
Cannabis Use Induces Distinctive Proteomic Alterations in Olfactory Neuroepithelial Cells of Schizophrenia Patients

 , , , , ,
, , , , , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Clinical and Neuropsychological Evaluation
2.3. Quantification of Cannabis Metabolites
2.4. Nasal Exfoliation and Cell Culture
2.5. Quantitative Proteomics in ON Cells
2.6. Pathway Analysis, Statistics and Bioinformatics
3. Results
3.1. Demographics and Neuropsychological Outcomes
3.2. Plasmatic Concentrations of THC-COOH
3.3. Proteomic Analyses
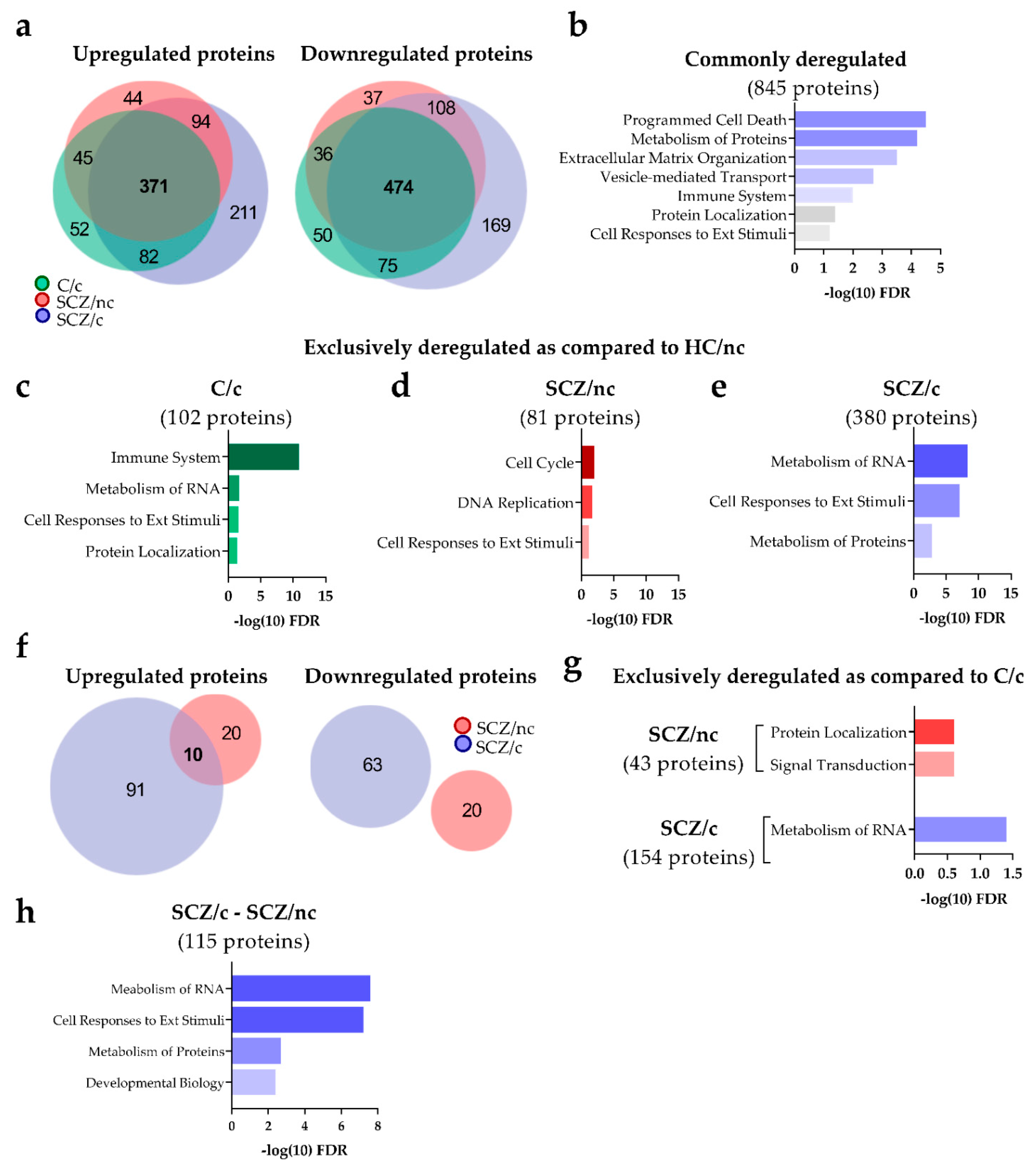
3.3.1. Quantitative and Functional Proteomic Profile of ON Cells from SCZ Patients as Compared to C/nc
3.3.2. Quantitative and Functional Proteomic Profile of ON Cells from SCZ Patients as Compared to C/nc
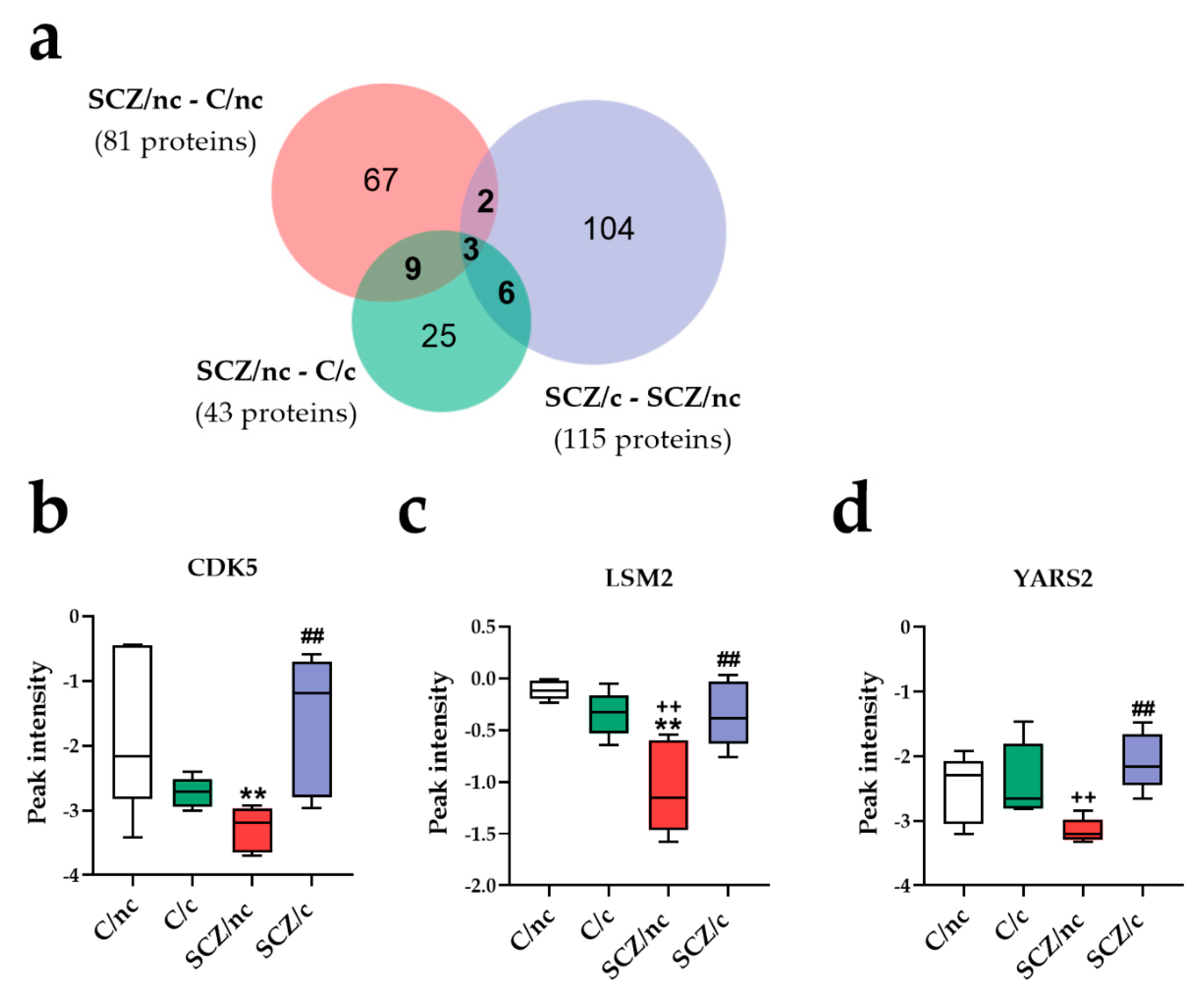
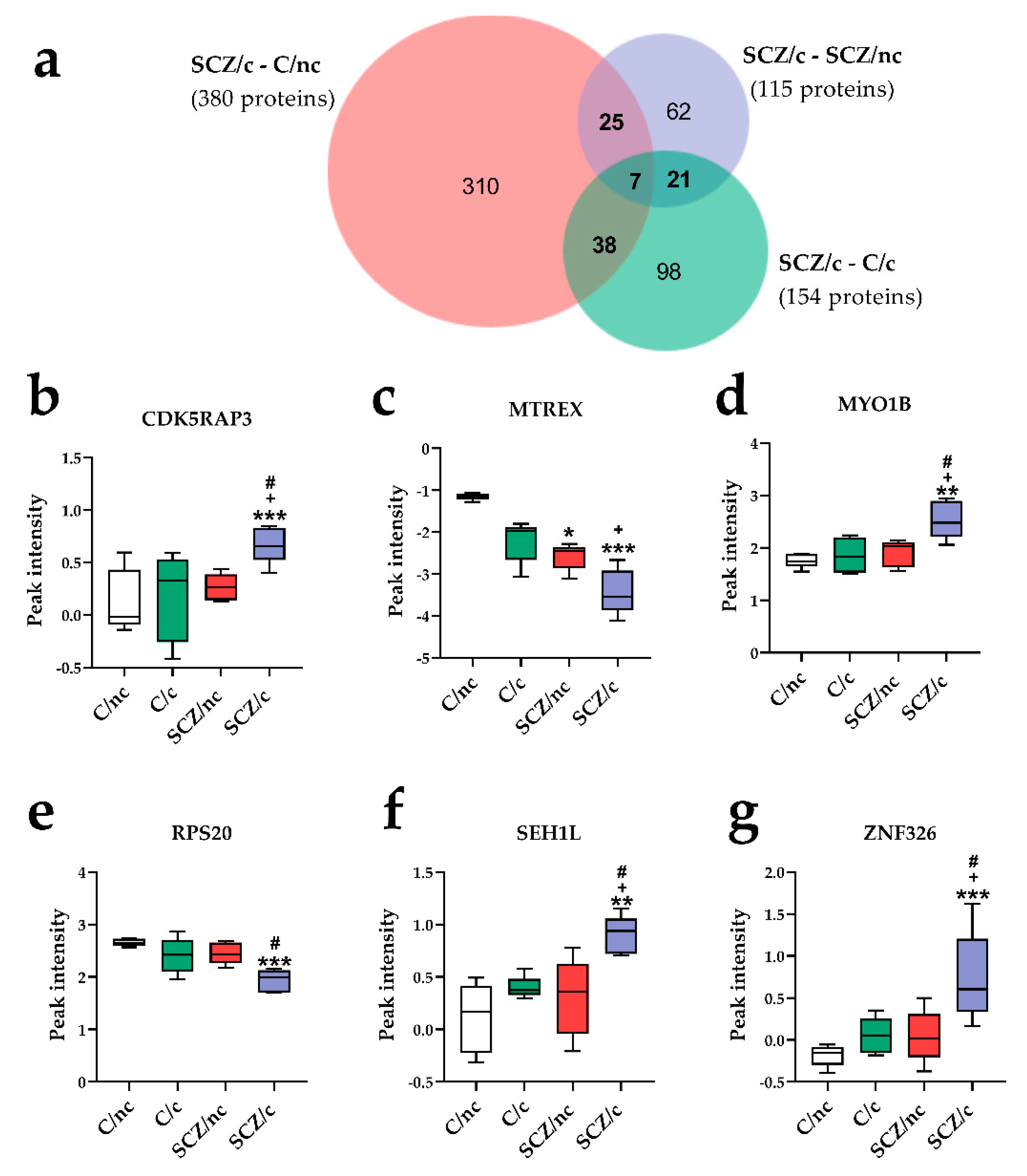
3.3.3. Specific Proteins Markers of SCZ Depending on Cannabis Use
3.4. Correlations between Specific Protein Markers, Cannabis Use and Cognitive Performance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Os, J.; Rutten, B.P.F.; Poulton, R. Gene-environment interactions in schizophrenia: Review of epidemiological findings and future directions. Schizophr. Bull. 2008, 34, 1066–1082. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, J.M.; Martins-De-Souza, D. The proteome of schizophrenia. NPJ Schizophr. 2015, 1, 14003. [Google Scholar] [CrossRef] [Green Version]
- Jaros, J.A.J.; Martins-de-Souza, D.; Rahmoune, H.; Rothermundt, M.; Leweke, F.M.; Guest, P.C.; Bahn, S. Protein phosphorylation patterns in serum from schizophrenia patients and healthy controls. J. Proteom. 2012, 76, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wan, C.; Li, H.; Zhu, H.; La, Y.; Xi, Z.; Chen, Y.; Jiang, L.; Feng, G.; He, L. Altered levels of acute phase proteins in the plasma of patients with schizophrenia. Anal. Chem. 2006, 78, 3571–3576. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.T.J.; Wang, L.; Prabakaran, S.; Wengenroth, M.; Lockstone, H.E.; Koethe, D.; Gerth, C.W.; Gross, S.; Schreiber, D.; Lilley, K.; et al. Independent protein-profiling studies show a decrease in apolipoprotein A1 levels in schizophrenia CSF, brain and peripheral tissues. Mol. Psychiatry 2008, 13, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Khandaker, G.M.; Dantzer, R. Is there a role for immune-to-brain communication in schizophrenia? Psychopharmacology 2016, 233, 1559–1573. [Google Scholar] [CrossRef] [Green Version]
- Boiko, A.S.; Mednova, I.A.; Kornetova, E.G.; Semke, A.V.; Bokhan, N.A.; Loonen, A.J.M.; Ivanova, S.A. Apolipoprotein serum levels related to metabolic syndrome in patients with schizophrenia. Heliyon 2019, 5, e02033. [Google Scholar] [CrossRef] [Green Version]
- Föcking, M.; Dicker, P.; English, J.A.; Schubert, K.O.; Dunn, M.J.; Cotter, D.R. Common proteomic changes in the hippocampus in schizophrenia and bipolar disorder and particular evidence for involvement of cornu ammonis regions 2 and 3. Arch. Gen. Psychiatry 2011, 68, 477–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins-de-Souza, D.; Maccarrone, G.; Wobrock, T.; Zerr, I.; Gormanns, P.; Reckow, S.; Falkai, P.; Schmitt, A.; Turck, C.W. Proteome analysis of the thalamus and cerebrospinal fluid reveals glycolysis dysfunction and potential biomarkers candidates for schizophrenia. J. Psychiatr. Res. 2010, 44, 1176–1189. [Google Scholar] [CrossRef]
- Wesseling, H.; Chan, M.K.; Tsang, T.M.; Ernst, A.; Peters, F.; Guest, P.C.; Holmes, E.; Bahn, S. A combined metabonomic and proteomic approach identifies frontal cortex changes in a chronic phencyclidine rat model in relation to human schizophrenia brain pathology. Neuropsychopharmacology 2013, 38, 2532–2544. [Google Scholar] [CrossRef] [Green Version]
- Beasley, C.L.; Pennington, K.; Behan, A.; Wait, R.; Dunn, M.J.; Cotter, D. Proteomic analysis of the anterior cingulate cortex in the major psychiatric disorders: Evidence for disease-associated changes. Proteomics 2006, 6, 3414–3425. [Google Scholar] [CrossRef]
- Saia-Cereda, V.M.; Cassoli, J.S.; Martins-de-Souza, D.; Nascimento, J.M. Psychiatric disorders biochemical pathways unraveled by human brain proteomics. Eur. Arch. Psychiatry Clin. Neurosci. 2017, 267, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Willard, S.L.; Sinclair, D.; Mirza, N.; Turetsky, B.; Berretta, S.; Hahn, C. Translational potential of olfactory mucosa for the study of neuropsychiatric illness. Transl. Psychiatry 2015, 5, e527-12. [Google Scholar]
- Mackay-Sim, A. Concise review: Patient-derived olfactory stem cells: New models for brain diseases. Stem Cells 2012, 30, 2361–2365. [Google Scholar] [CrossRef]
- Benítez-King, G.; Riquelme, A.; Ortíz-López, L.; Berlanga, C.; Rodríguez-Verdugo, M.S.; Romo, F.; Calixto, E.; Solís-Chagoyán, H.; Jímenez, M.; Montaño, L.M.; et al. A non-invasive method to isolate the neuronal linage from the nasal epithelium from schizophrenic and bipolar diseases. J. Neurosci. Methods 2011, 201, 35–45. [Google Scholar] [CrossRef]
- Galindo, L.; Moreno, E.; Lopez-Armenta, F.; Guinart, D.; Cuenca-Royo, A.; Izquierdo-Serra, M.; Xicota, L.; Fernandez, C.; Menoyo, E.; Fernandez-Fernandez, J.M.; et al. Cannabis Users Show Enhanced Expression of CB1-5HT2A Receptor Heteromers in Olfactory Neuroepithelium Cells. Mol. Neurobiol. 2018, 55, 6347–6361. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Medina, M.B.; Perea, M.; Torales, J.; Ventriglio, A.; Vitrani, G.; Aguilar, L.; Roncero, C. Cannabis consumption and psychosis or schizophrenia development. Int. J. Soc. Psychiatry 2018, 64, 690–704. [Google Scholar] [CrossRef]
- Di Forti, M.; Quattrone, D.; Freeman, T.P.; Tripoli, G.; Gayer-Anderson, C.; Quigley, H.; Rodriguez, V.; Jongsma, H.E.; Ferraro, L.; La Cascia, C.; et al. The contribution of cannabis use to variation in the incidence of psychotic disorder across Europe (EU-GEI): A multicentre case-control study. Lancet Psychiatry 2019, 6, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Muguruza, C.; Morentin, B.; Meana, J.J.; Alexander, S.P.H.; Callado, L.F. Endocannabinoid system imbalance in the postmortem prefrontal cortex of subjects with schizophrenia. J. Psychopharmacol. 2019, 33, 1132–1140. [Google Scholar] [CrossRef]
- Bioque, M.; García-Bueno, B.; MacDowell, K.S.; Meseguer, A.; Saiz, P.A.; Parellada, M.; Gonzalez-Pinto, A.; Rodriguez-Jimenez, R.; Lobo, A.; Leza, J.C.; et al. Peripheral endocannabinoid system dysregulation in first-episode psychosis. Neuropsychopharmacology 2013, 38, 2568–2577. [Google Scholar] [CrossRef] [PubMed]
- Twitchell, W.; Brown, S.; Mackie, K. Cannabinoids inhibit n- and p/q-type calcium channels in cultured rat hippocampal neurons. J. Neurophysiol. 1997, 78, 43–50. [Google Scholar] [CrossRef]
- Fisyunov, A.; Tsintsadze, V.; Min, R.; Burnashev, N.; Lozovaya, N. Cannabinoids modulate the P-type high-voltage-activated calcium currents in Purkinje neurons. J. Neurophysiol. 2006, 96, 1267–1277. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.L.; Chadwick, B.; Dickstein, D.L.; Purushothaman, I.; Egervari, G.; Rahman, T.; Tessereau, C.; Hof, P.R.; Roussos, P.; Shen, L.; et al. Adolescent exposure to Δ 9 -tetrahydrocannabinol alters the transcriptional trajectory and dendritic architecture of prefrontal pyramidal neurons. Mol. Psychiatry 2019, 24, 588–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarra-Lecue, I.; Mollinedo-Gajate, I.; Meana, J.J.; Callado, L.F.; Diez-Alarcia, R.; Urigüen, L. Chronic cannabis promotes pro-hallucinogenic signaling of 5-HT2A receptors through Akt/mTOR pathway. Neuropsychopharmacology 2018, 43, 2028–2035. [Google Scholar] [CrossRef] [PubMed]
- Jayanthi, S.; Buie, S.; Moore, S.; Herning, R.I.; Better, W.; Wilson, N.M.; Contoreggi, C.; Cadet, J.L. Heavy marijuana users show increased serum apolipoprotein C-III levels: Evidence from proteomic analyses. Mol. Psychiatry 2010, 15, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Saladrigas-Manjón, S.; Dučić, T.; Galindo, L.; Fernández-Avilés, C.; Pérez, V.; de la Torre, R.; Robledo, P. Effects of cannabis use on the protein and lipid profile of olfactory neuroepithelium cells from schizophrenia patients studied by synchrotron-based FTIR spectroscopy. Biomolecules 2020, 10, 329. [Google Scholar] [CrossRef] [Green Version]
- Guinart, D.; Moreno, E.; Galindo, L.; Cuenca-Royo, A.; Barrera-Conde, M.; Pérez, E.J.; Fernández-Avilés, C.; Correll, C.U.; Canela, E.I.; Casadó, V.; et al. Altered Signaling in CB1R-5-HT2AR Heteromers in Olfactory Neuroepithelium Cells of Schizophrenia Patients is Modulated by Cannabis Use. Schizophr. Bull. 2020, 46, 1547–1557. [Google Scholar] [CrossRef]
- Lachén-Montes, M.; Mendizuri, N.; Ausín, K.; Pérez-Mediavilla, A.; Azkargorta, M.; Iloro, I.; Elortza, F.; Kondo, H.; Ohigashi, I.; Ferrer, I.; et al. Smelling the Dark Proteome: Functional Characterization of PITH Domain-Containing Protein 1 (C1orf128) in Olfactory Metabolism. J. Proteome Res. 2020, 19, 4826–4843. [Google Scholar] [CrossRef]
- Pasman, J.A.; Verweij, K.J.H.; Gerring, Z.; Stringer, S.; Sanchez-roige, S.; Treur, J.L.; Abdellaoui, A.; Nivard, M.G.; Baselmans, B.M.L.; Ong, J.; et al. A Causal Influence of Schizophrenia. Nat. Neurosci. 2018, 21, 1161–1170. [Google Scholar] [CrossRef]
- Gillet, L.C.; Navarro, P.; Tate, S.; Röst, H.; Selevsek, N.; Reiter, L.; Bonner, R.; Aebersold, R. Targeted data extraction of the MS/MS spectra generated by data-independent acquisition: A new concept for consistent and accurate proteome analysis. Mol. Cell. Proteom. 2012, 11, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Cook, J.; Gillespie, M.; Haw, R.; et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2020, 48, D498–D503. [Google Scholar] [CrossRef]
- Hulsen, T.; de Vlieg, J.; Alkema, W. BioVenn—A web application for the comparison and visualization of biological lists using area-proportional Venn diagrams. BMC Genom. 2008, 9, 488. [Google Scholar] [CrossRef] [Green Version]
- Geyer, P.E.; Holdt, L.M.; Teupser, D.; Mann, M. Revisiting biomarker discovery by plasma proteomics. Mol. Syst. Biol. 2017, 13, 942. [Google Scholar] [CrossRef]
- Cruz, J.C.; Tseng, H.C.; Goldman, J.A.; Shih, H.; Tsai, L.H. Aberrant Cdk5 activation by p25 triggers pathological events leading to neurodegeneration and neurofibrillary tangles. Neuron 2003, 40, 471–483. [Google Scholar] [CrossRef] [Green Version]
- Nishi, A.; Bibb, J.A.; Matsuyama, S.; Hamada, M.; Higashi, H.; Nairn, A.C.; Greengard, P. Regulation of DARPP-32 dephosphorylation at PKA- and Cdk5-sites by NMDA and AMPA receptors: Distinct roles of calcineurin and protein phosphatase-2A. J. Neurochem. 2002, 832–841. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Blasco, D.; Santofimia-Castanõ, P.; Gonzalez, A.; Almeida, A.; Bolanõs, J.P. Astrocyte NMDA receptors’ activity sustains neuronal survival through a Cdk5-Nrf2 pathway. Cell Death Differ. 2015, 22, 1877–1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Miguel, A.; Javier Meana, J.; García-Sevilla, J.A. Cyclin-dependent kinase-5 and p35/p25 activators in schizophrenia and major depression prefrontal cortex: Basal contents and effects of psychotropic medications. Int. J. Neuropsychopharmacol. 2013, 16, 683–689. [Google Scholar] [CrossRef] [Green Version]
- Bertram, K.; Agafonov, D.E.; Dybkov, O.; Haselbach, D.; Leelaram, M.N.; Will, C.L.; Urlaub, H.; Kastner, B.; Lührmann, R.; Stark, H. Cryo-EM Structure of a Pre-catalytic Human Spliceosome Primed for Activation. Cell 2017, 170, 701–713.e11. [Google Scholar] [CrossRef]
- Reble, E.; Dineen, A.; Barr, C.L. The contribution of alternative splicing to genetic risk for psychiatric disorders. Genes Brain Behav. 2018, 17, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, L.G.; Cooper, S.; Hickey, P.; Rudinger-Thirion, J.; McKenzie, M.; Compton, A.; Lim, S.C.; Thorburn, D.; Ryan, M.T.; Giegé, R.; et al. Mutation of the mitochondrial tyrosyl-tRNA synthetase gene, YARS2, causes myopathy, lactic acidosis, and sideroblastic anemia—MLASA syndrome. Am. J. Hum. Genet. 2010, 87, 52–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezin, G.T.; Amboni, G.; Zugno, A.I.; Quevedo, J.; Streck, E.L. Mitochondrial dysfunction and psychiatric disorders. Neurochem. Res. 2009, 34, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Maurer, I.; Zierz, S.; Möller, H.J. Evidence for a mitochondrial oxidative phosphorylation defect in brains from patients with schizophrenia. Schizophr. Res. 2001, 48, 125–136. [Google Scholar] [CrossRef]
- Gunes, M.; Altindag, A.; Bulut, M.; Demir, S.; Ibiloglu, A.O.; Kaya, M.C.; Atli, A.; Aksoy, N. Oxidative metabolism may be associated with negative symptoms in schizophrenia. Psychiatry Clin. Psychopharmacol. 2017, 27, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Melser, S.; Zottola, A.C.P.; Serrat, R.; Puente, N.; Grandes, P.; Marsicano, G.; Hebert-Chatelain, E. Functional Analysis of Mitochondrial CB1 Cannabinoid Receptors (mtCB1) in the Brain, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; Volume 593. [Google Scholar]
- Bénard, G.; Massa, F.; Puente, N.; Lourenço, J.; Bellocchio, L.; Soria-Gómez, E.; Matias, I.; Delamarre, A.; Metna-Laurent, M.; Cannich, A.; et al. Mitochondrial CB 1 receptors regulate neuronal energy metabolism. Nat. Neurosci. 2012, 15, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Camargo, L.M.; Collura, V.; Rain, J.C.; Mizuguchi, K.; Hermjakob, H.; Kerrien, S.; Bonnert, T.P.; Whiting, P.J.; Brandon, N.J. Disrupted in Schizophrenia 1 interactome: Evidence for the close connectivity of risk genes and a potential synaptic basis for schizophrenia. Mol. Psychiatry 2007, 12, 74–86. [Google Scholar] [CrossRef]
- Shetty, G.A.; Hattiangady, B.; Shetty, A.K. Neural stem cell- and neurogenesis-related gene expression profiles in the young and aged dentate gyrus. Age 2013, 35, 2165–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Forti, M.; Iyegbe, C.; Sallis, H.; Kolliakou, A.; Falcone, M.A.; Paparelli, A.; Sirianni, M.; La Cascia, C.; Stilo, S.A.; Marques, T.R.; et al. Confirmation that the AKT1 (rs2494732) genotype influences the risk of psychosis in cannabis users. Biol. Psychiatry 2012, 72, 811–816. [Google Scholar] [CrossRef]
- Platani, M.; Samejima, I.; Samejima, K.; Kanemaki, M.T.; Earnshaw, W.C. Seh1 targets GATOR2 and Nup153 to mitotic chromosomes. J. Cell Sci. 2018, 131, jcs213140. [Google Scholar] [CrossRef] [Green Version]
- Salas-Cortes, L.; Ye, F.; Tenza, D.; Wilhelm, C.; Theos, A.; Louvard, D.; Raposo, G.; Coudrier, E. Myosin lb modulates the morphology and the protein transport within multi-vesicular sorting endosomes. J. Cell Sci. 2005, 118, 4823–4832. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Xiong, Y.; Montani, J.P.; Yang, Z.; Ming, X.F. Arginase-II activates mTORC1 through myosin-1b in vascular cell senescence and apoptosis. Cell Death Dis. 2018, 9, 313. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Murthy, P.; Bharath, M.M.S. Chemistry, metabolism, and toxicology of cannabis: Clinical implications. Iran. J. Psychiatry 2012, 7, 149–156. [Google Scholar] [PubMed]
- English, J.A.; Fan, Y.; Föcking, M.; Lopez, L.M.; Hryniewiecka, M.; Wynne, K.; Dicker, P.; Matigian, N.; Cagney, G. Reduced protein synthesis in schizophrenia patient-derived olfactory cells. Transl. Psychiatry 2015, 5, e663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarra-Lecue, I.; Diez-Alarcia, R.; Morentin, B.; Meana, J.J.; Callado, L.F.; Urigüen, L. Ribosomal Protein S6 Hypofunction in Postmortem Human Brain Links mTORC1-Dependent Signaling and Schizophrenia. Front. Pharmacol. 2020, 11, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Subjects (C/nc) | Cannabis Users (C/c) | Schizophrenia Non-Cannabis (SCZ/nc) | Schizophrenia Cannabis (SCZ/c) | |
|---|---|---|---|---|
| Age (years) | 31.4 ± 5.5 | 29.2 ± 5.6 | 37 ± 10.1 | 41.4 ± 6.2 |
| Gender (M-F) | 3/2 | 4/1 | 2/3 | 5/0 |
| Tobacco use | ||||
| Users—n (%) | 5 (100%) | 4 (80%) | 4 (80%) | 5 (100%) |
| Units per week µ | 53.8 ± 50.2 | 36.4 ± 40.6 | 91.2 ± 91.0 | 147 ± 62.6 |
| Use duration (years) µ | 11.4 ± 4.9 | 10.4 ± 10.0 | 5.8 ± 6.8 | 24.2 ± 9.6 *,+,# |
| Cannabis use | ||||
| Age first use (years) µ | - | 15.4 ± 2.5 | - | 17.2 ± 3 |
| Units per week µ | - | 16.2 ± 10.2 | - | 19.4 ± 14.6 |
| Use duration (years) µ | - | 12.2 ± 7.2 | - | 17.6 ± 11.5 |
| Antipsychotic treatment | ||||
| Clozapine/Olanzapine | - | - | 1/5 | 3/5 |
| Aripiprazole | - | - | 2/5 | 1/5 |
| Paliperidone | - | - | 1/5 | 1/5 |
| Risperidone | - | - | 1/5 | - |
| Control Subjects (C/nc) | Cannabis Users (C/c) | Schizophrenia Non-Cannabis (SCZ/nc) | Schizophrenia Cannabis (SCZ/c) | |
|---|---|---|---|---|
| Direct series score | 8.8 ± 0.4 | 8.2 ± 1.8 | 7.4 ± 1.5 | 8.8 ± 2.2 |
| Inverse series score µ | 6.3 ± 1.3 | 8.2 ± 5.5 | 4.4 ± 1.5 | 8.8 ± 2.1 |
| Verbal fluency score | 26 ± 1.2 | 22.8 ± 4.5 | 18.8 ± 6.9 | 17.2 ± 5.3 *,+ |
| Direct spatial span | 7.4 ± 0.6 | 5.6 ± 1.9 | 5.2 ± 1.5 | 5.6 ± 1.7 |
| Inverse spatial span | 7.6 ± 2.9 | 5.8 ± 1.5 | 5.6 ± 3 | 5.6 ± 1.7 |
| Emotional recognition | 65.9 ± 8.6 | 73.8 ± 7.4 | 58.2 ± 22.5 | 58.9 ± 10.2 |
| Global assessment of functioning µ | 99 ± 5 | 96 ± 5.5 | 67 ± 8.4 ***,+,++ | 63 ± 2.7 ***,+,++ |
| Neurological soft signs µ | 2.8 ± 1.1 | 5.2 ± 3.1 | 14.8 ± 8.6 **,+ | 13.2 ± 2.3 **,+ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrera-Conde, M.; Ausin, K.; Lachén-Montes, M.; Fernández-Irigoyen, J.; Galindo, L.; Cuenca-Royo, A.; Fernández-Avilés, C.; Pérez, V.; de la Torre, R.; Santamaría, E.; et al. Cannabis Use Induces Distinctive Proteomic Alterations in Olfactory Neuroepithelial Cells of Schizophrenia Patients. J. Pers. Med. 2021, 11, 160. https://doi.org/10.3390/jpm11030160
Barrera-Conde M, Ausin K, Lachén-Montes M, Fernández-Irigoyen J, Galindo L, Cuenca-Royo A, Fernández-Avilés C, Pérez V, de la Torre R, Santamaría E, et al. Cannabis Use Induces Distinctive Proteomic Alterations in Olfactory Neuroepithelial Cells of Schizophrenia Patients. Journal of Personalized Medicine. 2021; 11(3):160. https://doi.org/10.3390/jpm11030160
Chicago/Turabian StyleBarrera-Conde, Marta, Karina Ausin, Mercedes Lachén-Montes, Joaquín Fernández-Irigoyen, Liliana Galindo, Aida Cuenca-Royo, Cristina Fernández-Avilés, Víctor Pérez, Rafael de la Torre, Enrique Santamaría, and et al. 2021. "Cannabis Use Induces Distinctive Proteomic Alterations in Olfactory Neuroepithelial Cells of Schizophrenia Patients" Journal of Personalized Medicine 11, no. 3: 160. https://doi.org/10.3390/jpm11030160
APA StyleBarrera-Conde, M., Ausin, K., Lachén-Montes, M., Fernández-Irigoyen, J., Galindo, L., Cuenca-Royo, A., Fernández-Avilés, C., Pérez, V., de la Torre, R., Santamaría, E., & Robledo, P. (2021). Cannabis Use Induces Distinctive Proteomic Alterations in Olfactory Neuroepithelial Cells of Schizophrenia Patients. Journal of Personalized Medicine, 11(3), 160. https://doi.org/10.3390/jpm11030160