Co-Expression of a Homologous Cytochrome P450 Reductase Is Required for In Vivo Validation of the Tetranychus urticae CYP392A16-Based Abamectin Resistance in Drosophila



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Insects
2.3. Generation of pUAST.CYP392A16
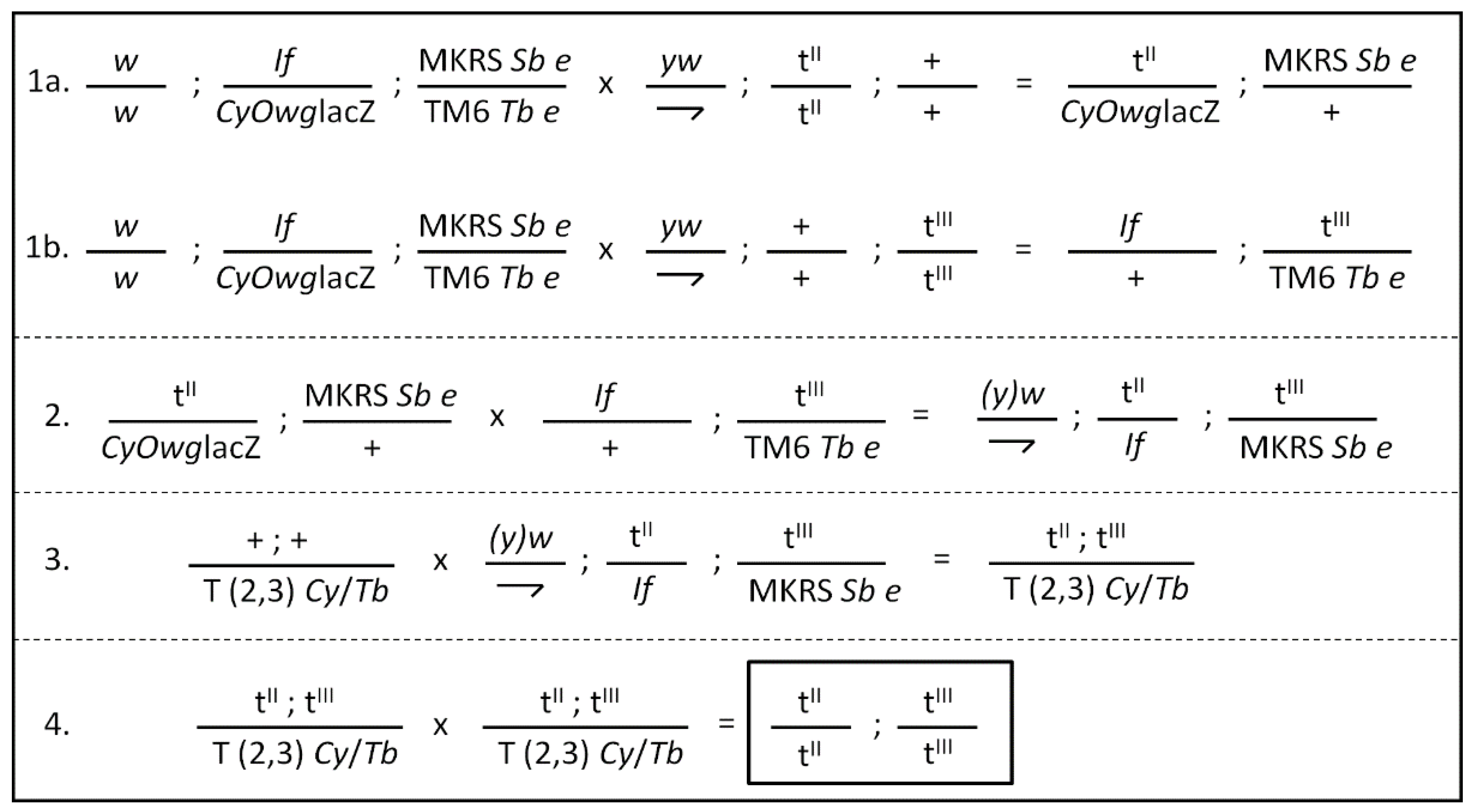
2.4. Construction of the Transgenic Fly Strains
2.5. Expression of CYP392A16 and/or TuCPR in D. Melanogaster
2.6. Extraction of RNA, cDNA Synthesis and Reverse Transcription PCR
2.7. Toxicity Bioassays
3. Results
3.1. Generation of Transgenic Lines Bearing UAS-CYP392A16 and UAS-TuCPR
3.2. GAL4/UAS Transgenic Co-Expression of CYP392A16 and TuCPR Confers Resistance to Abamectin
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Migeon, A.; Nouguier, E.; Dorkeld, F. Spider mites web: A comprehensive database for the Tetranychidae. In Trends in Acarology; Sabelis, M., Bruin, J., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 557–560. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Dermauw, W.; Grbić, M.; Tirry, L.; Feyereisen, R. Spider mite control and resistance management: Does a genome help? Pest Manag. Sci. 2013, 69, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, T.; Tirry, L.; Yamamoto, A.; Nauen, R.; Dermauw, W. The economic importance of acaricides in the control of phytophagous mites and an update on recent acaricide mode of action research. Pestic. Biochem. Physiol. 2015, 121, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Tirry, L. Mechanisms of acaricide resistance in the two-spotted spider mite Tetranychus urticae. In Biorational Control of Arthropod Pests; Ishaaya, I., Horowitz, A.R., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 347–393. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Dermauw, W. The molecular evolution of xenobiotic metabolism and resistance in chelicerate mites. Annu. Rev. Entomol. 2016, 61, 475–498. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Vanholme, B.; Van Pottelberge, S.; Van Nieuwenhuyse, P.; Nauen, R.; Tirry, L.; Denholm, I. Mitochondrial heteroplasmy and the evolution of insecticide resistance: Non-Mendelian inheritance in action. Proc. Nat. Acad. Sci. USA 2008, 105, 5980–5985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsagkarakou, A.; Van Leeuwen, T.; Khajehali, J.; Ilias, A.; Grispou, M.; Williamson, M.S.; Tirry, L.; Vontas, J. Identifications of pyrethroid resistance associated mutations in the para sodium channel of the two-spotted spider mite Tetranychus urticae (Acari: Tetranychidae). Insect Mol. Biol. 2009, 18, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Khajehali, J.; Van Leeuwen, T.; Grispoy, M.; Morou, E.; Alout, H.; Weill, M.; Tirry, L.; Vontas, J.; Tsagkarakou, A. Acetylcholinesterase point mutations in European strains of Tetranychus urticae (Acari: Tetranychidae) resistant to organophosphates. Pest Manag. Sci. 2010, 66, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.H.; Yoon, K.S.; Clark, J.M.; Lee, S.H. A point mutation in a glutamate-gated chloride channel confers abamectin resistance in the two-spotted spider mite, Tetranychus urticae Koch. Insect Mol. Biol. 2010, 19, 583–591. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Demaeght, P.; Osborne, E.J.; Dermauw, W.; Gohlke, S.; Nauen, R.; Grbić, M.; Tirry, L.; Merzendorfer, H.; Clark, R.M. Population bulk segregant mapping uncovers resistance mutations and the mode of action of a chitin synthesis inhibitor in arthropods. Proc. Nat. Acad. Sci. USA 2012, 109, 4407–4412. [Google Scholar] [CrossRef] [Green Version]
- Dermauw, W.; Ilias, A.; Riga, M.; Tsagkarakou, A.; Grbić, M.; Tirry, L.; Van Leeuwen, T.; Vontas, J. The cys-loop ligand-gated ion channel gene family of Tetranychus urticae: Implications for acaricide toxicology and a novel mutation associated with abamectin resistance. Insect Biochem. Mol. Biol. 2012, 42, 455–465. [Google Scholar] [CrossRef]
- Ilias, A.; Vontas, J.; Tsagkarakou, A. Global distribution and origin of target site insecticide resistance mutations in Tetranychus urticae. Insect Biochem. Mol. Biol. 2014, 48, 17–28. [Google Scholar] [CrossRef]
- Bajda, S.; Dermauw, W.; Panteleri, R.; Sugimoto, N.; Douris, V.; Tirry, L.; Osakabe, M.; Vontas, J.; Van Leeuwen, T. A mutation in the PSST homologue of complex I (NADH: Ubiquinone oxidoreductase) from Tetranychus urticae is associated with resistance to METI acaricides. Insect Biochem. Mol. Biol. 2017, 80, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Riga, M.; Bajda, S.; Themistokleous, C.; Papadaki, S.; Palzewicz, M.; Dermauw, W.; Vontas, J.; Van Leeuwen, T. The relative contribution of target-site mutations in complex acaricide resistant phenotypes as assessed by marker assisted backcrossing in Tetranychus urticae. Sci. Rep. 2017, 7, 9202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Adenasaya, A.W.; Morales, M.A.; Walsh, D.B.; Lavine, L.C.; Lavine, M.D.; Zhu, F. Multiple acaricide resistance and underlying mechanisms in Tetranychus urticae on hops. J. Pest Sci. 2019, 92, 543–555. [Google Scholar] [CrossRef]
- Fotoukkiaii, S.M.; Tan, Z.; Xue, W.; Wybouw, N.; Van Leeuwen, T. Identification and characterization of new mutations in mitochondrial cytochrome b that confer resistance to bifenazate and acequinocyl in the spider mite Tetranychus urticae. Pest Manag. Sci. 2020, 76, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Snoeck, S.; Njiru, C.; Inak, E.; Dermauw, W.; Van Leeuwen, T. Geographical distribution and molecular insights into abamectin and milbemectin cross-resistance in European field populations of Tetranychus urticae. Pest Manag. Sci. 2020, 76, 2569–2581. [Google Scholar] [CrossRef]
- Sugimoto, N.; Takahashi, A.; Ihara, R.; Itoh, Y.; Jouraku, A.; Van Leeuwen, T.; Osakabe, M. QTL mapping using microsatellite linkage reveals target-site mutations associated with high levels of resistance against three mitochondrial complex II inhibitors in Tetranychus urticae. Insect Biochem. Mol. Biol. 2020, 123, 103410. [Google Scholar] [CrossRef]
- Papapostolou, K.M.; Riga, M.; Charamis, J.; Skoufa, E.; Souchlas, V.; Ilias, A.; Dermauw, D.; Ioannidis, P.; Van Leeuwen, T.; Vontas, J. Identification and characterization of striking multiple-insecticide resistance in a Tetranychus urticae field population from Greece. Pest Manag. Sci. 2020, in press. [Google Scholar] [CrossRef]
- Dermauw, W.; Wybouw, N.; Rombauts, S.; Menten, B.; Vontas, J.; Grbić, M.; Clark, R.M.; Feyereisen, R.; Van Leeuwen, T. A link between host plant adaptation and pesticide resistance in the polyphagous spider mite Tetranychus urticae. Proc. Nat. Acad. Sci. USA 2013, 110, E113–E122. [Google Scholar] [CrossRef] [Green Version]
- Demaeght, P.; Dermauw, W.; Tsakireli, D.; Khajehali, J.; Nauen, R.; Tirry, L.; Vontas, J.; Lümmen, P.; Van Leeuwen, T. Molecular analysis of resistance to acaricidal spirocyclic tetronic acids in Tetranychus urticae: CYP392E10 metabolizes spirodiclofen, but not its corresponding enol. Insect Biochem. Mol. Biol. 2013, 43, 544–554. [Google Scholar] [CrossRef]
- Riga, M.; Tsakireli, D.; Ilias, A.; Morou, E.; Myridakis, A.; Stephanou, E.G.; Nauen, R.; Dermauw, W.; Van Leeuwen, T.; Paine, M.; et al. Abamectin is metabolized by CYP392A16, a cytochrome P450 associated with high levels of acaricide resistance in Tetranychus urticae. Insect Biochem. Mol. Biol. 2014, 46, 43–53. [Google Scholar] [CrossRef]
- Riga, M.; Myridakis, A.; Tsakireli, D.; Morou, E.; Stephanou, E.G.; Nauen, R.; Van Leeuwen, T.; Douris, V.; Vontas, J. Functional characterization of the Tetranychus urticae CYP392A11, a cytochrome P450 that hydroxylates the METI acaricides cyenopyrafen and fenpyroximate. Insect Biochem. Mol. Biol. 2015, 65, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Fotoukkiaii, S.M.; Mermans, C.; Wybouw, N.; Van Leeuwen, T. Resistance risk assessment of the novel complex II inhibitor pyflubumide in the polyphagous pest Tetranychus urticae. J. Pest Sci. 2020, 93, 1085–1096. [Google Scholar] [CrossRef] [Green Version]
- Pavlidi, N.; Mousaalreza, K.; Myridakis, A.; Dermauw, W.; Wybouw, N.; Tsakireli, D.; Stephanou, E.G.; Labrou, N.E.; Vontas, J.; Van Leeuwen, T. A glutathione-S-transferase (TuGSTd05) associated with acaricide resistance in Tetranychus urticae directly metabolizes the complex II inhibitor cyflumetofen. Insect Biochem. Mol. Biol. 2017, 80, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Koo, H.N.; Kim, S.I.; Park, B.; Kim, H.; Kim, G.H. Target-site mutations and glutathione S-transferases are associated with acequinocyl and pyridaben resistance in the two-spotted spider mite Tetranychus urticae (Acari: Tetranychidae). Insects 2020, 11, 511. [Google Scholar] [CrossRef]
- Wei, P.; Demaeght, P.; De Schutter, K.; Grigoraki, L.; Labropoulou, V.; Riga, M.; Vontas, J.; Nauen, R.; Dermauw, W.; Van Leeuwen, T. Overexpression of an alternative allele of carboxyl/choline esterase 4 (CCE04) of Tetranychus urticae is associated with high levels of resistance to the keto-enol acaricide spirodiclofen. Pest Manag. Sci. 2020, 76, 1142–1153. [Google Scholar] [CrossRef]
- Dermauw, W.; Van Leeuwen, T. The ABC gene family in arthropods: Comparative genomics and role in insecticide transport and resistance. Insect Biochem. Mol. Biol. 2014, 45, 89–110. [Google Scholar] [CrossRef]
- Snoeck, S.; Pavlidi, N.; Pipini, D.; Vontas, J.; Dermauw, W.; Van Leeuwen, T. Substrate specificity and promiscuity of horizontally transferred UDP-glucosyltransferases in the generalist herbivore Tetranychus urticae. Insect Biochem. Mol. Biol. 2019, 109, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.M.; Scott, J.G.; Campos, F.; Bloomquist, J.R. Resistance to avermectins: Extent, mechanisms, and management implications. Annu. Rev. Entomol. 1994, 40, 1–30. [Google Scholar] [CrossRef]
- Wolstenholme, A.J. Glutamate-gated chloride channels. J. Biol. Chem. 2012, 287, 40232–40238. [Google Scholar] [CrossRef] [Green Version]
- Bajda, S.; Riga, M.; Wybouw, N.; Papadaki, S.; Ouranou, E.; Fotoukkiaii, S.M.; Vontas, J.; Van Leeuwen, T. Fitness costs of key point mutations that underlie acaricide target-site resistance in the two-spotted spider mite Tetranychus urticae. Evol. Appl. 2018, 11, 1540–1553. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Y.; Zhang, Y.; Wu, Q.; Guo, Z.; Xie, W.; Zhou, X.; Wang, S. Transcriptome profiling and functional analysis suggest that the constitutive overexpression of four cytochrome P450s confers resistance to abamectin in Tetranychus urticae from China. Pest Manag. Sci. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Insect CYP genes and P450 Enzymes. In Insect Molecular Biology and Biochemistry; Gilbert, L.I., Ed.; Academic Press: London, UK, 2012; pp. 236–316. [Google Scholar]
- Grbić, M.; Van Leeuwen, T.; Clark, R.M.; Rombauts, S.; Rouzé, P.; Grbić, V.; Osborne, E.J.; Dermauw, W.; Ngoc, P.C.T.; Ortego, F.; et al. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature 2011, 479, 487–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snoeck, S.; Kurlovs, A.H.; Bajda, S.; Feyereisen, R.; Greenhalgh, R.; Villacis-Perez, E.; Kosterlitz, O.; Dermauw, W.; Clark, R.M.; Van Leeuwen, T. High-resolution QTL mapping in Tetranychus urticae reveals acaricide-specific responses and common target-site resistance after selection by different METI-I acaricides. Insect Biochem. Mol. Biol. 2019, 110, 19–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wybouw, N.; Kosterlitz, O.; Kurlovs, A.H.; Bajda, S.; Greenhalgh, R.; Snoeck, S.; Bui, H.; Bryon, A.; Dermauw, W.; Van Leeuwen, T.; et al. Long-term population studies uncover the genome structure and genetic basis of xenobiotic and host plant adaptation in the herbivore Tetranychus urticae. Genetics 2019, 211, 1409–1427. [Google Scholar] [CrossRef] [Green Version]
- Adesanya, A.W.; Cardenas, A.; Lavine, M.D.; Walsh, D.B.; Lavine, L.C.; Zhu, F. RNA interference of NADPH-cytochrome P450 reductase increases susceptibilities to multiple acaricides in Tetranychus urticae. Pestic. Biochem. Physiol. 2020, 165, 104550. [Google Scholar] [CrossRef]
- Homem, R.A.; Davies, T.G.E. An overview of functional genomic tools in deciphering insect resistance. Curr. Opin. Insect Sci. 2018, 27, 103–110. [Google Scholar] [CrossRef]
- Perry, T.; Batterham, P. Harnessing model organisms to study insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 61–67. [Google Scholar] [CrossRef]
- Scott, J.G.; Buchon, N. Drosophila melanogaster as a powerful tool for studying insect toxicology. Pestic. Biochem. Physiol. 2019, 161, 95–103. [Google Scholar] [CrossRef]
- Douris, V.; Denecke, S.; Van Leeuwen, T.; Nauen, R.; Bass, C.; Vontas, J. Using CRISPR/Cas9 genome modification to understand the genetic basis of insecticide resistance: Drosophila and beyond. Pestic. Biochem. Physiol. 2020, 167, 104595. [Google Scholar] [CrossRef]
- McLeman, A.; Troczka, B.J.; Homem, R.A.; Duarte, A.; Zimmer, C.; Garrood, W.T.; Pym, A.; Beadle, K.; Reid, R.J.; Douris, V.; et al. Fly-Tox: A panel of transgenic flies expressing pest and pollinator cytochrome P450s. Pestic. Biochem. Physiol. 2020, 169, 104674. [Google Scholar] [CrossRef]
- Samantsidis, G.R.; Panteleri, R.; Denecke, S.; Kounadi, S.; Christou, I.; Nauen, R.; Douris, V.; Vontas, J. ‘What I cannot create, I do not understand’: Functionally validated synergism of metabolic and target site insecticide resistance. Proc. R. Soc. B 2020, 287, 20200838. [Google Scholar] [CrossRef] [PubMed]
- Daborn, P.J.; Lumb, C.; Boey, A.; Wong, W.; ffrench-Constant, R.H.; Batterham, P. Evaluating the insecticide resistance potential of eight Drosophila melanogaster cytochrome P450 genes by transgenic over-expression. Insect Biochem. Mol. Biol. 2007, 37, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Bloomington Drosophila Stock Center, Indiana University Bloomington. Available online: https://bdsc.indiana.edu/information/recipes/uclfood.html (accessed on 22 November 2020).
- Pavlidi, N.; Monastirioti, M.; Daborn, P.; Livadaras, I.; Van Leeuwen, T.; Vontas, J. Transgenic expression of the Aedes aegypti CYP9J28 confers pyrethroid resistance in Drosophila melanogaster. Pestic. Biochem. Physiol. 2012, 104, 132–135. [Google Scholar] [CrossRef]
- Chung, H.; Bogwitz, M.R.; McCart, C.; Andrianopoulos, A.; ffrench-Constant, R.H.; Batterham, P.; Daborn, P.J. Cis-regulatory elements in the Accord retrotransposon result in tissue-specific expression of the Drosophila melanogaster insecticide resistance gene Cyp6g1. Genetics 2007, 175, 1071–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, J.L.; Russel, R.M.; Preisler, H.K.; Savin, N.E. Bioassays in Arthropods, 2nd ed.; CRC Press, Taylor and Francis Group LLC: Boca Raton, FL, USA, 2007. [Google Scholar]
- Karunker, I.; Morou, E.; Nikou, D.; Nauen, R.; Sertchook, R.; Stevenson, B.J.; Paine, M.J.I.; Morin, S.; Vontas, J. Structural model and functional characterization of the Bemisia tabaci CYP6CM1vQ, a cytochrome P450 associated with high levels of imidacloprid resistance. Insect Biochem. Mol. Biol. 2009, 39, 697–706. [Google Scholar] [CrossRef]
- Zimmer, C.T.; Garrood, W.T.; Singh, K.S.; Randall, E.; Lueke, B.; Gutbrod, O.; Matthiesen, S.; Kohler, M.; Nauen, R.; Davies, T.G.E. Neofunctionalization of duplicated P450 genes drives the evolution of insecticide resistance in the brown planthopper. Curr. Biol. 2018, 28, 268–274. [Google Scholar] [CrossRef] [Green Version]
- Manjon, C.; Troczka, B.J.; Zaworra, M.; Beadle, K.; Randall, E.; Hertlein, G.; Singh, K.S.; Zimmer, C.T.; Homem, R.A.; Lueke, B.; et al. Unravelling the molecular determinants of bee sensitivity to neonicotinoid insecticides. Curr. Biol. 2018, 28, 1137–1143. [Google Scholar] [CrossRef] [Green Version]
- Troczka, B.J.; Homem, R.A.; Reid, R.; Beadle, K.; Kohler, M.; Zaworra, M.; Field, L.M.; Williamson, M.S.; Nauen, R.; Bass, C.; et al. Identification and functional characterisation of a novel N-cyanoamidine neonicotinoid metabolising cytochrome P450, CYP9Q6, from the buff-tailed bumblebee Bombus terrestris. Insect Biochem. Mol. Biol. 2019, 111, 103171. [Google Scholar] [CrossRef]
- Daborn, P.J.; Lumb, C.; Harrop, T.W.R.; Blasetti, A.; Pasricha, S.; Morin, S.; Michell, S.N.; Donnelly, M.J.; Muller, P.; Batterham, P. Using Drosophila melanogaster to validate metabolism-based insecticide resistance from insect pests. Insect Biochem. Mol. Biol. 2012, 42, 918–924. [Google Scholar] [CrossRef]
- Riveron, J.M.; Irving, H.; Ndula, M.; Barnes, K.G.; Ibrahim, S.; Cuamba, N.; Paine, M.J.I.; Wondji, C. Directionally selected P450 allelles are driving the spread of pyrethroid resistance in the major malaria vector Anopheles funestus. Proc. Natl. Acad. Sci. USA 2013, 110, 252–257. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, S.S.; Riveron, J.M.; Bibby, J.; Irving, H.; Yunta, C.; Paine, M.J.I.; Wondji, C.S. Allelic variation of cytochrome P450s drives resistance to bednet insecticides in a major malaria vector. PLoS Genet. 2015, 11, e1005618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishak, I.H.; Riveron, J.M.; Ibrahim, S.S.; Stott, R.; Longbottom, J.; Irving, H.; Wondji, C.S. The cytochrome P450 gene CYP6P12 confers pyrethroid resistance in kdr-free Malaysian populations of the dengue vector Aedes albopictus. Sci. Rep. 2016, 6, 24707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsakireli, D.; Riga, M.; Kounadi, S.; Douris, V.; Vontas, J. Functional characterization of CYP6A51, a cytochrome P450 associated with pyrethroid resistance in the Mediterranean fruit fly Ceratitis capitata. Pestic. Biochem. Physiol. 2019, 157, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Vassiliou, V.A.; Kitsis, P. Acaricide resistance in Tetranychus urticae (Acari: Tetranychidae) populations from Cyprus. J. Econ. Entomol. 2013, 106, 1848–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, C.B.S.; Andrade, F.H.N.; Rodrigues, A.R.S.; Siqueira, H.A.A. Resistance in field populations of Tetranychus urticae to acaricides and characterization of the inheritance of abamectin resistance. Crop Prot. 2015, 67, 77–83. [Google Scholar] [CrossRef]
- Çağatay, N.S.; Menault, P.; Riga, M.; Vontas, J.; Ay, R. Identification and characterization of abamectin resistance in Tetranychus urticae Koch populations from greenhouses in Turkey. Crop Prot. 2018, 112, 112–117. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Primer Name | Sequence (5′–3′) | Product Size (bp) | Reference |
|---|---|---|---|
| CYP392A16_F | AAATACCGAGGTCGGACGTA | 117 | [20] |
| CYP392A16_R | AAGCACTTTTTCAATCTGGTCAC | ||
| RPL11_Dm_F | CGATCCCTCCATCGGTATCT | 120 | [45] |
| RPL11_Dm_R | AACCACTTCATGGCATCCTC | ||
| pUASTF | TATGTCACACCACAGAAGTAAG | n/a | [47] |
| pUASTR | CAAGTAAATCAACTGCAACTACTG |
| Cross | Transgene Dosage | LC50 (95% FL) (mg/L) | Slope | χ2 (df) | RR 1 (95% FL) | |||
|---|---|---|---|---|---|---|---|---|
| Female | Male | Gal4 | CPR | A16 | ||||
| yw | HR-GAL4 | 1 | - | - | 45.5 (33.3–56.9) | 1.9 ± 0.35 | 12.1 (16) | |
| UAS-CYP392A16.71 | HR-GAL4 | 1 | - | 1 | 53.4 (38.7–76.2) | 1.316 ± 0.25 | 17.8 (16) | 1.17 (0.81–1.71) |
| UAS-TuCPR92; UAS-CYP392A16.71 | HR-GAL4 | 1 | 1 | 1 | 85.2 (78.6–92.7) | 7.1 ± 1.6 | 10.9 (16) | 1.88 (1.44–2.44) |
| UAS-TuCPR32; UAS-CYP392A16.71 | HR-GAL4 | 1 | 1 | 1 | 101.8 (88.6–127.6) | 3.7 ± 0.62 | 10.2 (15) | 2.24 (1.65–3.04) |
| HR-GAL4; UAS-CYP392A16.71 | HRGAL4; UAS-CYP392A16.71 | 2 | - | 2 | 45.07 (37.85–51.91) | 3.41 ± 0.43 | 7.98 (10) | 0.99 (0.74–1.33) |
| UAS-TuCPR32; UAS-CYP392A16.71 | HR-GAL4; UAS-CYP392A16.71 | 1 | 1 | 2 | 77.01 (65.99–89.73) | 4.37 ± 0.47 | 14.4 (10) | 1.69 (1.29–2.23) |
| UAS-CYP392A16.9 | HR-GAL4 | 1 | - | 1 | 28.3 (10.3–42) | 2.1 ± 0.4 | 40.3 (15) | 0.62 (0.42–0.93) |
| UAS-TuCPR92; UAS-CYP392A16.9 | HR-GAL4 | 1 | 1 | 1 | 54.4 (40.1–64.3) | 4.9 ± 0.8 | 30.3 (16) | 1.19 (0.89–1.59) |
| UAS-TuCPR32; UAS-CYP392A16.9 | HR-GAL4 | 1 | 1 | 1 | 82.7 (61.7–117.5) | 3.1 ± 0.4 | 49.6 (13) | 1.82 (1.37–2.42) |
| HR-GAL4; UAS-CYP392A16.9 | HR-GAL4; UAS-CYP392A16.9 | 2 | - | 2 | 31.06 (18.82–40.36) | 3.2 ± 0.5 | 14.88 (9) | 0.68 (0.79–0.96) |
| UAS-TuCPR32; UAS-CYP392A16.9 | HR-GAL4; UAS-CYP392A16.9 | 1 | 1 | 2 | 105.29 (79.91–135.06) | 4.6 ± 0.6 | 17.4 (8) | 2.32 (1.74–3.08) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riga, M.; Ilias, A.; Vontas, J.; Douris, V. Co-Expression of a Homologous Cytochrome P450 Reductase Is Required for In Vivo Validation of the Tetranychus urticae CYP392A16-Based Abamectin Resistance in Drosophila. Insects 2020, 11, 829. https://doi.org/10.3390/insects11120829
Riga M, Ilias A, Vontas J, Douris V. Co-Expression of a Homologous Cytochrome P450 Reductase Is Required for In Vivo Validation of the Tetranychus urticae CYP392A16-Based Abamectin Resistance in Drosophila. Insects. 2020; 11(12):829. https://doi.org/10.3390/insects11120829
Chicago/Turabian StyleRiga, Maria, Aris Ilias, John Vontas, and Vassilis Douris. 2020. "Co-Expression of a Homologous Cytochrome P450 Reductase Is Required for In Vivo Validation of the Tetranychus urticae CYP392A16-Based Abamectin Resistance in Drosophila" Insects 11, no. 12: 829. https://doi.org/10.3390/insects11120829
APA StyleRiga, M., Ilias, A., Vontas, J., & Douris, V. (2020). Co-Expression of a Homologous Cytochrome P450 Reductase Is Required for In Vivo Validation of the Tetranychus urticae CYP392A16-Based Abamectin Resistance in Drosophila. Insects, 11(12), 829. https://doi.org/10.3390/insects11120829