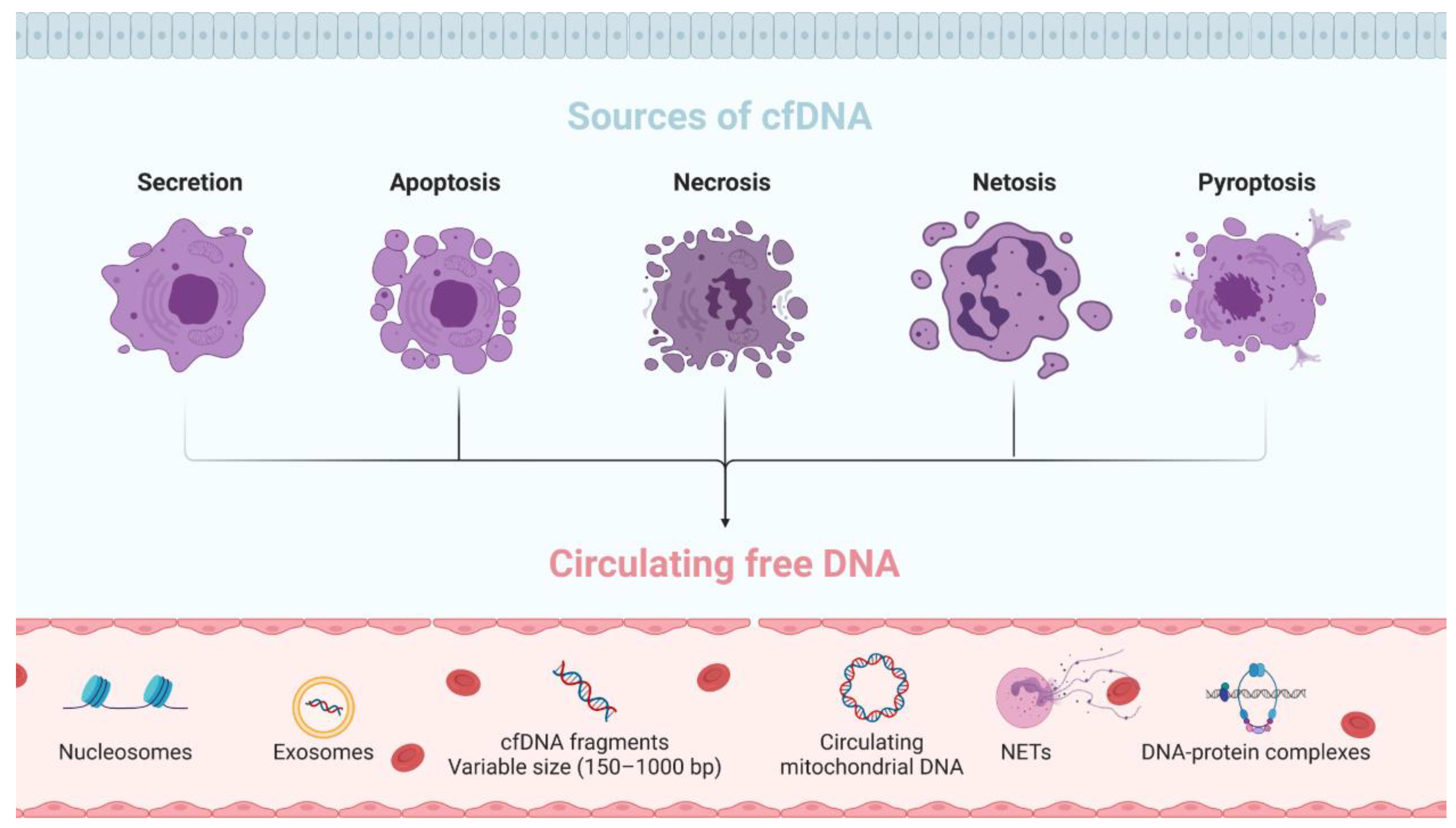
2.1. Origin and Characteristics
2.2. Clearance of cfDNA
2.3. Clinical Interest of cfDNA
3. Circulating Free DNA in Autoimmune Rheumatic Diseases
3.1. Systemic Lupus Erythaematosus
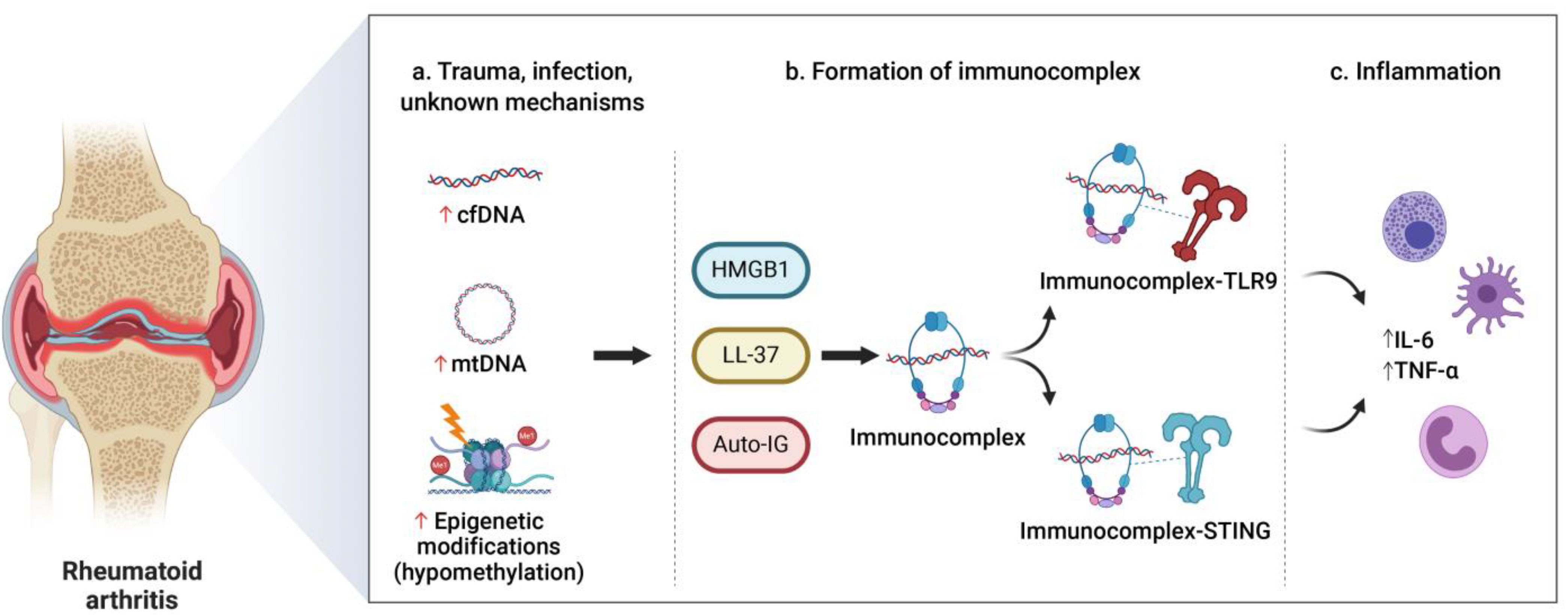
3.2. Rheumatoid Arthritis
3.3. Clinical Implication of cfDNA in Autoimmune Rheumatic Diseases
4. Circulating Free DNA in Inflammatory Bowel Disease
5. Circulating Free DNA in Other Autoimmune Disorders
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shoenfeld, Y.; Selmi, C.; Zimlichman, E.; Gershwin, M.E. The autoimmunologist: Geoepidemiology, a new center of gravity, and prime time for autoimmunity. J. Autoimmun. 2008, 31, 325–330. [Google Scholar] [CrossRef]
- Rose, N.R. Prediction and prevention of autoimmune disease in the 21st Century: A review and preview. Am. J. Epidemiol. 2016, 183, 403–406. [Google Scholar] [CrossRef]
- Gutierrez-Arcelus, M.; Rich, S.S.; Raychaudhuri, S. Autoimmune diseases-connecting risk alleles with molecular traits of the immune system. Nat. Rev. Genet. 2016, 17, 160–174. [Google Scholar] [CrossRef] [Green Version]
- Duncan, L.; Shen, H.; Gelaye, B.; Meijsen, J.; Ressler, K.; Feldman, M.; Peterson, R.; Domingue, B. Analysis of polygenic risk score usage and performance in diverse human populations. Nat. Commun. 2019, 10, 3328. [Google Scholar] [CrossRef] [PubMed]
- Rahat, B.; Ali, T.; Sapehia, D.; Mahajan, A.; Kaur, J. Circulating Cell-Free Nucleic Acids as Epigenetic Biomarkers in Precision Medicine. Front. Genet. 2020, 11, 844. [Google Scholar] [CrossRef]
- Pös, O.; Biró, O.; Szemes, T.; Nagy, B. Circulating cell-free nucleic acids: Characteristics and applications. Eur. J. Hum. Genet. 2018, 26, 937–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leon, S.A.; Shapiro, B.; Sklaroff, D.M.; Yaros, M.J. Free DNA in the serum of cancer patients and the effect of therapy. Cancer Res. 1977, 37, 646–650. [Google Scholar]
- Fleischhacker, M.; Schmidt, B. Circulating nucleic acids (CNAs) and cancer—A survey. Biochim. Biophys. Acta 2007, 1775, 181–232. [Google Scholar] [CrossRef]
- Duvvuri, B.; Lood, C. Cell-free DNA as a biomarker in autoimmune rheumatic diseases. Front. Immunol. 2019, 10, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szilágyi, M.; Pös, O.; Márton, É.; Buglyó, G.; Soltész, B.; Keserű, J.; Penyige, A.; Szemes, T.; Nagy, B. Circulating Cell-Free Nucleic Acids: Main Characteristics and Clinical Application. Int. J. Mol. Sci. 2020, 21, 6827. [Google Scholar] [CrossRef]
- Kustanovich, A.; Schwartz, R.; Peretz, T.; Grinshpun, A. Life and death of circulating cell-free DNA. Cancer Biol. Ther. 2019, 20, 1057–1067. [Google Scholar] [CrossRef] [Green Version]
- Aucamp, J.; Bronkhorst, A.J.; Badenhorst, C.P.S.; Pretorius, P.J. The diverse origins of circulating cell-free DNA in the human body: A critical re-evaluation of the literature. Biol. Rev. 2018, 93, 1649–1683. [Google Scholar] [CrossRef]
- Thierry, A.R.; El Messaoudi, S.; Gahan, P.B.; Anker, P.; Stroun, M. Origins, structures, and functions of circulating DNA in oncology. Cancer Metastasis Rev. 2016, 35, 347–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, M.R.; Jiang, C.; Krzyzanowski, G.D.; Ryan, W.L. New evidence that a large proportion of human blood plasma cell-free DNA is localized in exosomes. PLoS ONE 2017, 12, e0183915. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Xie, M.; Zhou, C.; Zheng, M. The relevance of pyroptosis in the pathogenesis of liver diseases. Life Sci. 2019, 223, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Tian, S.; Pan, Y.; Li, W.; Wang, Q.; Tang, Y.; Yu, T.; Wu, X.; Shi, Y.; Ma, P.; et al. Pyroptosis: A new frontier in cancer. Biomed. Pharmacother. Biomed. Pharmacother. 2020, 121, 109595. [Google Scholar] [CrossRef] [PubMed]
- Celec, P.; Vlková, B.; Lauková, L.; Bábíčková, J.; Boor, P. Cell-free DNA: The role in pathophysiology and as a biomarker in kidney diseases. Expert Rev. Mol. Med. 2018, 20, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Teo, Y.V.; Capri, M.; Morsiani, C.; Pizza, G.; Faria, A.M.C.; Franceschi, C.; Neretti, N. Cell-free DNA as a biomarker of aging. Aging Cell 2019, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Hummel, E.M.; Hessas, E.; Müller, S.; Beiter, T.; Fisch, M.; Eibl, A.; Wolf, O.T.; Giebel, B.; Platen, P.; Kumsta, R.; et al. Cell-free DNA release under psychosocial and physical stress conditions. Transl. Psychiatry 2018, 8, 236. [Google Scholar] [CrossRef] [Green Version]
- Vittori, L.N.; Tarozzi, A.; Latessa, P.M. Circulating cell-free DNA in physical activities. Methods Mol. Biol. 2019, 1909, 183–197. [Google Scholar]
- Zhong, S.; Ng, M.C.Y.; Lo, Y.M.D.; Chan, J.C.N.; Johnson, P.J. Presence of mitochondrial tRNA(Leu(UUR)) A to G 3243 mutation in DNA extracted from serum and plasma of patients with type 2 diabetes mellitus. J. Clin. Pathol. 2000, 53, 466–469. [Google Scholar] [CrossRef] [Green Version]
- Lowes, H.; Pyle, A.; Duddy, M.; Hudson, G. Cell-free mitochondrial DNA in progressive multiple sclerosis. Mitochondrion 2019, 46, 307–312. [Google Scholar] [CrossRef]
- Brodbeck, K.; Schick, S.; Bayer, B.; Anslinger, K.; Krüger, K.; Mayer, Z.; Holdenrieder, S.; Peldschus, S. Biological variability of cell-free DNA in healthy females at rest within a short time course. Int. J. Leg. Med. 2020, 134, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Meddeb, R.; Dache, Z.A.A.; Thezenas, S.; Otandault, A.; Tanos, R.; Pastor, B.; Sanchez, C.; Azzi, J.; Tousch, G.; Azan, S.; et al. Quantifying circulating cell-free DNA in humans. Sci. Rep. 2019, 9, 1–16. [Google Scholar]
- García Moreira, V.; De La Cera Martínez, T.; Gago González, E.; Prieto García, B.; Alvarez Menéndez, F.V. Increase in and clearance of cell-free plasma DNA in hemodialysis quantified by real-time PCR. Clin. Chem. Lab. Med. 2006, 44, 1410–1415. [Google Scholar] [CrossRef]
- Gaipl, U.S.; Beyer, T.D.; Heyder, P.; Kuenkele, S.; Böttcher, A.; Voll, R.E.; Kalden, J.R.; Herrmann, M. Cooperation between C1q and DNase I in the clearance of necrotic cell-derived chromatin. Arthritis Rheum. 2004, 50, 640–649. [Google Scholar] [CrossRef] [PubMed]
- Mandel, P.; Metais, P. Les acides nucléiques du plasma sanguin chez l’homme. Comptes Rendus Seances Soc. Biol. Fil. 1948, 142, 241–243. [Google Scholar]
- Tan, E.M.; Schur, P.H.; Carr, R.I.; Kunkel, H.G. Deoxybonucleic acid (DNA) and antibodies to DNA in the serum of patients with systemic lupus erythematosus. J. Clin. Investig. 1966, 45, 1732–1740. [Google Scholar] [CrossRef]
- Vasioukhin, V.; Anker, P.; Maurice, P.; Lyautey, J.; Lederrey, C.; Stroun, M. Point mutations of the N-ras gene in the blood plasma DNA of patients with myelodysplastic syndrome or acute myelogenous leukaemia. Br. J. Haematol. 1994, 86, 774–779. [Google Scholar] [CrossRef]
- Sorenson, G.D.; Pribish, D.M.; Valone, F.H.; Memoli, V.A.; Bzik, D.J.; Yao, S.L. Soluble normal and mutated DNA sequences from single-copy genes in human blood. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 1994, 3, 67–71. [Google Scholar]
- Rolfo, C.; Cardona, A.F.; Cristofanilli, M.; Paz-Ares, L.; Diaz Mochon, J.J.; Duran, I.; Raez, L.E.; Russo, A.; Lorente, J.A.; Malapelle, U.; et al. Challenges and opportunities of cfDNA analysis implementation in clinical practice: Perspective of the International Society of Liquid Biopsy (ISLB). Crit. Rev. Oncol. Hematol. 2020, 151, 102978. [Google Scholar] [CrossRef]
- Butler, T.M.; Spellman, P.T.; Gray, J. Circulating-tumor DNA as an early detection and diagnostic tool. Curr. Opin. Genet. Dev. 2017, 42, 14–21. [Google Scholar] [CrossRef]
- Diehl, F.; Schmidt, K.; Choti, M.A.; Romans, K.; Goodman, S.; Li, M.; Thornton, K.; Agrawal, N.; Sokoll, L.; Szabo, S.A.; et al. Circulating mutant DNA to assess tumor dynamics. Nat. Med. 2008, 14, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Vidal, J.; Muinelo, L.; Dalmases, A.; Jones, F.; Edelstein, D.; Iglesias, M.; Orrillo, M.; Abalo, A.; Rodríguez, C.; Brozos, E.; et al. Plasma ctDNA RAS mutation analysis for the diagnosis and treatment monitoring of metastatic colorectal cancer patients. Ann. Oncol. 2017, 28, 1325–1332. [Google Scholar] [CrossRef]
- Malapelle, U.; Sirera, R.; Jantus-Lewintre, E.; Reclusa, P.; Calabuig-Fariñas, S.; Blasco, A.; Pisapia, P.; Rolfo, C.; Camps, C. Profile of the Roche cobas® EGFR mutation test v2 for non-small cell lung cancer. Expert Rev. Mol. Diagn. 2017, 17, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Lamb, Y.N.; Dhillon, S. Epi proColon® 2.0 CE: A Blood-Based Screening Test for Colorectal Cancer. Mol. Diagn. Ther. 2017, 21, 225–232. [Google Scholar] [CrossRef]
- André, F.; Ciruelos, E.; Rubovszky, G.; Campone, M.; Loibl, S.; Rugo, H.S.; Iwata, H.; Conte, P.; Mayer, I.A.; Kaufman, B.; et al. Alpelisib for PIK3CA -Mutated, Hormone Receptor–Positive Advanced Breast Cancer. N. Engl. J. Med. 2019, 380, 1929–1940. [Google Scholar] [CrossRef] [PubMed]
- Laufer-Geva, S.; Rozenblum, A.B.; Twito, T.; Grinberg, R.; Dvir, A.; Soussan-Gutman, L.; Ilouze, M.; Roisman, L.C.; Dudnik, E.; Zer, A.; et al. The Clinical Impact of Comprehensive Genomic Testing of Circulating Cell-Free DNA in Advanced Lung Cancer. J. Thorac. Oncol. 2018, 13, 1705–1716. [Google Scholar] [CrossRef] [Green Version]
- Woodhouse, R.; Li, M.; Hughes, J.; Delfosse, D.; Skoletsky, J.; Ma, P.; Meng, W.; Dewal, N.; Milbury, C.; Clark, T.; et al. Clinical and analytical validation of FoundationOne Liquid CDx, a novel 324-Gene cfDNA-based comprehensive genomic profiling assay for cancers of solid tumor origin. PLoS ONE 2020, 15, e0237802. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.M.; Corbetta, N.; Chamberlain, P.F.; Rai, V.; Sargent, I.L.; Redman, C.W.; Wainscoat, J.S. Presence of fetal DNA in maternal plasma and serum. Lancet 1997, 350, 485–487. [Google Scholar] [CrossRef]
- Bianchi, D.W.; Chiu, R.W.K. Sequencing of Circulating Cell-free DNA during Pregnancy. N. Engl. J. Med. 2018, 379, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Isenberg, D.A.; Manson, J.J.; Ehrenstein, M.R.; Rahman, A. Fifty years of anti-ds DNA antibodies: Are we approaching journey’s end? Rheumatology 2007, 46, 1052–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, M.H.; Cizman, B.; Madaio, M.P. Nephritogenic autoantibodies in systemic lupus erythematosus: Immunochemical properties, mechanisms of immune deposition, and genetic origins. Lab. Investig. 1993, 69, 494–507. [Google Scholar]
- Pisetsky, D.S. The complex role of DNA, histones and HMGB1 in the pathogenesis of SLE. Autoimmunity 2014, 47, 487–493. [Google Scholar] [CrossRef]
- Pascual, V.; Farkas, L.; Banchereau, J. Systemic lupus erythematosus: All roads lead to type I interferons. Curr. Opin. Immunol. 2006, 18, 676–682. [Google Scholar] [CrossRef]
- Muller, S.; Dieker, J.; Tincani, A.; Meroni, P.L. Pathogenic anti-nucleosome antibodies. Lupus 2008, 17, 431–436. [Google Scholar] [CrossRef]
- Huang, H.; Evankovich, J.; Yan, W.; Nace, G.; Zhang, L.; Ross, M.; Liao, X.; Billiar, T.; Xu, J.; Esmon, C.T.; et al. Endogenous histones function as alarmins in sterile inflammatory liver injury through Toll-like receptor 9 in mice. Hepatology 2011, 54, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Willcox, G.; Mariotti, M.; Kaplan, C.; Lucotte, G. Characteristic parameters of the population of circulating DNA in cases of disseminated lupus erythematosus. Pathol. Biol. 1986, 34, 983–987. [Google Scholar]
- Raptis, L.; Menard, H.A. Quantitation and characterization of plasma DNA in normals and patients with systemic lupus erythematosus. J. Clin. Investig. 1980, 66, 1391–1399. [Google Scholar] [CrossRef] [Green Version]
- Klemp, P.; Meyers, O.L.; Harley, E.H. Measurement of plasma DNA by a physicochemical method: Relevance in SLE. Ann. Rheum. Dis. 1981, 40, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Compton, L.J.; Steinberg, A.D.; Sano, H. Nuclear DNA degradation in lymphocytes of patients with systemic lupus erythematosus. J. Immunol. 1984, 133, 213–216. [Google Scholar]
- Rumore, P.M.; Steinman, C.R. Endogenous circulating DNA in systemic lupus erythematosus. Occurrence as multimeric complexes bound to histone. J. Clin. Investig. 1990, 86, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Davis, G.L.; Davis Iv, J.S. Detection of circulating dna by counterimmunoelectrophoresis (cie). Arthritis Rheum. 1973, 16, 52–58. [Google Scholar] [CrossRef]
- Steinman, C.R. Circulating DNA in systemic lupus erythematosus. Association with central nervous system involvement and systemic vasculitis. Am. J. Med. 1979, 67, 429–435. [Google Scholar] [CrossRef]
- Chen, J.A.; Meister, S.; Urbonaviciute, V.; Rödel, F.; Wilhelm, S.; Kalden, J.R.; Manger, K.; Voll, R.E. Sensitive detection of plasma/serum DNA in patients with systemic lupus erythematosus. Autoimmunity 2007, 40, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Tobón, G.J.; Youinou, P.; Saraux, A. The environment, geo-epidemiology, and autoimmune disease: Rheumatoid arthritis. J. Autoimmun. 2010, 35, 10–14. [Google Scholar] [CrossRef]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O.; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef] [PubMed]
- Coras, R.; Narasimhan, R.; Guma, M. Liquid biopsies to guide therapeutic decisions in rheumatoid arthritis. Transl. Res. 2018, 201, 1–12. [Google Scholar] [CrossRef]
- Leon, S.A.; Ehrlich, G.E.; Shapiro, B.; Labbate, V.A. Free DNA in the serum of rheumatoid arthritis patients. J. Rheumatol. 1977, 4, 139–143. [Google Scholar] [PubMed]
- Zhong, X.-Y.; von Mühlenen, I.; Li, Y.; Kang, A.; Gupta, A.K.; Tyndall, A.; Holzgreve, W.; Hahn, S.; Hasler, P. Increased Concentrations of Antibody-Bound Circulatory Cell-Free DNA in Rheumatoid Arthritis. Clin. Chem. 2007, 53, 1609–1614. [Google Scholar] [CrossRef] [PubMed]
- Rykova, E.; Sizikov, A.; Roggenbuck, D.; Antonenko, O.; Bryzgalov, L.; Morozkin, E.; Skvortsova, K.; Vlassov, V.; Laktionov, P.; Kozlov, V. Circulating DNA in rheumatoid arthritis: Pathological changes and association with clinically used serological markers. Arthritis Res. Ther. 2017, 19, 85. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, T.; Yoshida, K.; Hashimoto, N.; Nakai, A.; Kaneshiro, K.; Suzuki, K.; Kawasaki, Y.; Shibanuma, N.; Hashiramoto, A. Circulating cell free DNA: A marker to predict the therapeutic response for biological DMARDs in rheumatoid arthritis. Int. J. Rheum. Dis. 2017, 20, 722–730. [Google Scholar] [CrossRef]
- Leon, S.A.; Revach, M.; Ehrlich, G.E.; Adler, R.; Petersen, V.; Shapiro, B. Dna in synovial fluid and the circulation of patients with arthritis. Arthritis Rheum. 1981, 24, 1142–1150. [Google Scholar] [CrossRef]
- Hajizadeh, S.; DeGroot, J.; TeKoppele, J.M.; Tarkowski, A.; Collins, L.V. Extracellular mitochondrial DNA and oxidatively damaged DNA in synovial fluid of patients with rheumatoid arthritis. Arthritis Res. Ther. 2003, 5, R234–R240. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Liu, Y.; Sun, C.; Liang, H.; Dai, L.; Shen, J.; Wei, S.; Guo, S.; Leong, K.W.; Chen, Y.; et al. Identification of Specific Joint-Inflammatogenic Cell-Free DNA Molecules From Synovial Fluids of Patients With Rheumatoid Arthritis. Front. Immunol. 2020, 11, 662. [Google Scholar] [CrossRef]
- Atamaniuk, J.; Hsiao, Y.-Y.; Mustak, M.; Bernhard, D.; Erlacher, L.; Fodinger, M.; Tiran, B.; Stuhlmeier, K.M. Analysing cell-free plasma DNA and SLE disease activity. Eur. J. Clin. Investig. 2011, 41, 579–583. [Google Scholar] [CrossRef]
- Xu, Y.; Song, Y.; Chang, J.; Zhou, X.; Qi, Q.; Tian, X.; Li, M.; Zeng, X.; Xu, M.; Zhang, W.; et al. High levels of circulating cell-free DNA are a biomarker of active SLE. Eur. J. Clin. Investig. 2018, 48, e13015. [Google Scholar] [CrossRef] [PubMed]
- Nuzzo, P.V.; Berchuck, J.E.; Korthauer, K.; Spisak, S.; Nassar, A.H.; Abou Alaiwi, S.; Chakravarthy, A.; Shen, S.Y.; Bakouny, Z.; Boccardo, F.; et al. Detection of renal cell carcinoma using plasma and urine cell-free DNA methylomes. Nat. Med. 2020, 26, 1041–1043. [Google Scholar] [CrossRef]
- Reyes-Thomas, J.; Blanco, I.; Putterman, C. Urinary Biomarkers in Lupus Nephritis. Clin. Rev. Allergy Immunol. 2011, 40, 138–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aragón, C.C.; Tafúr, R.-A.; Suárez-Avellaneda, A.; Martínez, M.T.; de las Salas, A.; Tobón, G.J. Urinary biomarkers in lupus nephritis. J. Transl. Autoimmun. 2020, 3, 100042. [Google Scholar] [CrossRef] [PubMed]
- Elkon, K.B. Review: Cell Death, Nucleic Acids, and Immunity. Arthritis Rheumatol. 2018, 70, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Cepika, A.-M.; Soldo Jureša, D.; Morović Vergles, J.; Malenica, B.; Šantak, M.; Kapitanović, S.; Mayer, M.; Anić, B.; Sentić, M.; Gagro, A. Decrease in circulating DNA, IL-10 and BAFF levels in newly-diagnosed SLE patients after corticosteroid and chloroquine treatment. Cell. Immunol. 2012, 276, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Bird, A.P. CpG-rich islands and the function of DNA methylation. Nature 1986, 321, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Aryee, M.J.; Padyukov, L.; Fallin, M.D.; Hesselberg, E.; Runarsson, A.; Reinius, L.; Acevedo, N.; Taub, M.; Ronninger, M.; et al. Epigenome-wide association data implicate DNA methylation as an intermediary of genetic risk in rheumatoid arthritis. Nat. Biotechnol. 2013, 31, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Lauková, L.; Konečná, B.; Vlková, B.; Mlynáriková, V.; Celec, P.; Šteňová, E. Anti-cytokine therapy and plasma DNA in patients with rheumatoid arthritis. Rheumatol. Int. 2018, 38, 1449–1454. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.; Plant, D.; Webster, A.P.; Barton, A. Genetic and genomic markers of anti-TNF treatment response in rheumatoid arthritis. Biomark. Med. 2015, 9, 499–512. [Google Scholar] [CrossRef]
- Birkelund, S.; Bennike, T.B.; Kastaniegaard, K.; Lausen, M.; Poulsen, T.B.G.; Kragstrup, T.W.; Deleuran, B.W.; Christiansen, G.; Stensballe, A. Proteomic analysis of synovial fluid from rheumatic arthritis and spondyloarthritis patients. Clin. Proteom. 2020, 17, 29. [Google Scholar] [CrossRef]
- Kubiritova, Z.; Radvanszky, J.; Gardlik, R. Cell-Free Nucleic Acids and their Emerging Role in the Pathogenesis and Clinical Management of Inflammatory Bowel Disease. Int. J. Mol. Sci. 2019, 20, 3662. [Google Scholar] [CrossRef] [Green Version]
- Podolsky, D.K. Inflammatory Bowel Disease. N. Engl. J. Med. 2002, 347, 417–429. [Google Scholar] [CrossRef]
- Casellas, F.; Antolín, M.; Varela, E.; García-Lafuente, A.; Guarner, F.; Borruel, N.; Armengol Miró, J.R.; Malagelada, J.R. Fecal excretion of human deoxyribonucleic acid as an index of inflammatory activity in ulcerative colitis. Clin. Gastroenterol. Hepatol. 2004, 2, 683–689. [Google Scholar] [CrossRef]
- Casellas, F.; Borruel, N.; Antolín, M.; Varela, E.; Torrejón, A.; Armadans, L.; Guarner, F.; Malagelada, J.-R. Fecal Excretion of Deoxyribonucleic Acid in Long-term Follow-up of Patients with Inactive Ulcerative Colitis. Inflamm. Bowel Dis. 2007, 13, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Vincent, C.; Mehrotra, S.; Loo, V.G.; Dewar, K.; Manges, A.R. Excretion of Host DNA in Feces Is Associated with Risk of Clostridium difficile Infection. J. Immunol. Res. 2015, 2015, 246203. [Google Scholar] [CrossRef] [Green Version]
- Koike, Y.; Uchida, K.; Tanaka, K.; Ide, S.; Otake, K.; Okita, Y.; Inoue, M.; Araki, T.; Mizoguchi, A.; Kusunoki, M. Dynamic pathology for circulating free DNA in a dextran sodium sulfate colitis mouse model. Pediatric Surg. Int. 2014, 30, 1199–1206. [Google Scholar] [CrossRef]
- Maronek, M.; Gromova, B.; Liptak, R.; Klimova, D.; Cechova, B.; Gardlik, R. Extracellular DNA is increased in dextran sulphate sodium-induced colitis in mice. Folia Biol. 2018, 64, 167–172. [Google Scholar]
- Geremia, A.; Biancheri, P.; Allan, P.; Corazza, G.R.; Di Sabatino, A. Innate and adaptive immunity in inflammatory bowel disease. Autoimmun. Rev. 2014, 13, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Peña, R.; Vidal-Castiñeira, J.R.; Moro-García, M.A.; Alonso-Arias, R.; Castro-Santos, P. Significant association of the KIR2DL3/HLA-C1 genotype with susceptibility to Crohn’s disease. Hum. Immunol. 2016, 77, 104–109. [Google Scholar] [CrossRef]
- Ii, W.A.R.; Sakamoto, K.; Leifer, C.A. TLR9 is important for protection against intestinal damage and for intestinal repair. Sci. Rep. 2012, 2, 574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obermeier, F.; Dunger, N.; Deml, L.; Herfarth, H.; Schölmerich, J.; Falk, W. CpG motifs of bacterial DNA exacerbate colitis of dextran sulfate sodium-treated mice. Eur. J. Immunol. 2002, 32, 2084–2092. [Google Scholar] [CrossRef]
- Jostins, L.; Ripke, S.; Weersma, R.K.; Duerr, R.H.; McGovern, D.P.; Hui, K.Y.; Lee, J.C.; Philip Schumm, L.; Sharma, Y.; Anderson, C.A.; et al. Host–microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 2012, 491, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Boyapati, R.K.; Dorward, D.A.; Tamborska, A.; Kalla, R.; Ventham, N.T.; Doherty, M.K.; Whitfield, P.D.; Gray, M.; Loane, J.; Rossi, A.G.; et al. Mitochondrial DNA Is a Pro-Inflammatory Damage-Associated Molecular Pattern Released During Active IBD. Inflamm. Bowel Dis. 2018, 24, 2113–2122. [Google Scholar] [CrossRef] [PubMed]
- Shimada, K.; Crother, T.R.; Karlin, J.; Dagvadorj, J.; Chiba, N.; Chen, S.; Ramanujan, V.K.; Wolf, A.J.; Vergnes, L.; Ojcius, D.M.; et al. Oxidized Mitochondrial DNA Activates the NLRP3 Inflammasome during Apoptosis. Immunity 2012, 36, 401–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coimbra, S.; Oliveira, H.; Reis, F.; Belo, L.; Rocha, S.; Quintanilha, A.; Figueiredo, A.; Teixeira, F.; Castro, E.; Rocha-Pereira, P.; et al. Circulating levels of adiponectin, oxidized LDL and C-reactive protein in Portuguese patients with psoriasis vulgaris, according to body mass index, severity and duration of the disease. J. Dermatol. Sci. 2009, 55, 202–204. [Google Scholar] [CrossRef] [PubMed]
- Beranek, M.; Fiala, Z.; Kremlacek, J.; Andrys, C.; Krejsek, J.; Hamakova, K.; Chmelarova, M.; Palicka, V.; Borska, L. Changes in circulating cell-free DNA and nucleosomes in patients with exacerbated psoriasis. Arch. Dermatol. Res. 2017, 309, 815–821. [Google Scholar] [CrossRef]
- Sakamoto, R.; Sawamura, S.; Kajihara, I.; Miyauchi, H.; Urata, K.; Otsuka-Maeda, S.; Kanemaru, H.; Kanazawa-Yamada, S.; Honda, N.; Makino, K.; et al. Circulating tumor necrosis factor-α DNA are elevated in psoriasis. J. Dermatol. 2020, 47, 1037–1040. [Google Scholar] [CrossRef]
- Bowcock, A.M.; Krueger, J.G. Getting under the skin: The immunogenetics of psoriasis. Nat. Rev. Immunol. 2005, 5, 699–711. [Google Scholar] [CrossRef]
- Brynychova, I.; Zinkova, A.; Hoffmanova, I.; Korabecna, M.; Dankova, P. Immunoregulatory properties of cell-free DNA in plasma of celiac disease patients—A pilot study. Autoimmunity 2019, 52, 88–94. [Google Scholar] [CrossRef]
- Flier, J.S.; Underhill, L.H.; Eisenbarth, G.S. Type I Diabetes Mellitus. N. Engl. J. Med. 1986, 314, 1360–1368. [Google Scholar] [CrossRef] [PubMed]
- Speake, C.; Ylescupidez, A.; Neiman, D.; Shemer, R.; Glaser, B.; Tersey, S.A.; Usmani-Brown, S.; Clark, P.; Wilhelm, J.J.; Bellin, M.D.; et al. Circulating Unmethylated Insulin DNA As a Biomarker of Human Beta Cell Death: A Multi-laboratory Assay Comparison. J. Clin. Endocrinol. Metab. 2020, 105, 781–791. [Google Scholar] [CrossRef]
- Lehmann-Werman, R.; Neiman, D.; Zemmour, H.; Moss, J.; Magenheim, J.; Vaknin-Dembinsky, A.; Rubertsson, S.; Nellgård, B.; Blennow, K.; Zetterberg, H.; et al. Identification of tissue-specific cell death using methylation patterns of circulating DNA. Proc. Natl. Acad. Sci. USA 2016, 113, E1826–E1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AlRashidi, F.T.; Gillespie, K.M. Biomarkers in Islet Cell Transplantation for Type 1 Diabetes. Curr. Diabetes Rep. 2018, 18, 94. [Google Scholar] [CrossRef] [Green Version]
- Gala-Lopez, B.L.; Neiman, D.; Kin, T.; O’Gorman, D.; Pepper, A.R.; Malcolm, A.J.; Pianzin, S.; Senior, P.A.; Campbell, P.; Glaser, B.; et al. Beta Cell Death by Cell-free DNA and Outcome After Clinical Islet Transplantation. Transplantation 2018, 102, 978–985. [Google Scholar] [CrossRef] [Green Version]
- Bartoloni, E.; Ludovini, V.; Alunno, A.; Pistola, L.; Bistoni, O.; Crinò, L.; Gerli, R. Increased levels of circulating DNA in patients with systemic autoimmune diseases: A possible marker of disease activity in Sjögren’s syndrome. Lupus 2011, 20, 928–935. [Google Scholar] [CrossRef]
- Vakrakou, A.G.; Boiu, S.; Ziakas, P.D.; Xingi, E.; Boleti, H.; Manoussakis, M.N. Systemic activation of NLRP3 inflammasome in patients with severe primary Sjögren’s syndrome fueled by inflammagenic DNA accumulations. J. Autoimmun. 2018, 91, 23–33. [Google Scholar] [CrossRef]
- Lublin, F.D.; Reingold, S.C. Defining the clinical course of multiple sclerosis: Results of an international survey. Neurology 1996, 46, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Varhaug, K.N.; Vedeler, C.A.; Myhr, K.-M.; Aarseth, J.H.; Tzoulis, C.; Bindoff, L.A. Increased levels of cell-free mitochondrial DNA in the cerebrospinal fluid of patients with multiple sclerosis. Mitochondrion 2017, 34, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Leurs, C.E.; Podlesniy, P.; Trullas, R.; Balk, L.; Steenwijk, M.D.; Malekzadeh, A.; Piehl, F.; Uitdehaag, B.M.; Killestein, J.; van Horssen, J.; et al. Cerebrospinal fluid mtDNA concentration is elevated in multiple sclerosis disease and responds to treatment. Mult. Scler. J. 2018, 24, 472–480. [Google Scholar] [CrossRef]
- Fissolo, N.; Cervera-Carles, L.; Villar Guimerans, L.M.; Lleó, A.; Clarimón, J.; Drulovic, J.; Dujmovic, I.; Voortman, M.; Khalil, M.; Gil, E.; et al. Cerebrospinal fluid mitochondrial DNA levels in patients with multiple sclerosis. Mult. Scler. J. 2019, 25, 1535–1538. [Google Scholar] [CrossRef]
- Liggett, T.; Melnikov, A.; Tilwalli, S.; Yi, Q.; Chen, H.; Replogle, C.; Feng, X.; Reder, A.; Stefoski, D.; Balabanov, R.; et al. Methylation patterns of cell-free plasma DNA in relapsing–remitting multiple sclerosis. J. Neurol. Sci. 2010, 290, 16–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, J.; Urnovitz, H.B.; Saresella, M.; Caputo, D.; Clerici, M.; Mitchell, W.M.; Schütz, E. Serum DNA Motifs Predict Disease and Clinical Status in Multiple Sclerosis. J. Mol. Diagn. 2010, 12, 312–319. [Google Scholar] [CrossRef]
- Dunaeva, M.; Derksen, M.; Pruijn, G.J.M. LINE-1 Hypermethylation in Serum Cell-Free DNA of Relapsing Remitting Multiple Sclerosis Patients. Mol. Neurobiol. 2018, 55, 4681–4688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dor, Y.; Cedar, H. Principles of DNA methylation and their implications for biology and medicine. Lancet 2018, 392, 777–786. [Google Scholar] [CrossRef]
- Schwarzenbach, H.; Hoon, D.S.B.; Pantel, K. Cell-free nucleic acids as biomarkers in cancer patients. Nat. Rev. Cancer 2011, 11, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Serra, E.G.; Schwerd, T.; Moutsianas, L.; Cavounidis, A.; Fachal, L.; Pandey, S.; Kammermeier, J.; Croft, N.M.; Posovszky, C.; Rodrigues, A.; et al. Somatic mosaicism and common genetic variation contribute to the risk of very-early-onset inflammatory bowel disease. Nat. Commun. 2020, 11, 995. [Google Scholar] [CrossRef] [Green Version]
- Olafsson, S.; McIntyre, R.E.; Coorens, T.; Butler, T.; Jung, H.; Robinson, P.S.; Lee-Six, H.; Sanders, M.A.; Arestang, K.; Dawson, C.; et al. Somatic Evolution in Non-neoplastic IBD-Affected Colon. Cell 2020, 182, 672–684.e11. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Wysocki, R.W.; Amoozgar, Z.; Maiorino, L.; Fein, M.R.; Jorns, J.; Schott, A.F.; Kinugasa-Katayama, Y.; Lee, Y.; Won, N.H.; et al. Cancer cells induce metastasis-supporting neutrophil extracellular DNA traps. Sci. Transl. Med. 2016, 8, 361ra138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauková, L.; Konečná, B.; Bábíčková, J.; Wagnerová, A.; Melišková, V.; Vlková, B.; Celec, P. Exogenous deoxyribonuclease has a protective effect in a mouse model of sepsis. Biomed. Pharmacother. 2017, 93, 8–16. [Google Scholar] [CrossRef]
- Stearns, N.A.; Lee, J.; Leong, K.W.; Sullenger, B.A.; Pisetsky, D.S. The inhibition of anti-DNA binding to DNA by nucleic acid binding polymers. PLoS ONE 2012, 7, e40862. [Google Scholar] [CrossRef]
- Dawulieti, J.; Sun, M.; Sun, M.; Zhao, Y.; Shao, D.; Shao, D.; Yan, H.; Lao, Y.H.; Hu, H.; Cui, L.; et al. Treatment of severe sepsis with nanoparticulate cell-free DNA scavengers. Sci. Adv. 2020, 6, eaay7148. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mondelo-Macía, P.; Castro-Santos, P.; Castillo-García, A.; Muinelo-Romay, L.; Diaz-Peña, R. Circulating Free DNA and Its Emerging Role in Autoimmune Diseases. J. Pers. Med. 2021, 11, 151. https://doi.org/10.3390/jpm11020151
Mondelo-Macía P, Castro-Santos P, Castillo-García A, Muinelo-Romay L, Diaz-Peña R. Circulating Free DNA and Its Emerging Role in Autoimmune Diseases. Journal of Personalized Medicine. 2021; 11(2):151. https://doi.org/10.3390/jpm11020151
Chicago/Turabian StyleMondelo-Macía, Patricia, Patricia Castro-Santos, Adrián Castillo-García, Laura Muinelo-Romay, and Roberto Diaz-Peña. 2021. "Circulating Free DNA and Its Emerging Role in Autoimmune Diseases" Journal of Personalized Medicine 11, no. 2: 151. https://doi.org/10.3390/jpm11020151
APA StyleMondelo-Macía, P., Castro-Santos, P., Castillo-García, A., Muinelo-Romay, L., & Diaz-Peña, R. (2021). Circulating Free DNA and Its Emerging Role in Autoimmune Diseases. Journal of Personalized Medicine, 11(2), 151. https://doi.org/10.3390/jpm11020151