Queen Recognition Signals in Two Primitively Eusocial Halictid Bees: Evolutionary Conservation and Caste-Specific Perception

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bee Collection
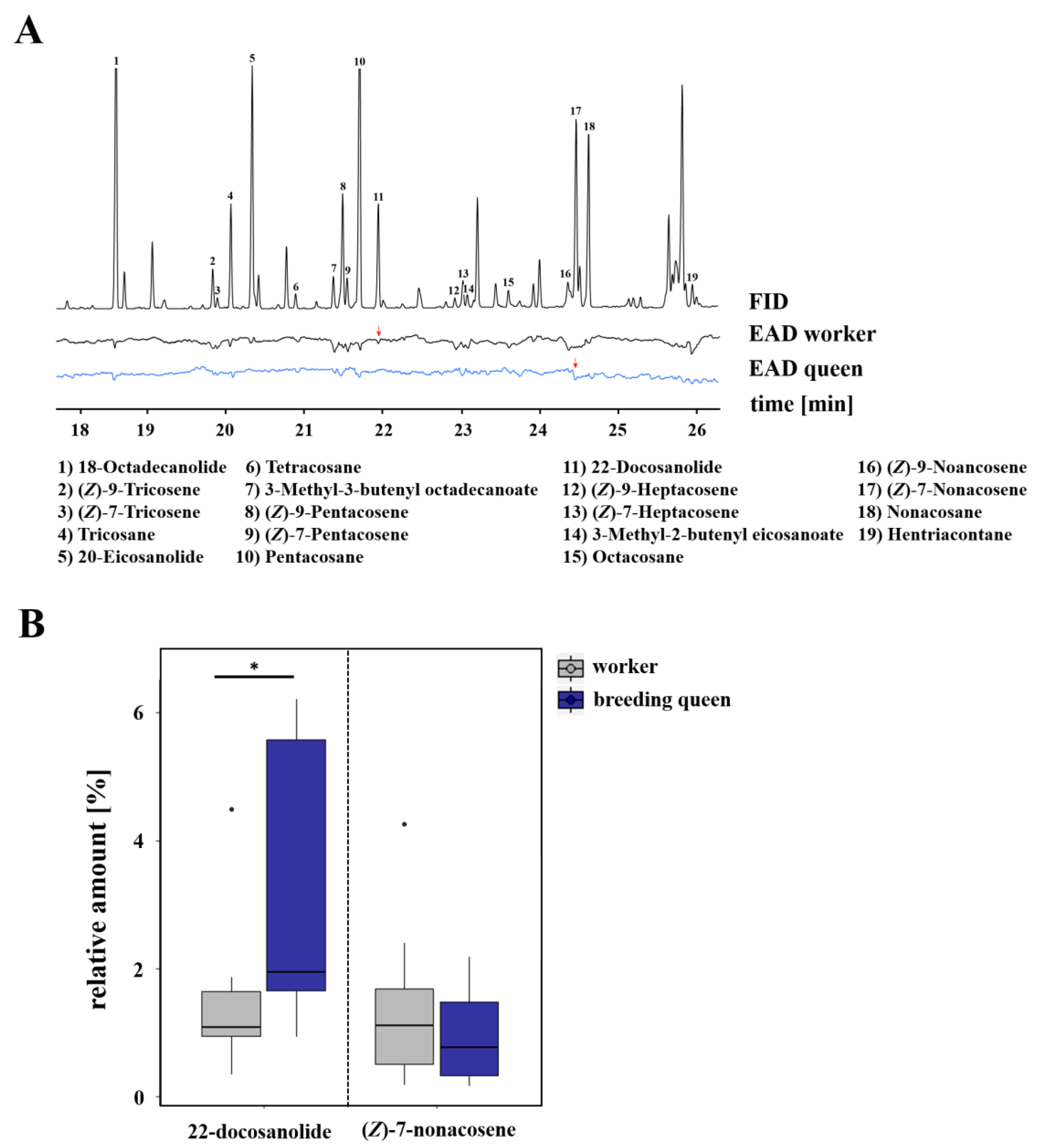
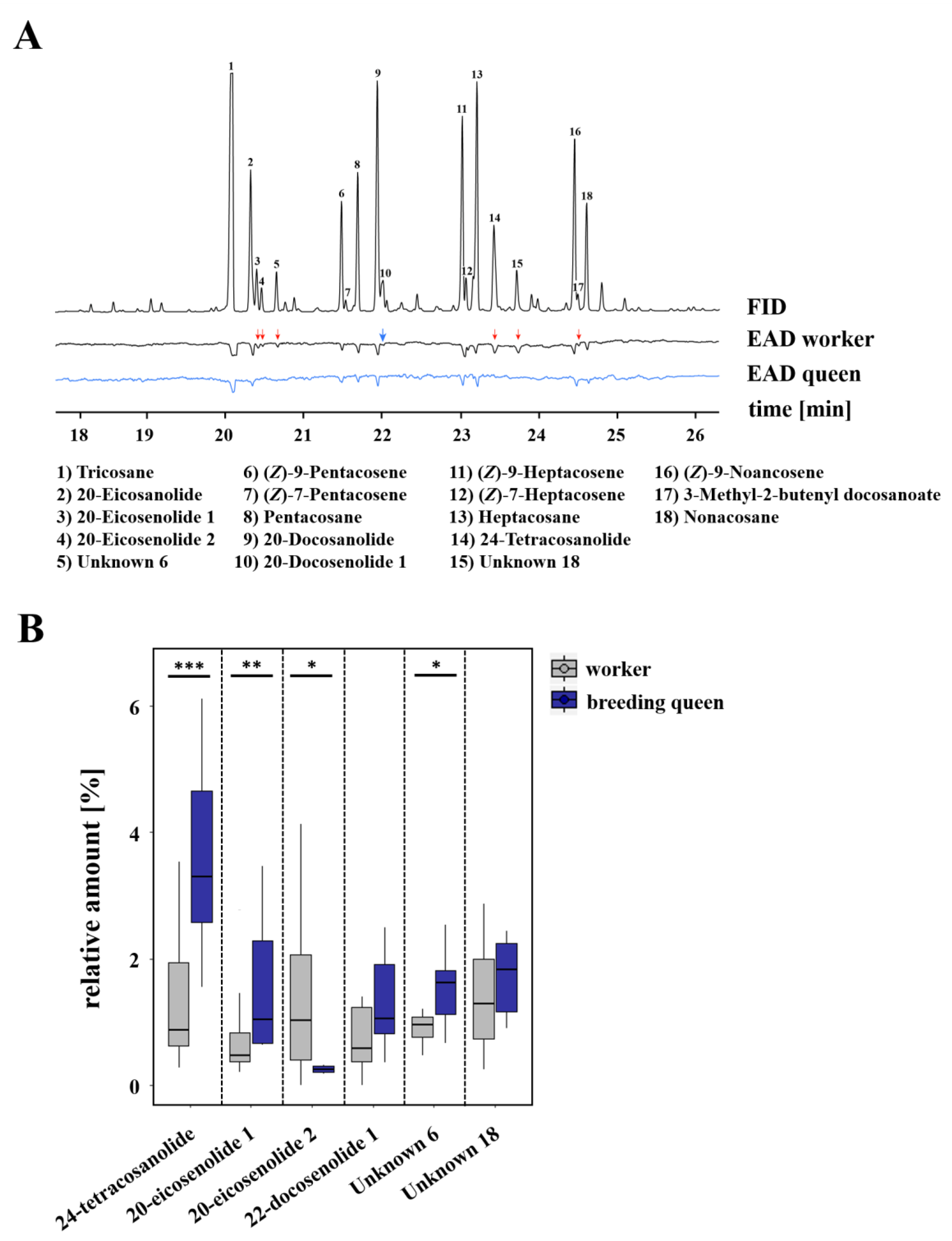
2.2. Chemical Analyses
2.3. Measurement of Physiological State
2.4. Electrophysiological Analyses
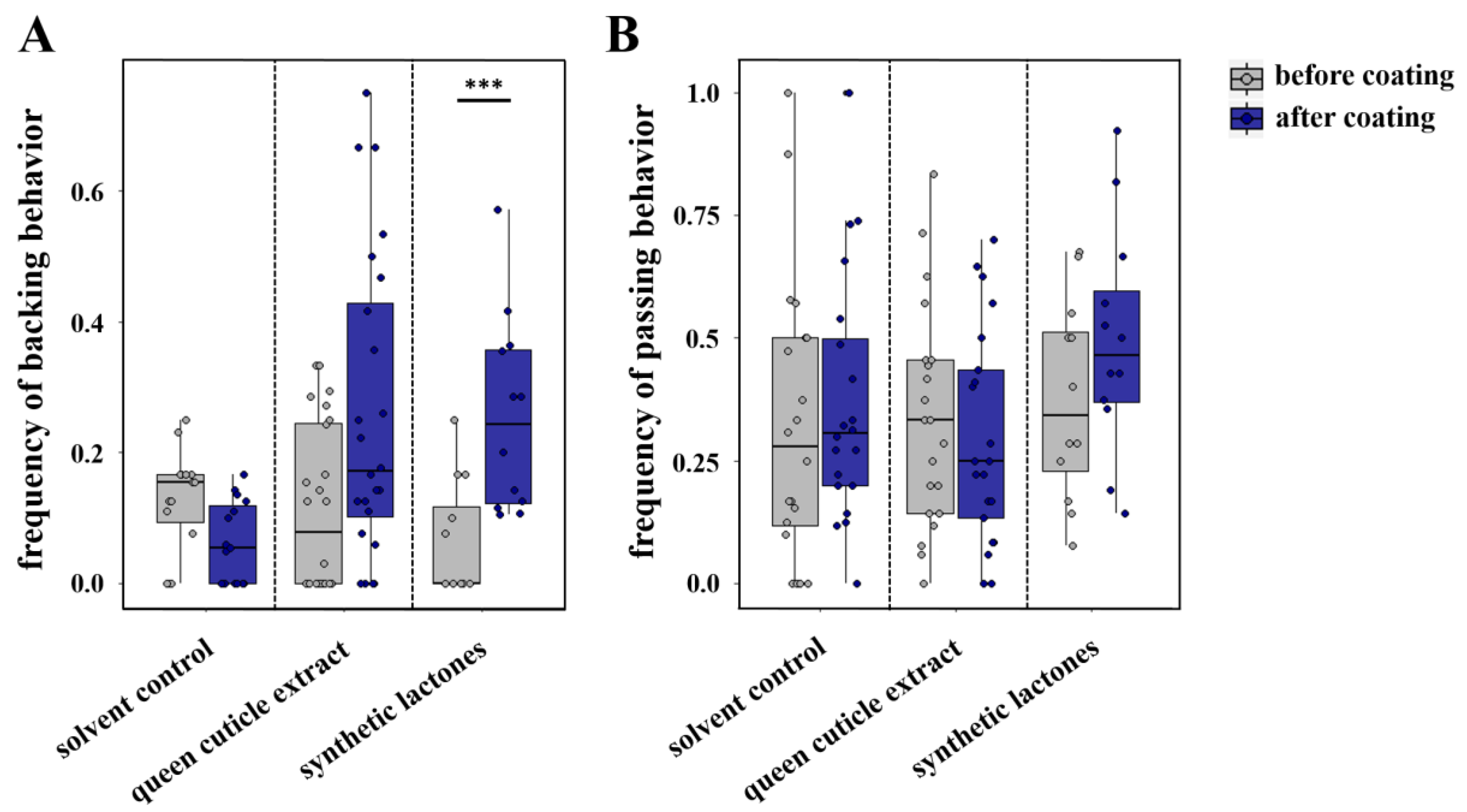
2.5. Circle Tube Bioassays
2.6. Statistics
3. Results
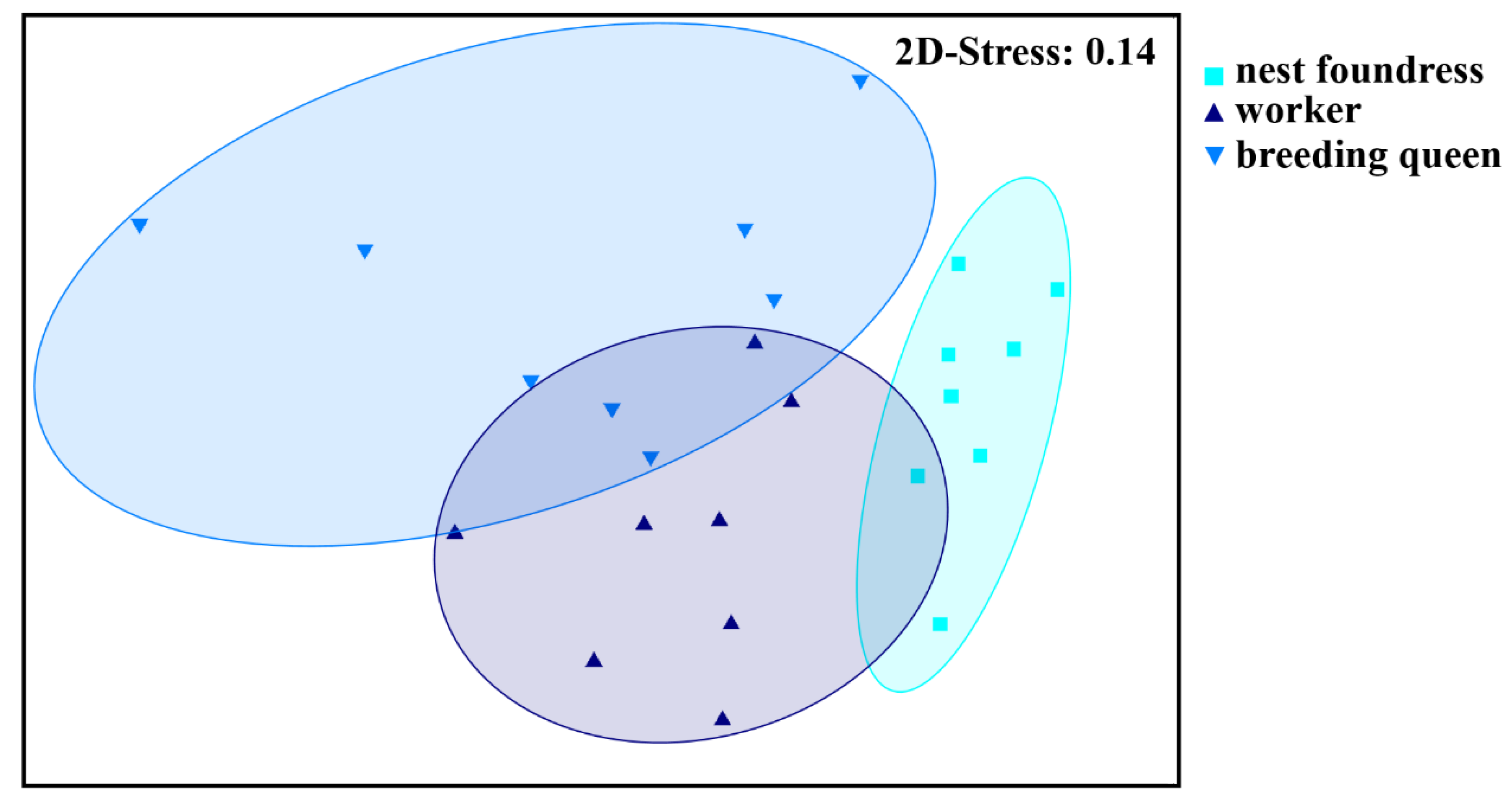
3.1. Caste-Specific and Female-Function-Specific Odor Profiles in L. pauxillum
3.2. Queen–Worker Interactions and Queen Recognition in L. pauxillum
3.3. Caste-Specific Perception of Odor Signals
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilson, E.O. The Insect Societies; Belknap Press of Harvard University Press: Cambridge, MA, USA, 1971. [Google Scholar]
- Ayasse, M.; Jarau, S. Chemical ecology of bumble bees. Annu. Rev. Entomol. 2014, 59, 299–319. [Google Scholar] [CrossRef] [PubMed]
- Kocher, S.D.; Grozinger, C.M. Cooperation, conflict, and the evolution of queen pheromones. J. Chem. Ecol. 2011, 37, 1263–1275. [Google Scholar] [CrossRef] [PubMed]
- Le Conte, Y.; Hefetz, A. Primer pheromones in social Hymenoptera. Annu. Rev. Entomol. 2008, 53, 523–542. [Google Scholar] [CrossRef] [PubMed]
- Monnin, T. Chemical recognition of reproductive status in social insects. Ann. Zool. Fenn. 2006, 43, 515–530. [Google Scholar]
- Oi, C.A.; van Zweden, J.S.; Oliveira, R.C.; Van Oystaeyen, A.; Nascimento, F.S.; Wenseleers, T. The origin and evolution of social insect queen pheromones: Novel hypotheses and outstanding problems. BioEssays 2015, 37, 808–821. [Google Scholar] [CrossRef]
- Rottler-Hoemann, A.M.; Schulz, S.; Ayasse, M. Nest wax triggers worker reproduction in the bumblebee Bombus terrestris. R. Soc. Open Sci. 2016, 3, 150599. [Google Scholar] [CrossRef]
- Holman, L.; Jørgensen, C.G.; Nielsen, J.; d’Ettorre, P. Identification of an ant queen pheromone regulating worker sterility. Proc. R. Soc. Lond. B Biol. Sci. 2010, 277, 3793–3800. [Google Scholar] [CrossRef]
- Holman, L.; Lanfear, R.; d’Ettorre, P. The evolution of queen pheromones in the ant genus Lasius. J. Evol. Biol. 2013, 26, 1549–1558. [Google Scholar] [CrossRef]
- Holman, L.; Hanley, B.; Millar, J.G. Highly specific responses to queen pheromone in three Lasius ant species. Behav. Ecol. Sociobiol. 2016, 70, 387–392. [Google Scholar] [CrossRef]
- Oi, C.A.; Millar, J.G.; van Zweden, J.S.; Wenseleers, T. Conservation of queen pheromones across two species of vespine wasps. J. Chem. Ecol. 2016, 42, 1175–1180. [Google Scholar] [CrossRef]
- Nunes, T.M.; Mateus, S.; Favaris, A.P.; Amaral, M.F.; Von Zuben, L.G.; Clososki, G.C.; Bento, J.M.S.; Oldroyd, B.P.; Silva, R.; Zucchi, R.; et al. Queen signals in a stingless bee: Suppression of worker ovary activation and spatial distribution of active compounds. Sci. Rep. 2014, 4, 7449. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.A.; Hölldobler, B.; Liebig, J. Cuticular hydrocarbons reliably identify cheaters and allow enforcement of altruism in a social insect. Curr. Biol. 2009, 19, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.A.; Millar, J.G.; Hanks, L.M.; Suarez, A.V. Experimental evidence that workers recognize reproductives through cuticular hydrocarbons in the ant Odontomachus brunneus. Behav. Ecol. Sociobiol. 2012, 66, 1267–1276. [Google Scholar] [CrossRef]
- Van Oystaeyen, A.; Oliveira, R.C.; Holman, L.; van Zweden, J.S.; Romero, C.; Oi, C.A.; d’Ettorre, P.; Khalesi, M.; Billen, J.; Wäckers, F.; et al. Conserved class of queen pheromones stops social insect workers from reproducing. Science 2014, 343, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Butler, C.G.; Callow, R.K.; Johnston, N.C. The isolation and synthesis of queen substance, 9-oxodec-trans-2-enoic acid, a honeybee pheromone. Proc. R. Soc. Lond. B Biol. Sci. 1962, 155, 417–432. [Google Scholar]
- Hoover, S.E.R.; Keeling, C.I.; Winston, M.L.; Slessor, K.N. The effect of queen pheromones on worker honey bee ovary development. Naturwissenschaften 2003, 90, 477–480. [Google Scholar] [CrossRef]
- Matsuura, K.; Himuro, C.; Yokoi, T.; Yamamoto, Y.; Vargo, E.L.; Keller, L. Identification of a pheromone regulating caste differentiation in termites. Proc. Natl. Acad. Sci. USA 2010, 107, 12963–12968. [Google Scholar] [CrossRef]
- Smith, A.A.; Millar, J.G.; Suarez, A.V. Comparative analysis of fertility signals and sex-specific cuticular chemical profiles of Odontomachus trap-jaw ants. J. Exp. Biol. 2016, 219, 419–430. [Google Scholar] [CrossRef]
- Steitz, I.; Ayasse, M. Macrocyclic lactones act as a queen pheromone in a primitively eusocial sweat bee. Curr. Biol. under review.
- Steitz, I.; Kingwell, C.; Paxton, R.J.; Ayasse, M. Evolution of caste-specific chemical profiles in halictid bees. J. Chem. Ecol. 2018, 44, 827–837. [Google Scholar] [CrossRef]
- Johansson, I.; Svensson, B.G.; Tengö, J.; Bergström, G. Systematic relationship of halictinae bees based on the pattern of macrocyclic lactones in the Dufour gland secretion. Insect Biochem. 1982, 12, 161–170. [Google Scholar] [CrossRef]
- Cane, J.H. Dufour’s gland secretion in the cell linings of bees (Hymenoptera: Apoidea). J. Chem. Ecol. 1981, 7, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Duffield, R.M.; Fernandes, A.; Lamb, C.; Wheeler, J.W.; Eickwort, G.C. Macrocyclic lactones and isopentenyl esters in the Dufour’s gland secretion of halictine bees (Hymenoptera: Halictidae). J. Chem. Ecol. 1981, 7, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Hefetz, A. The role of Dufour’s gland secretions in bees. Physiol. Entomol. 1987, 12, 243–253. [Google Scholar] [CrossRef]
- Hefetz, A.; Bergström, G.; Tengö, J. Species, individual and kin specific blends in Dufour’s gland secretions of halictine bees. J. Chem. Ecol. 1986, 12, 197–208. [Google Scholar] [CrossRef]
- Smith, B.H.; Carlson, R.G.; Frazier, J. Identification and bioassay of macrocyclic lactone sex pheromone of the halictine bee Lasioglossum zephyrum. J. Chem. Ecol. 1985, 11, 1447–1456. [Google Scholar] [CrossRef]
- Ayasse, M.; Engels, W.; Lübke, G.; Taghizadeh, T.; Francke, W. Mating expenditures reduced via female sex pheromone modulation in the primitively eusocial halictine bee, Lasioglossum (Evylaeus) malachurum (Hymenoptera: Halictidae). Behav. Ecol. Sociobiol. 1999, 45, 95–106. [Google Scholar] [CrossRef]
- Ayasse, M.; Paxton, R.J.; Tengö, J. Mating behavior and chemical communication in the order Hymenoptera. Annu. Rev. Entomol. 2001, 46, 31–78. [Google Scholar] [CrossRef]
- Wcislo, W.T. The role of learning in the mating biology of a sweat bee Lasioglossum zephyrum (Hymenoptera: Halictidae). Behav. Ecol. Sociobiol. 1987, 20, 179–185. [Google Scholar] [CrossRef]
- Smith, B.; Weller, C. Social competition among gynes in halictine bees: The influence of bee size and pheromones on behavior. J. Insect Behav. 1989, 2, 397–411. [Google Scholar] [CrossRef]
- Keller, L.; Nonacs, P. The role of queen pheromones in social insects: Queen control or queen signal? Anim. Behav. 1993, 45, 787–794. [Google Scholar] [CrossRef]
- Babu, M.J.; Ankolekar, S.M.; Rajashekhar, K.P. Castes of the weaver ant Oecophylla smaragdina (Fabricius) differ in the organization of sensilla in their antennae and mouthparts. Curr. Sci. 2011, 101, 755–764. [Google Scholar]
- Fialho, M.C.Q.; Guss-Matiello, C.P.; Zanuncio, J.C.; de Oliveira Campos, L.A.; Serrao, J.E. A comparative study of the antennal sensilla in corbiculate bees. J. Apicult. Res. 2014, 53, 392–403. [Google Scholar] [CrossRef]
- Ravaiano, S.V.; de Paiva Ferreira, R.; de Oliveira Campos, L.A.; Martins, G.F. The antennal sensilla of Melipona quadrifasciata (Hymenoptera: Apidae: Meliponini): A study of different sexes and castes. Naturwissenschaften 2014, 101, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Leonhardt, S.D.; Menzel, F.; Nehring, V.; Schmitt, T. Ecology and evolution of communication in social insects. Cell 2016, 164, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.A.; Liebig, J. The evolution of cuticular fertility signals in eusocial insects. Curr. Opin. Insect Sci. 2017, 22, 79–84. [Google Scholar] [CrossRef]
- Villalta, I.; Abril, S.; Cerdá, X.; Boulay, R. Queen control or queen signal in ants: What remains of the controversy 25 years after Keller and Nonacs’ seminal paper? J. Chem. Ecol. 2018, 44, 805–817. [Google Scholar] [CrossRef]
- Westrich, P. Die Wildbienen Baden-Württembergs; Verlag Eugen Ulmer: Stuttgart, Germany, 1989. [Google Scholar]
- Hölldobler, B.; Michener, C.D. Mechanisms of identification and discrimination in social Hymenoptera. In Evolution of Social Behavior: Hypotheses and Empirical Tests; Markl, H., Ed.; Verlag Chemie: Weinheim, Germany, 1980. [Google Scholar]
- Soro, A.; Ayasse, M.; Zobel, M.U.; Paxton, R.J. Complex sociogenetic organization and the origin of unrelated workers in a eusocial sweat bee, Lasioglossum malachurum. Insect Soc. 2009, 56, 55–63. [Google Scholar] [CrossRef]
- Duchateau, M.J.; Velthuis, H.H.W. Ovarian development and egg laying in workers of Bombus terrestris. Entomol. Exp. Appl. 1989, 51, 199–213. [Google Scholar] [CrossRef]
- Dani, F.R.; Jones, G.R.; Corsi, S.; Beard, R.; Pradella, D.; Turillazzi, S. Nestmate recognition cues in the honey bee: Differential importance of cuticular alkanes and alkenes. Chem. Senses 2005, 30, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Clarke, K.R.; Gorley, R.N. PRIMER V6: User Manual-Tutorial; Plymouth Marine Laboratory: Plymouth, UK, 2006. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Ayasse, M.; Engels, W.; Hefetz, A.; Tengö, J.; Lübke, G.; Francke, W. Ontogenetic patterns of volatiles identified in Dufour’s gland extracts from queens and workers of the primitively eusocial halictine bee, Lasioglossum malachurum (Hymenoptera: Halictidae). Insect Soc. 1993, 40, 41–58. [Google Scholar] [CrossRef]
- Soro, A.; Ayasse, M.; Zobel, M.U.; Paxton, R.J. Kin discriminators in the eusocial sweat bee Lasioglossum malachurum: The reliability of cuticular and Dufour’s gland odours. Behav. Ecol. Sociobiol. 2011, 65, 641–653. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Bagnères, A.G. Insect Hydrocarbons: Biology, Biochemistry and Chemical Ecology; Cambridge University Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Sprenger, P.P.; Burkert, L.H.; Abou, B.; Federle, W.; Menzel, F. Coping with the climate: Cuticular hydrocarbon acclimation of ants under constant and fluctuating conditions. J. Exp. Biol. 2018, 221, jeb171488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, A.G.; Pomonis, J.G. Physical properties of insect cuticular hydrocarbons: The effects of chain length, methyl-branching and unsaturation. Comp. Biochem. Physiol. 1995, 112, 243–249. [Google Scholar] [CrossRef]
- Chapuisat, M. Smells like queen since the cretaceous. Science 2014, 343, 254–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amsalem, E.; Orlova, M.; Grozinger, C.M. A conserved class of queen pheromones? Re-evaluating the evidence in bumblebees (Bombus impatiens). Proc. R. Soc. B Biol. Sci. 2015, 282, 20151800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amsalem, E.; Padilla, M.; Schreiber, P.M.; Altman, N.S.; Hefetz, A.; Grozinger, C.M. Do bumble bee, Bombus impatiens, queens signal their reproductive and mating status to their workers? J. Chem. Ecol. 2017, 43, 563–572. [Google Scholar] [CrossRef]
- Oi, C.A.; Oliveira, R.C.; van Zweden, J.S.; Mateus, S.; Millar, J.G.; Nascimento, F.S.; Wenseleers, T. Do primitively eusocial wasps use queen pheromones to regulate reproduction? A case study of the paper wasp Polistes satan. Front. Ecol. Evol. 2019, 7, 199. [Google Scholar] [CrossRef] [Green Version]
- Padilla, M.; Amsalem, E.; Altman, N.; Hefetz, A.; Grozinger, C.M. Chemical communication is not sufficient to explain reproductive inhibition in the bumblebee Bombus impatiens. R. Soc. Open Sci. 2016, 3, 160576. [Google Scholar] [CrossRef]
- Pardi, L. Dominance order in Polistes wasps. Physiol. Zool. 1948, 21, 1–13. [Google Scholar] [CrossRef]
- Starkey, J.; Brown, A.; Amsalem, E. The road to sociality: Brood regulation of worker reproduction in the simple eusocial bee Bombus impatiens. Anim. Behav. 2019, 154, 57–65. [Google Scholar] [CrossRef]
- Buckle, G.R. Queen-worker behavior and nestmate interactions in young colonies of Lasioglossum zephyrum. Insect Soc. 1982, 29, 125–137. [Google Scholar] [CrossRef]
- Brothers, D.J.; Michener, C.D. Interactions in colonies of primitively social bees. J. Comp. Physiol. 1974, 90, 129–168. [Google Scholar] [CrossRef]
- Groh, C.; Rössler, W. Caste-specific postembryonic development of primary and secondary olfactory centers in the female honeybee brain. Arthropod Struct. Dev. 2008, 37, 459–468. [Google Scholar] [CrossRef]
- Oliveira, R.C.; Oi, C.A.; do Nascimento, M.M.C.; Vollet-Neto, A.; Alves, D.A.; Campos, M.C.; Nascimento, F.; Wenseleers, T. The origin and evolution of queen and fertility signals in Corbiculate bees. BMC Evol. Biol. 2015, 15, 254. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior | Description | Mode of Interaction |
|---|---|---|
| backing | fast movement backwards apart from the encountering bee | avoidance/submission |
| moving backwards | slow movement backwards apart from the encountering bee | avoidance |
| nudging | fast movement forwards, bee brings its face into contact with face of encountering bee | dominance |
| passing | both encountering bees pass and move on in opposite directions | cooperation |
| withdraw | 180 degree turn away from encountering bee | avoidance |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steitz, I.; Brandt, K.; Biefel, F.; Minat, Ä.; Ayasse, M. Queen Recognition Signals in Two Primitively Eusocial Halictid Bees: Evolutionary Conservation and Caste-Specific Perception. Insects 2019, 10, 416. https://doi.org/10.3390/insects10120416
Steitz I, Brandt K, Biefel F, Minat Ä, Ayasse M. Queen Recognition Signals in Two Primitively Eusocial Halictid Bees: Evolutionary Conservation and Caste-Specific Perception. Insects. 2019; 10(12):416. https://doi.org/10.3390/insects10120416
Chicago/Turabian StyleSteitz, Iris, Katharina Brandt, Felix Biefel, Ädem Minat, and Manfred Ayasse. 2019. "Queen Recognition Signals in Two Primitively Eusocial Halictid Bees: Evolutionary Conservation and Caste-Specific Perception" Insects 10, no. 12: 416. https://doi.org/10.3390/insects10120416
APA StyleSteitz, I., Brandt, K., Biefel, F., Minat, Ä., & Ayasse, M. (2019). Queen Recognition Signals in Two Primitively Eusocial Halictid Bees: Evolutionary Conservation and Caste-Specific Perception. Insects, 10(12), 416. https://doi.org/10.3390/insects10120416