Compatibility between Entomopathogenic Fungi and Egg Parasitoids (Trichogrammatidae): A Laboratory Study for Their Combined Use to Control Duponchelia fovealis

 ,
, 
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Duponchelia fovealis Rearing
2.2. Trichogramma Rearing
2.3. Entomopathogenic Fungal Strains and Commercial Bioinsecticides
2.4. Trichogramma atopovirilia and Trichogramma pretiosum Parasitism in Free-Choice Test
2.5. Spraying of Entomopathogenic Fungi Pre- and Post-Parasitism (No-Choice Tests)
2.6. Statistical Analysis
3. Results
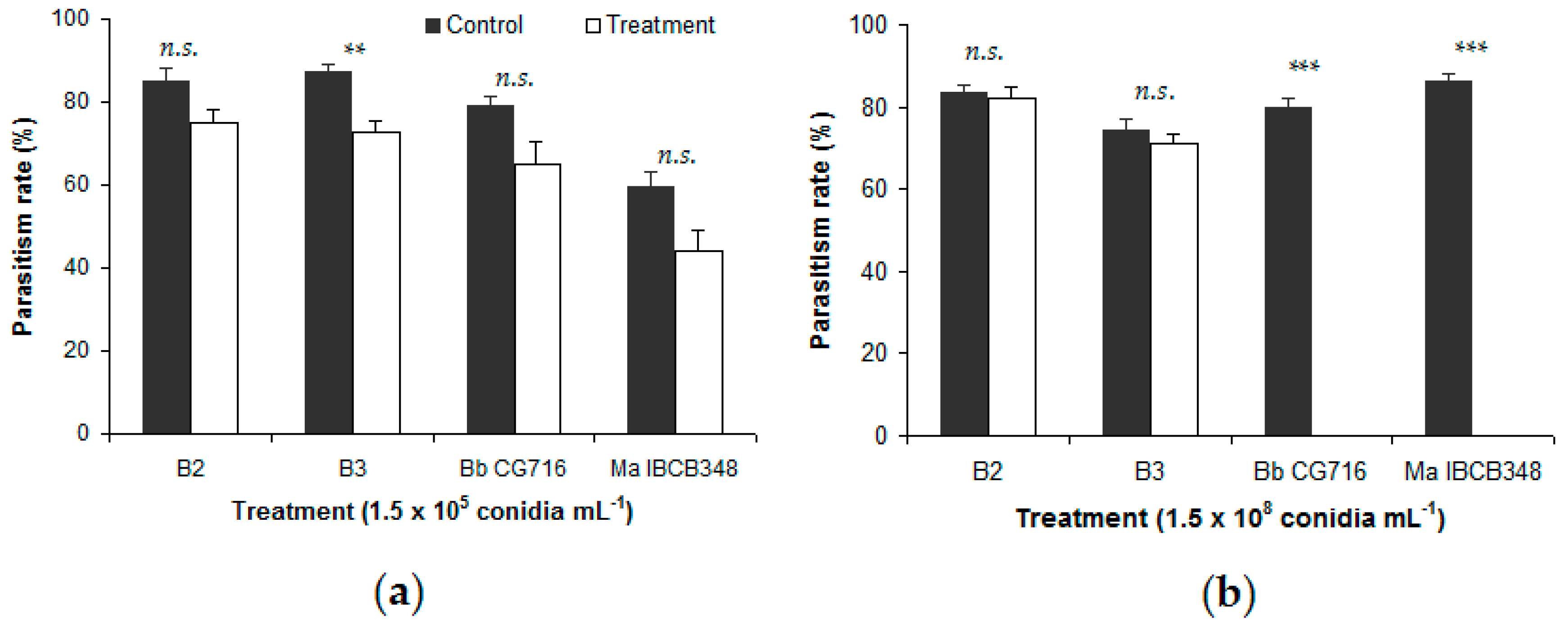
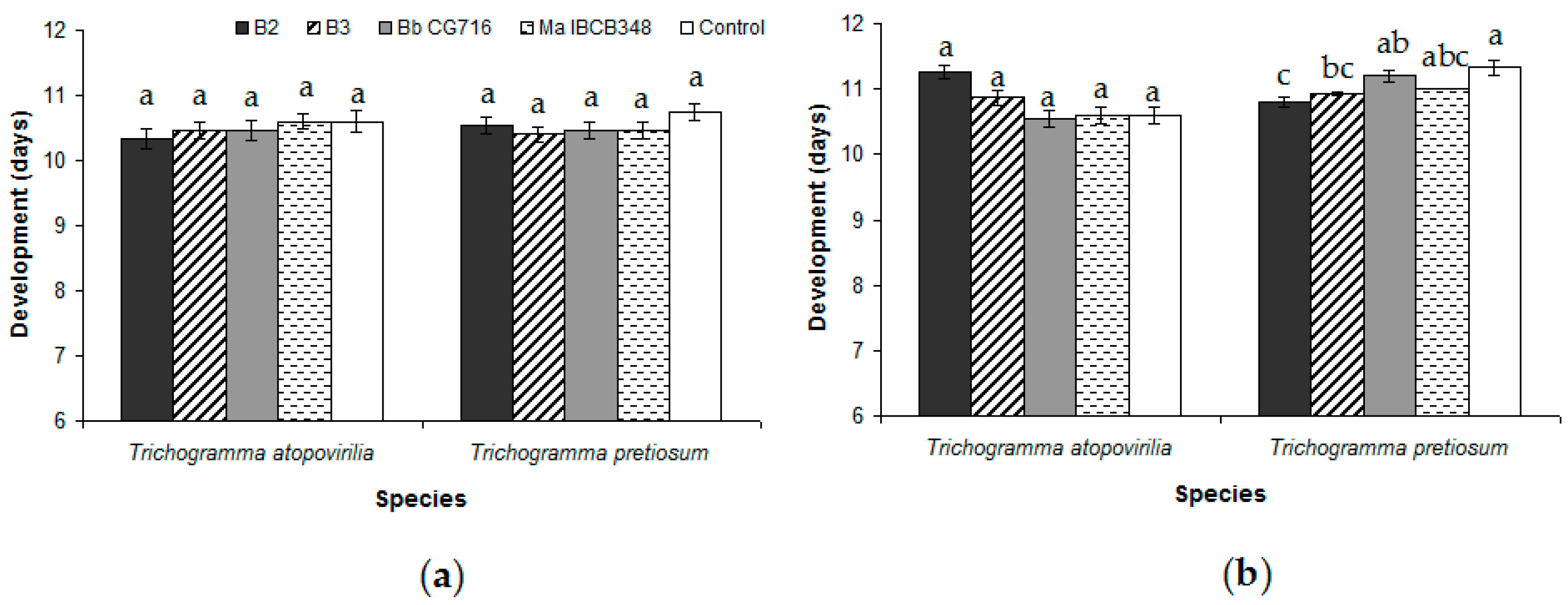
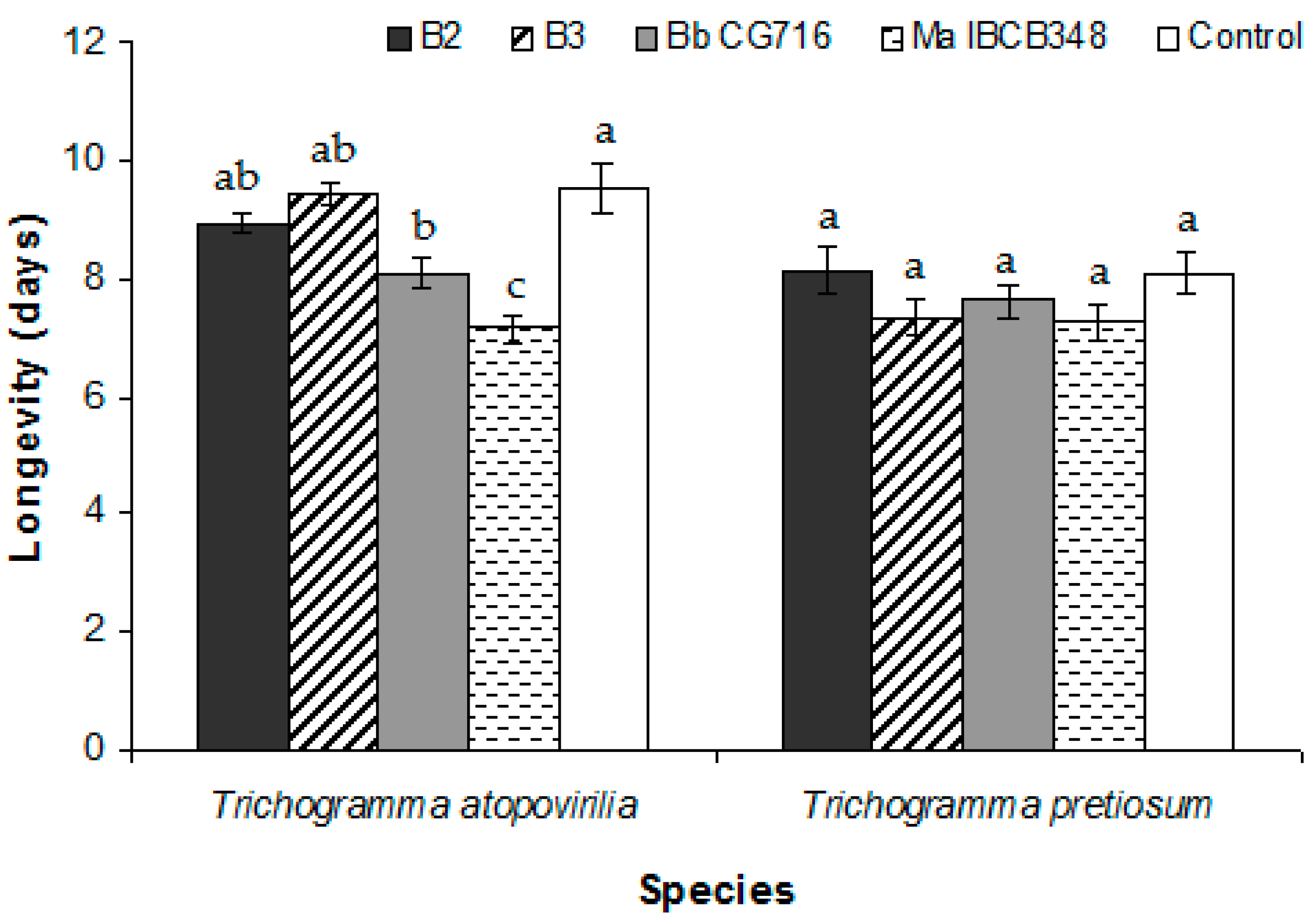
3.1. Trichogramma atopovirilia and Trichogramma pretiosum Parasitism in Free-Choice Test
3.2. Spraying of Entomopathogenic Fungi Pre- and Post-Parasitism (No-Choice Tests)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ponce Ados, R.; Bastiani, M.I.D.; Minim, V.P.; Vanetti, M.C.D. Características físico-químicas e microbiológicas de morango minimamente processado. Ciência Tecnol. Aliment. 2009, 30, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Zawadneak, M.A.C.; Vidal, H.R.; Santos, B. Lagarta-da-coroa, Duponchelia fovealis (Lepidoptera: Crambidae). In Pragas Introduzidas: Insetos e Acaros; Vilela, E., Zucchi, R.A., Eds.; FEALQ: Piracicaba, Brazil, 2015. [Google Scholar]
- Zawadneak, M.A.C.; Gonçalves, R.B.; Pimentel, I.C.; Schuber, J.M.; Santos, B.; Poltronieri, A.S.; Solis, M.A. First record of Duponchelia fovealis (Lepidoptera: Crambidae) in South America. Idesia 2016, 34, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Bonsignore, C.P.; Vacante, V. Duponchelia fovealis (Zeller). A new emergency for strawberry? Protez. Colt. 2010, 3, 40–43. [Google Scholar]
- Franco, M.C.; Baptista, M.C. Duponchelia fovealis Zeller—Nova praga em Portugal. Frutas Legumes Flores 2010, 110, 34–35. [Google Scholar]
- Efil, L.; Özgür, O.; Efil, F. A new pest, Duponchelia fovealis Zeller, on strawberries in Turkey: Damage, distribution and parasitoid. J. Entomol. Zool. Stud. 2014, 2, 328–334. [Google Scholar]
- BRASIL, Ministério da Agricultura, Pecuária e Abastecimento. Agrofit—Sistema de Agrotóxicos Fitossanitários. Available online: http://agrofit.agricultura.gov.br/agrofit_cons/principal_agrofit_cons (accessed on 21 February 2020).
- Jalali, S.K. Natural occurrence, host range and distribution of trichogrammatid egg parasitoids. In Biological Control of Insect Pests Using Egg Parasitoids; Sithanantham, S., Ballal, C., Jalali, S., Bakthavatsalam, N., Eds.; Springer: New Delhi, India, 2013; pp. 67–76. [Google Scholar]
- Paes, J.P.P.; Lima, V.L.S.; Pratissoli, D.; de Carvalho, J.R.; de Freitas Bueno, R.C.O. Selection of parasitoids of the genus Trichogramma (Hymenoptera: Trichogrammatidae) and parasitism at different eggs ages of Duponchelia fovealis (Lepidoptera: Crambidae). Acta Sci. Biol. 2018, 40, e42216. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, R.; Jaras, L.I.; Poltronieri, A.S.; Pimentel, I.C.; Zawadneak, M.A.C. Selectivity of growth regulators and botanical insecticides in the parasitism of three Trichogramma species on eggs of Duponchelia fovealis Zeller (Lepidoptera: Crambidae). EntomoBrasilis 2017, 10, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Feltrin-Campos, E.; Ringenberg, R.; Carvalho, G.A.; Glaeser, D.F.; de Oliveira, H.N. Selectivity of insecticides against adult Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae) on cassava. J. Agric. Sci. 2019, 11, 546–552. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, L.; Herz, A. Suitability of European Trichogramma species as biocontrol agents against the tomato leaf miner Tuta absoluta. Insects 2020, 11, 357. [Google Scholar] [CrossRef]
- Fenik, J.; Tankiewicz, M.; Biziuk, M. Properties and determination of pesticides in fruits and vegetables. Trac. Trend Anal. Chem. 2011, 30, 814–826. [Google Scholar] [CrossRef]
- Srinivasan, R.; Sevgan, S.; Ekesi, S.; Tamò, M. Biopesticide based sustainable pest management for safer production of vegetable legumes and brassicas in Asia and Africa. Pest. Manag. Sci. 2019, 75, 2446–2454. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, G. Review on safety of the entomopathogenic fungus Metarhizium anisopliae. Biocontrol. Sci. Technol. 2007, 17, 879–920. [Google Scholar] [CrossRef]
- Araujo, E.S.; Benatto, A.; Rizzato, F.B.; Poltronieri, A.S.; Poitevin, C.G.; Zawadneak, M.A.C.; Pimentel, I.C. Combining biocontrol agents with different mechanisms of action to control Duponchelia fovealis, an invasive pest in South America. Crop. Prot. 2020, 134, 105184. [Google Scholar] [CrossRef]
- Miranda-Fuentes, P.; Quesada-Moraga, E.; Aldebis, H.K.; Yousef-Naef, M. Compatibility between the endoparasitoid Hyposoter didymator and the entomopathogenic fungi Metarhizium brunneum: A laboratory simulation for the simultaneous use to control Spodoptera littoralis. Pest. Manag. Sci. 2020, 76, 1060–1070. [Google Scholar] [CrossRef]
- Jaronski, S.T.; Goettel, M.S.; Lomer, C.J. Regulatory requirements for ecotoxicological assessments of microbial insecticides—How relevant are they? In Environmental Impacts of Microbial Insecticides. Progress in Biological Control; Hokkanen, H.M.T., Hajek, A.E., Eds.; Springer: Dordrecht, The Netherlands, 2003; Volume 1, pp. 237–260. [Google Scholar]
- Jazzar, C.; Hammad, E.A.F. Efficacy of multiple biocontrol agents against the sweet potato whitefly Bemissia tabaci (Gennadius) (Homoptera: Aleyrodidae) on tomato. J. Appl. Entomol. 2004, 128, 188–194. [Google Scholar] [CrossRef]
- Poitevin, C.G.; Porsani, M.V.; Poltronieri, A.S.; Zawadneak, M.A.C.; Pimentel, I.C. Fungi isolated from insects in strawberry crops act as potential biological control agents of Duponchelia fovealis (Lepidoptera: Crambidae). Appl. Entomol. Zool. 2018, 53, 323–331. [Google Scholar] [CrossRef]
- Baja, F.; Poitevin, C.G.; Araujo, E.S.; Mirás-Avalos, J.M.; Zawadneak, M.A.C.; Pimentel, I.C. Infection of Beauveria bassiana and Cordyceps javanica on different immature stages of Duponchelia fovealis Zeller (Lepidoptera: Crambidae). Crop. Prot. 2020, 138, 105347. [Google Scholar] [CrossRef]
- Potrich, M.; Alves, L.F.A.; Lozano, E.; Roman, J.C.; Pietrowski, V.; Neves, P.M.O.J. Interactions between Beauveria bassiana and Trichogramma pretiosum under laboratory conditions. Entomol. Exper. Appl. 2015, 154, 213–221. [Google Scholar] [CrossRef]
- Potrich, M.; Alves, L.F.A.; Lozano, E.R.; Bonini, A.K.; Neves, P.M.O.J. Potential side effects of the entomopathogenic fungus Metarhizium anisopliae on the egg parasitoid Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) under controlled conditions. J. Econ. Entomol. 2017, 110, 2318–2324. [Google Scholar] [CrossRef]
- Campbell, B.E.; Pereira, R.M.; Koehler, P.G. Complications with controlling insect eggs. In Insecticides Resistance; Trdan, S., Ed.; IntechOpen: Rijeka, Croatia, 2016; pp. 83–96. [Google Scholar]
- Pirovani, V.D.; Pratissoli, D.; Tibúrcio, M.O.; de Carvalho, J.R.; Damascena, A.P.; Faria, L.V. Trichogramma galloi and Trichogramma pretiosum for the management of Duponchelia fovealis (Lepidoptera: Crambidae) in strawberry plants. Pesq. Agr. Bras. 2017, 52, 690–693. [Google Scholar] [CrossRef]
- Zimmermann, O. Use of Trichogramma wasps in Germany: Present status of research and commercial application of egg parasitoids against lepidopterous pests for crop. Gesunde Pflanz. 2004, 56, 157–166. [Google Scholar] [CrossRef]
- Potrich, M.; Alves, L.F.A.; Haas, J.; Da Silva, E.R.L.; Daros, A.; Pietrowski, V.; Neves, P.M.O.J. Seletividade de Beauveria bassiana e Metarhizium anisopliae a Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae). Neotrop. Entomol. 2009, 38, 822–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zawadneak, M.A.C.; Gonçalves, R.B.; Poltronieri, A.S.; Santos, B.; Bischoff, M.A.; Borba, M.A.; Pimentel, I.C. Biological parameters of Duponchelia fovealis (Lepidoptera: Crambidae) reared in the laboratory on two diets. Eur. J. Entomol. 2017, 114, 291–294. [Google Scholar] [CrossRef]
- Marchioro, C.A.; Foerster, L.A. Performance of the wheat armyworm Pseudaletia sequax Franclemont, on natural and artificial diets. Neotrop. Entomol. 2012, 41, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Ito, F.A.D.; Pimentel, I.C.; Poitevin, C.G. Niger seed agar as an inductor of sporulation of filamentous fungi with potential in biological control of agricultural diseases and pests. Int. J. Microbiol. Res. 2019, 11, 1509–1513. [Google Scholar]
- Alves, S.B.; Almeida, J.E.M.; Moino Júnior, A.; Alves, L.F.A. Controle microbiano de insetos. In Técnicas de Laboratorio; Alves, S.B., Ed.; FEALQ: Piracicaba, Brazil, 1998; pp. 637–712. [Google Scholar]
- Hassan, A.S. Métodos padronizados para testes de seletividade, com ênfase em Trichogramma. In Trichogramma e Controle Biológico Aplicado; Parra, J.R.P., Zucchi, R.A., Eds.; FEALQ: Piracicaba, Brazil, 1997; pp. 207–233. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality. Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Bartlett, M.S. Properties of sufficiency and statistical tests. Proc. R. Stat. Soc. A 1937, 160, 268–282. [Google Scholar]
- Haddad, M.L.; Vendramin, J.D. Comparação de porcentagens observadas com casos extremos de 0 e 100%. Soc. Entomol. Bras. 2000, 29, 835–837. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 21 January 2020).
- Brivio, M.F.; Mastore, M. When appearance misleads: The role of the entomopathogen surface in the relationship with its host. Insects 2020, 11, 387. [Google Scholar] [CrossRef]
- Vinson, S.B. Comportamento de seleção hospedeira de parasitoides de ovos, com ênfase na família Trichogrammatidae. In Trichogramma e Controle Biológico Aplicado; Parra, J.R.P., Zucchi, R.A., Eds.; FEALQ: Piracicaba, Brazil, 1997; pp. 67–119. [Google Scholar]
- Polanczyk, R.A.; Pratissoli, D.; Dalvi, L.P.; Grecco, E.D.; Franco, C.R. Efeito de Beauveria bassiana (Bals.) Vuillemin e Metarhizium anisopliae (Metsch.) Sorokin nos parâmetros biológicos de Trichogramma atopovirilia Oatman & Platner, 1983 (Hymenoptera: Trichogrammatidade). Cienc. Agrotec. 2010, 34, 1412–1416. [Google Scholar]
- Cônsoli, F.L.; Vinson, S.B. Parasitóides (Hymenoptera). Bioecologia e nutrição de insetos. In Base Para o Manejo Integrado de Pragas; Panizzi, A.R., Parra, J.R.P., Eds.; Embrapa Informacão Tecnológica: Brasília, Brazil, 2009; pp. 837–873. [Google Scholar]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Navarro, M.A. Trichogramma spp Producción, uso y Manejo en Colombia; Impretec: Guadalajara de Buga, Colombia, 1998; p. 176. [Google Scholar]
- Goulart, R.M.; Volpe, H.X.L.; Vacari, A.M.; Thuler, R.T.; De Bortoli, S.A. Insecticide selectivity to two species of Trichogramma in three different hosts, as determined by IOBC/WPRS methodology. Pest. Manag. Sci. 2012, 68, 240–244. [Google Scholar] [CrossRef]
- Dos Santos, V.P.; Pratissoli, D.; Paes, J.P.P.; Fragoso, D.F.M.; de Carvalho, J.R. Desempenho de Trichogramma pretiosum Riley, 1879 (Hymenoptera: Trichogrammatidae), submetido a inseticidas e fungicidas em dois hospedeiros. Rev. Ceres 2016, 63, 653–660. [Google Scholar] [CrossRef]
- Gallego, J.R.; Guerrero-Manzano, J.; Fernández-Maldonado, F.J.; Cabello, T. Susceptibility of the egg parasitoid Trichogramma achaeae (Hymenoptera: Trichogrammatidae) to selected insecticides used in tomato greenhouses. Span. J. Agric. Res. 2019, 17, e1009. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Control | B2 | B3 | Bb CG716 | Ma IBCB348 |
|---|---|---|---|---|---|
| % eggs parasitized | |||||
| T. atopovirilia | 87.2 ± 2.7 AB 1 | 89.5 ± 2.6 A | 82.3 ± 2.9 AB | 78.4 ± 2.2 B | 78.3 ± 3.3 AB |
| T. pretiosum | 70.1 ± 4.1 B | 86.8 ± 3.6 AB | 82.9 ± 3.8 AB | 82.1 ± 2.3 AB | 85.7 ± 1.9 A |
| Significance | ** | n.s. | n.s. | n.s. | n.s. |
| % emerged adults | |||||
| T. atopovirilia | 93.3 ± 2.6 AB | 85.7 ± 2.5 B | 87.1 ± 2.1 B | 96.1 ± 1.6 A | 91.5 ± 2.1 AB |
| T. pretiosum | 92.3 ± 2.4 AB | 97.3 ± 1.0 A | 87.1 ± 2.0 B | 87.7 ± 2.6 AB | 89.0 ± 2.4 AB |
| Significance | n.s. | ** | n.s. | * | n.s. |
| Sex ratio | |||||
| T. atopovirilia | 0.72 ± 0.02 A | 0.72 ± 0.01 A | 0.72 ± 0.02 A | 0.63 ± 0.03 A | 0.70 ± 0.02 A |
| T. pretiosum | 0.71 ± 0.03 A | 0.69 ± 0.02 A | 0.77 ± 0.01 A | 0.73 ± 0.01 A | 0.74 ± 0.02 A |
| Significance | n.s. | n.s. | n.s. | n.s. | n.s. |
| Species | Control | B2 | B3 | Bb CG716 | Ma IBCB348 |
|---|---|---|---|---|---|
| % eggs parasitized | |||||
| T. atopovirilia | 89.8 ± 1.1 A 1 | 81.3 ± 1.9 A | 83.3 ± 2.2 A | 90.2 ± 2.1 A | 85.1 ± 3.5 A |
| T. pretiosum | 80.7 ± 4.3 A | 90.9 ± 1.8 A | 84.4 ± 2.6 A | 88.2 ± 2.3 A | 80.8 ± 3.2 A |
| Significance | n.s. | n.s. | n.s. | n.s. | n.s. |
| % emerged adults | |||||
| T. atopovirilia | 93.5 ± 1.8 A | 75.8 ± 1.7 B | 91.4 ± 2.4 A | 93.1 ± 2.0 A | 92.7 ± 2.2 A |
| T. pretiosum | 93.3 ± 2.3 A | 91.8 ± 1.9 A | 93.7 ± 2.3 A | 95.6 ± 2.2 A | 93.3 ± 1.7 A |
| Significance | n.s. | n.s. | n.s. | n.s. | n.s. |
| Sex ratio | |||||
| T. atopovirilia | 0.66 ± 0.02 A | 0.72 ± 0.01 A | 0.65 ± 0.01 A | 0.70 ± 0.01 A | 0.69 ± 0.03 A |
| T. pretiosum | 0.95 ± 0.02 A | 0.66 ± 0.01 C | 0.70 ± 0.02 BC | 0.77 ± 0.03 BC | 0.80 ± 0.03 B |
| Significance | *** | ** | n.s. | * | * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araujo, E.S.; Poltronieri, A.S.; Poitevin, C.G.; Mirás-Avalos, J.M.; Zawadneak, M.A.C.; Pimentel, I.C. Compatibility between Entomopathogenic Fungi and Egg Parasitoids (Trichogrammatidae): A Laboratory Study for Their Combined Use to Control Duponchelia fovealis. Insects 2020, 11, 630. https://doi.org/10.3390/insects11090630
Araujo ES, Poltronieri AS, Poitevin CG, Mirás-Avalos JM, Zawadneak MAC, Pimentel IC. Compatibility between Entomopathogenic Fungi and Egg Parasitoids (Trichogrammatidae): A Laboratory Study for Their Combined Use to Control Duponchelia fovealis. Insects. 2020; 11(9):630. https://doi.org/10.3390/insects11090630
Chicago/Turabian StyleAraujo, Emily Silva, Alex S. Poltronieri, Carolina G. Poitevin, José Manuel Mirás-Avalos, Maria Aparecida Cassilha Zawadneak, and Ida Chapaval Pimentel. 2020. "Compatibility between Entomopathogenic Fungi and Egg Parasitoids (Trichogrammatidae): A Laboratory Study for Their Combined Use to Control Duponchelia fovealis" Insects 11, no. 9: 630. https://doi.org/10.3390/insects11090630
APA StyleAraujo, E. S., Poltronieri, A. S., Poitevin, C. G., Mirás-Avalos, J. M., Zawadneak, M. A. C., & Pimentel, I. C. (2020). Compatibility between Entomopathogenic Fungi and Egg Parasitoids (Trichogrammatidae): A Laboratory Study for Their Combined Use to Control Duponchelia fovealis. Insects, 11(9), 630. https://doi.org/10.3390/insects11090630