
Joint Impacts of Drought and Habitat Fragmentation on Native Bee Assemblages in a California Biodiversity Hotspot



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
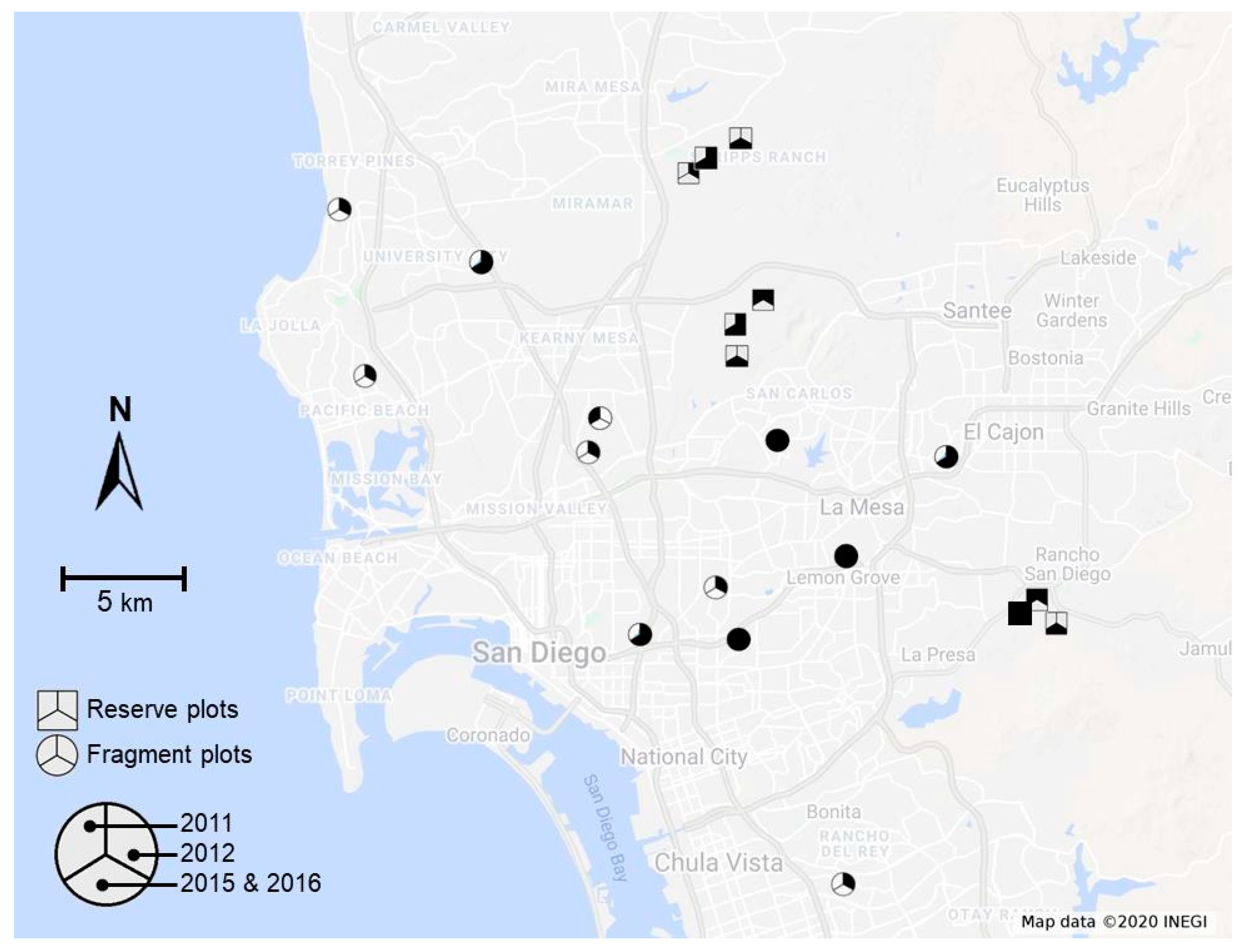
2.1. Study System
2.2. Data Collection
2.3. Statistical Analyses
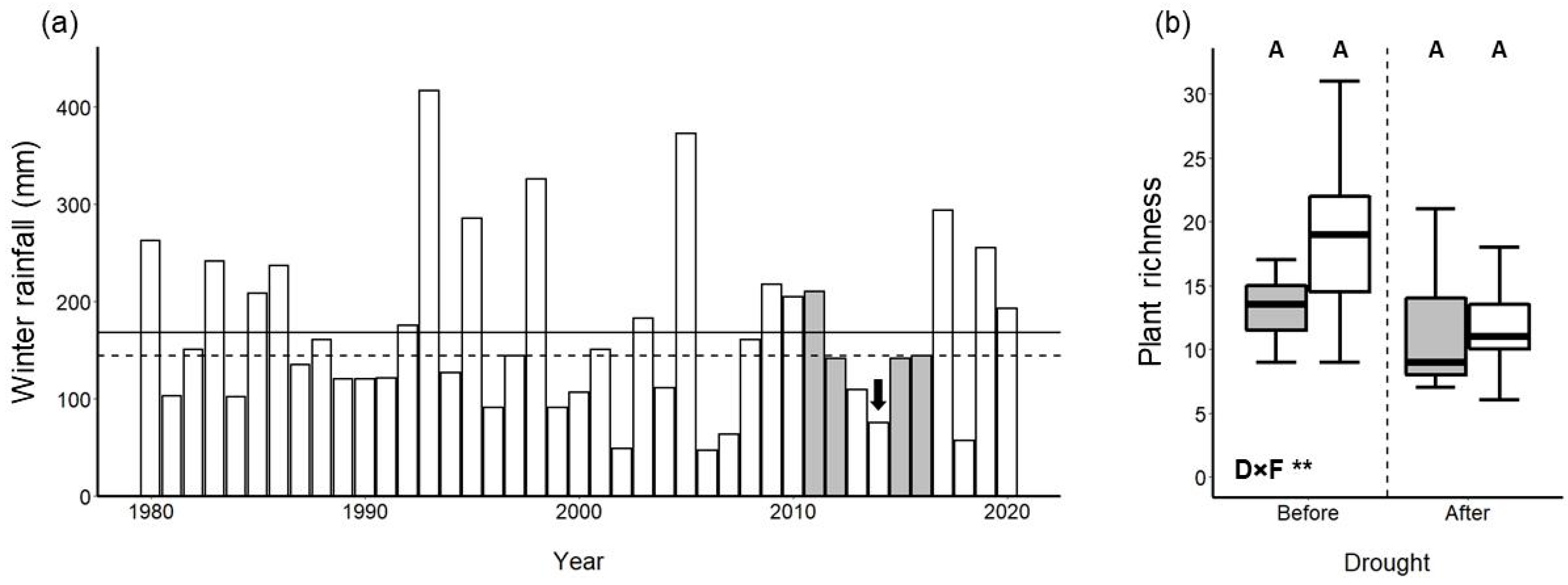
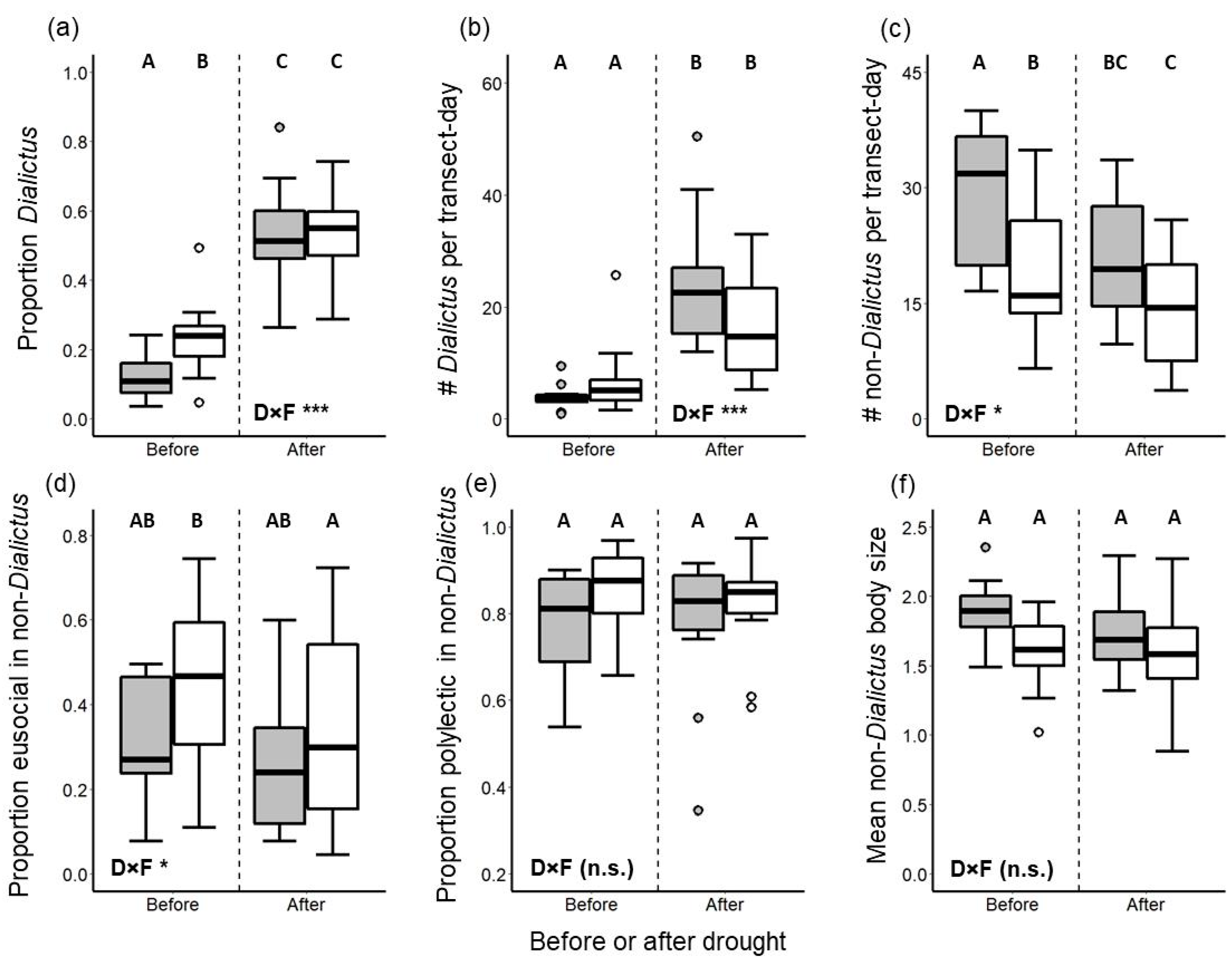
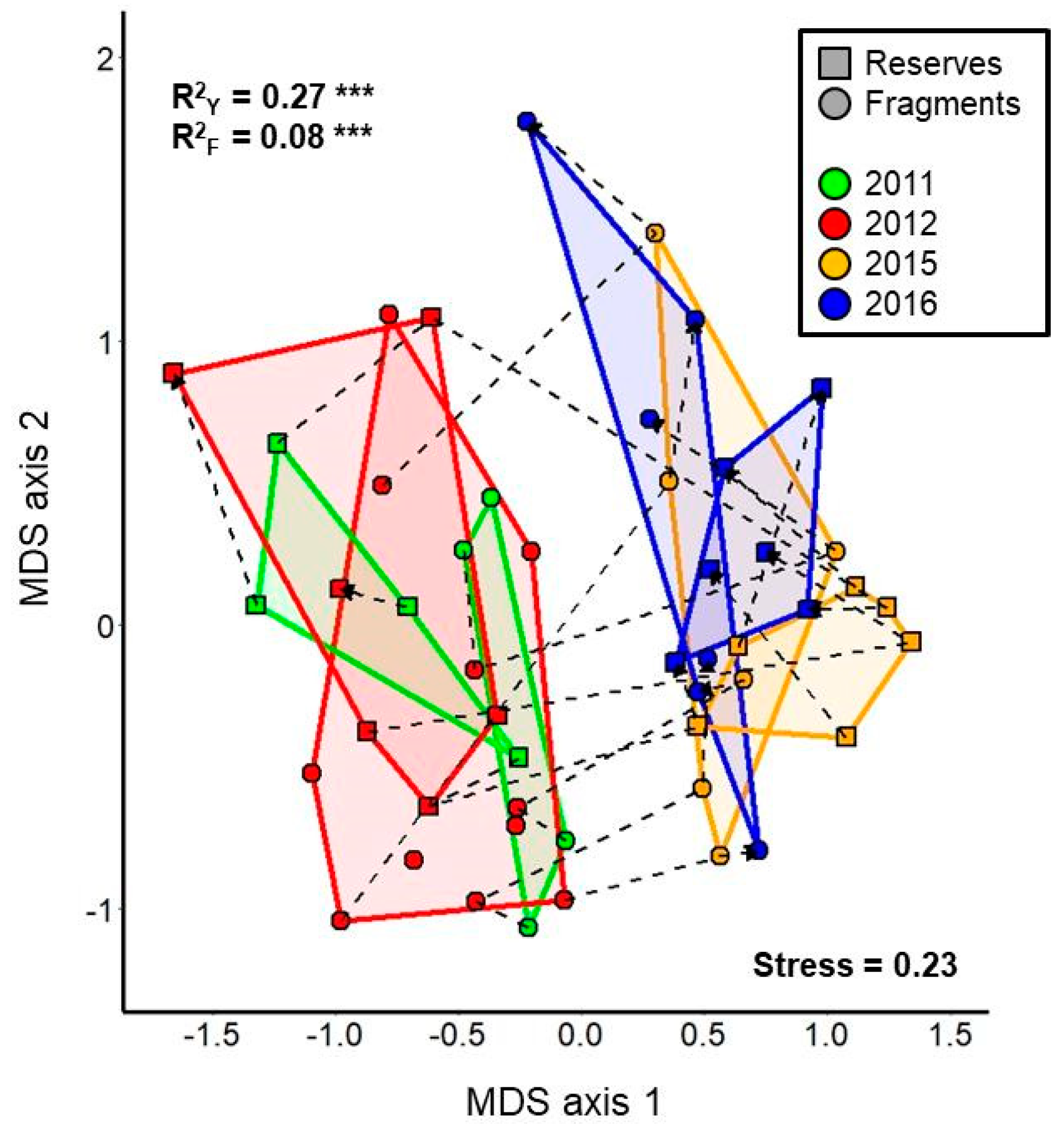
3. Results
3.1. Univariate Analyses
3.2. Multivariate Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Palmer, G.; Platts, P.J.; Brereton, T.; Chapman, J.W.; Dytham, C.; Fox, R.; Pearce-Higgins, J.W.; Roy, D.B.; Hill, J.K.; Thomas, C.D. Climate change, climatic variation and extreme biological responses. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160144. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, G.M. Severe and sustained drought in Southern California and the West: Present conditions and insights from the past on causes and impacts. Quat. Int. 2007, 173–174, 87–100. [Google Scholar] [CrossRef]
- Christie, D.A.; Boninsegna, J.A.; Cleaveland, M.K.; Lara, A.; Le Quesne, C.; Morales, M.S.; Mudelsee, M.; Stahle, D.W.; Villalba, R. Aridity changes in the Temperate-Mediterranean transition of the Andes since AD 1346 reconstructed from tree-rings. Clim. Dyn. 2011, 36, 1505–1521. [Google Scholar] [CrossRef]
- Okin, G.S.; Dong, C.; Willis, K.S.; Gillespie, T.W.; MacDonald, G.M. The impact of drought on native Southern California vegetation: Remote sensing analysis using MODIS-derived time series. J. Geophys. Res. Biogeosci. 2018, 123, 1927–1939. [Google Scholar] [CrossRef]
- Tsesmelis, D.; Karavitis, C.; Oikonomou, P.; Alexandris, S.; Kosmas, C. Assessment of the vulnerability to drought and desertification characteristics using the Standardized Drought Vulnerability Index (SDVI) and the Environmentally Sensitive Areas Index (ESAI). Resources 2018, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, M.T.K.; Robles, V.; Tamburrino, Í.; Martínez-Harms, J.; Garreaud, R.D.; Jara-Arancio, P.; Pliscoff, P.; Copier, A.; Arenas, J.; Keymer, J.; et al. Extreme drought affects visitation and seed set in a plant species in the central Chilean Andes heavily dependent on hummingbird pollination. Plants 2020, 9, 1553. [Google Scholar] [CrossRef]
- Berdanier, A.B.; Clark, J.S. Multiyear drought-induced morbidity preceding tree death in southeastern U.S. forests. Ecol. Appl. 2016, 26, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Sperry, J.H.; Weatherhead, P.J. Prey-mediated effects of drought on condition and survival of a terrestrial snake. Ecology 2008, 89, 2770–2776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tariq, M.; Rossiter, J.T.; Wright, D.J.; Staley, J.T. Drought alters interactions between root and foliar herbivores. Oecologia 2013, 172, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Descamps, C.; Quinet, M.; Jacquemart, A.-L. The effects of drought on plant–pollinator interactions: What to expect? Environ. Exp. Bot. 2021, 182, 104297. [Google Scholar] [CrossRef]
- Phillips, B.B.; Shaw, R.F.; Holland, M.J.; Fry, E.L.; Bardgett, R.D.; Bullock, J.M.; Osborne, J.L. Drought reduces floral resources for pollinators. Glob. Chang. Biol. 2018, 24, 3226–3235. [Google Scholar] [CrossRef]
- Waser, N.M.; Price, M.V. Drought, pollen and nectar availability, and pollination success. Ecology 2016, 97, 1400–1409. [Google Scholar] [CrossRef] [Green Version]
- Burkle, L.A.; Runyon, J.B. Drought and leaf herbivory influence floral volatiles and pollinator attraction. Glob. Chang. Biol. 2016, 22, 1644–1654. [Google Scholar] [CrossRef]
- Yurkonis, K.A.; Meiners, S.J. Drought impacts and recovery are driven by local variation in species turnover. Plant Ecol. 2006, 184, 325–336. [Google Scholar] [CrossRef]
- Mayer, C.; Kuhlmann, M. Synchrony of pollinators and plants in the winter rainfall area of South Africa—Observations from a drought year. Trans. R. Soc. S. Afr. 2004, 59, 55–57. [Google Scholar] [CrossRef]
- Minckley, R.L.; Roulston, T.H.; Williams, N.M. Resource assurance predicts specialist and generalist bee activity in drought. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122703. [Google Scholar] [CrossRef] [Green Version]
- Griffin, D.; Anchukaitis, K.J. How unusual is the 2012-2014 California drought? Geophys. Res. Lett. 2014, 41, 9017–9023. [Google Scholar] [CrossRef] [Green Version]
- Hung, K.-L.J. Effects of Habitat Fragmentation and Introduced Species on the Structure and Function of Plant-Pollinator Interactions. Ph.D. Thesis, University of California, San Diego, CA, USA, 2017. [Google Scholar]
- Hung, K.J.; Ascher, J.S.; Davids, J.A.; Holway, D.A. Ecological filtering in scrub fragments restructures the taxonomic and functional composition of native bee assemblages. Ecology 2019, 100, e02654. [Google Scholar] [CrossRef] [PubMed]
- Weiner, C.N.; Werner, M.; Linsenmair, K.E.; Blüthgen, N. Land-use impacts on plant-pollinator networks: Interaction strength and specialization predict pollinator declines. Ecology 2014, 95, 466–474. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Choate, B.A. Wild bee species abundance and richness across an urban-rural gradient. J. Insect Conserv. 2018, 22, 391–403. [Google Scholar] [CrossRef]
- Minckley, R.L.; Cane, J.H.; Kervin, L. Origins and ecological consequences of pollen specialization among desert bees. Proc. R. Soc. Lond. B Biol. Sci. 2000, 267, 265–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaluza, B.F.; Wallace, H.M.; Heard, T.A.; Minden, V.; Klein, A.; Leonhardt, S.D. Social bees are fitter in more biodiverse environments. Sci. Rep. 2018, 8, 12353. [Google Scholar] [CrossRef] [Green Version]
- Requier, F.; Jowanowitsch, K.K.; Kallnik, K.; Steffan-Dewenter, I. Limitation of complementary resources affects colony growth, foraging behavior, and reproduction in bumble bees. Ecology 2020, 101, e02946. [Google Scholar] [CrossRef]
- Michener, C.D. The Bees of the World, 2nd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2007; ISBN 978-0-8018-8573-0. [Google Scholar]
- Dornhaus, A.; Chittka, L. Information flow and regulation of foraging activity in bumble bees (Bombus spp.). Apidologie 2004, 35, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Greenleaf, S.S.; Williams, N.M.; Winfree, R.; Kremen, C. Bee foraging ranges and their relationship to body size. Oecologia 2007, 153, 589–596. [Google Scholar] [CrossRef]
- Koerner, S.E.; Collins, S.L. Interactive effects of grazing, drought, and fire on grassland plant communities in North America and South Africa. Ecology 2014, 95, 98–109. [Google Scholar] [CrossRef] [Green Version]
- Thomson, D.M. Local bumble bee decline linked to recovery of honey bees, drought effects on floral resources. Ecol. Lett. 2016, 19, 1247–1255. [Google Scholar] [CrossRef]
- Soulé, M.E.; Alberts, A.C.; Bolger, D.T. The effects of habitat fragmentation on chaparral plants and vertebrates. Oikos 1992, 63, 39–47. [Google Scholar] [CrossRef]
- Gotlieb, A.; Hollender, Y.; Mandelik, Y. Gardening in the desert changes bee communities and pollination network characteristics. Basic Appl. Ecol. 2011, 12, 310–320. [Google Scholar] [CrossRef]
- Ghahremaninejad, F.; Hoseini, E.; Fereidounfar, S. Cities in drylands as artificial protected areas for plants. Biodivers. Conserv. 2021, 30, 243–248. [Google Scholar] [CrossRef]
- Wojcik, V.A.; Buchmann, S. Pollinator conservation and management on electrical transmission and roadside rights-of-way: A review. J. Pollinat. Ecol. 2012, 7, 16–26. [Google Scholar] [CrossRef]
- McCann, K.S. The diversity-stability debate. Nature 2000, 405, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Elmqvist, T.; Folke, C.; Nyström, M.; Peterson, G.; Bengtsson, J.; Walker, B.; Norberg, J. Response diversity, ecosystem change, and resilience. Front. Ecol. Environ. 2003, 1, 488–494. [Google Scholar] [CrossRef]
- Rebman, J.P.; Simpson, M.G. Checklist of the Vascular Plants of San Diego County, 5th ed.; San Diego Natural History Museum: San Diego, CA, USA, 2014; ISBN 978-0-918969-08-8. [Google Scholar]
- Moldenke, A.R.; Neff, J.L. The Bees of California, a Catalogue with Special Reference to Pollination and Ecological Research; Origin and Structure of Ecosystems; Technical Reports; University of California Press: Santa Cruz, CA, USA, 1974. [Google Scholar]
- Michener, C.D. Biogeography of the bees. Ann. Mo. Bot. Gard. 1979, 66, 277–347. [Google Scholar] [CrossRef]
- Prugh, L.R.; Deguines, N.; Grinath, J.B.; Suding, K.N.; Bean, W.T.; Stafford, R.; Brashares, J.S. Ecological winners and losers of extreme drought in California. Nat. Clim. Chang. 2018, 8, 819–824. [Google Scholar] [CrossRef] [Green Version]
- Westman, W.E. Diversity relations and succession in Californian coastal sage scrub. Ecology 1981, 62, 170–184. [Google Scholar] [CrossRef]
- Bolger, D.T.; Alberts, A.C.; Soulé, M.E. Occurrence patterns of bird species in habitat fragments: Sampling, extinction, and nested species subsets. Am. Nat. 1991, 137, 155–166. [Google Scholar] [CrossRef]
- Bolger, D.T.; Beard, K.H.; Suarez, A.V.; Case, T.J. Increased abundance of native and non-native spiders with habitat fragmentation. Divers. Distrib. 2008, 14, 655–665. [Google Scholar] [CrossRef]
- Suarez, A.V.; Bolger, D.T.; Case, T.J. Effects of fragmentation and invasion on native ant communities in coastal Southern California. Ecology 1998, 79, 2041–2056. [Google Scholar] [CrossRef]
- Kam, J.; Stowers, K.; Kim, S. Monitoring of drought awareness from Google trends: A case study of the 2011–17 California drought. Weather. Clim. Soc. 2019, 11, 419–429. [Google Scholar] [CrossRef]
- Portman, Z.M.; Bruninga-Socolar, B.; Cariveau, D.P. The state of bee monitoring in the United States: A call to refocus away from bowl traps and towards more effective methods. Ann. Entomol. Soc. Am. 2020, 113, 337–342. [Google Scholar] [CrossRef]
- Prendergast, K.S.; Menz, M.H.M.; Dixon, K.W.; Bateman, P.W. The relative performance of sampling methods for native bees: An empirical test and review of the literature. Ecosphere 2020, 11, e03076. [Google Scholar] [CrossRef]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyörgyi, H.; Tscheulin, T.; et al. Measuring bee diversity in different European habitats and biogeographical regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef] [Green Version]
- Hung, K.-L.J.; Ascher, J.S.; Holway, D.A. Urbanization-induced habitat fragmentation erodes multiple components of temporal diversity in a Southern California native bee assemblage. PLoS ONE 2017, 12, e0184136. [Google Scholar] [CrossRef] [Green Version]
- Cane, J.H. Estimation of bee size using intertegular span (Apoidea). J. Kans. Entomol. Soc. 1987, 60, 145–147. [Google Scholar]
- Potts, S.G.; Vulliamy, B.; Dafni, A.; Ne’eman, G.; Willmer, P. Linking bees and flowers: How do floral communities structure pollinator communities? Ecology 2003, 84, 2628–2642. [Google Scholar] [CrossRef] [Green Version]
- Carson, B.D.; Bahlai, C.A.; Gibbs, J.; Landis, D.A. Flowering phenology influences bee community dynamics in old fields dominated by the invasive plant Centaurea stoebe. Basic Appl. Ecol. 2016, 17, 497–507. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020.
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.-Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill Numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Williams, N.M.; Minckley, R.L.; Silveira, F.A. Variation in native bee faunas and its implications for detecting community changes. Conserv. Ecol. 2001, 5, 7. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 11 April 2020).
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef]
- Anderson, M.J.; Crist, T.O.; Chase, J.M.; Vellend, M.; Inouye, B.D.; Freestone, A.L.; Sanders, N.J.; Cornell, H.V.; Comita, L.S.; Davies, K.F.; et al. Navigating the multiple meanings of β diversity: A roadmap for the practicing ecologist. Ecol. Lett. 2011, 14, 19–28. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2018. Available online: http://CRAN.R-project.org/package=vegan (accessed on 13 December 2019).
- Martinez Arbizu, P. pairwiseAdonis: Pairwise Multilevel Comparison Using Adonis. 2017. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 13 August 2020).
- Howell, A.D.; Alarcón, R.; Minckley, R.L. Effects of habitat fragmentation on the nesting dynamics of desert bees. Ann. Entomol. Soc. Am. 2017, 110, 233–243. [Google Scholar] [CrossRef]
- Cane, J.H.; Minckley, R.L.; Kervin, L.J.; Roulston, T.H.; Williams, N.M. Complex responses within a desert bee guild (Hymenoptera: Apiformes) to urban habitat fragmentation. Ecol. Appl. 2006, 16, 632–644. [Google Scholar] [CrossRef] [Green Version]
- Wray, J.C.; Elle, E. Flowering phenology and nesting resources influence pollinator community composition in a fragmented ecosystem. Landsc. Ecol. 2015, 30, 261–272. [Google Scholar] [CrossRef]
- Hall, D.M.; Camilo, G.R.; Tonietto, R.K.; Ollerton, J.; Ahrne, K.; Arduser, M.; Ascher, J.S.; Baldock, K.C.R.; Fowler, R.; Frankie, G.; et al. The city as a refuge for insect pollinators. Conserv. Biol. 2017, 31, 24–29. [Google Scholar] [CrossRef]
- Majewska, A.A.; Altizer, S. Planting gardens to support insect pollinators. Conserv. Biol. 2020, 34, 15–25. [Google Scholar] [CrossRef]
- Carril, O.M.; Griswold, T.; Haefner, J.; Wilson, J.S. Wild bees of Grand Staircase-Escalante National Monument: Richness, abundance, and spatio-temporal beta-diversity. PeerJ 2018, 6, e5867. [Google Scholar] [CrossRef] [PubMed]
- Meiners, J.M.; Griswold, T.L.; Carril, O.M. Decades of native bee biodiversity surveys at Pinnacles National Park highlight the importance of monitoring natural areas over time. PLoS ONE 2019, 14, e0207566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minckley, R.L. Faunal composition and species richness differences of bees (Hymenoptera: Apiformes) from two North American regions. Apidologie 2008, 39, 176–188. [Google Scholar] [CrossRef] [Green Version]
- Tonietto, R.; Fant, J.; Ascher, J.; Ellis, K.; Larkin, D. A comparison of bee communities of Chicago green roofs, parks and prairies. Landsc. Urban Plan. 2011, 103, 102–108. [Google Scholar] [CrossRef]
- Lowenstein, D.M. Evaluating the dependence of urban pollinators on ornamental, non-native, and ‘weedy’ floral resources. Urban Ecosyst. 2019, 22, 293–302. [Google Scholar] [CrossRef]
- Bommarco, R.; Biesmeijer, J.C.; Meyer, B.; Potts, S.G.; Poyry, J.; Roberts, S.P.M.; Steffan-Dewenter, I.; Ockinger, E. Dispersal capacity and diet breadth modify the response of wild bees to habitat loss. Proc. R. Soc. B Biol. Sci. 2010, 277, 2075–2082. [Google Scholar] [CrossRef] [Green Version]
- Selwood, K.E.; McGeoch, M.A.; Mac Nally, R. The effects of climate change and land-use change on demographic rates and population viability. Biol. Rev. 2015, 90, 837–853. [Google Scholar] [CrossRef] [Green Version]
- Kammerer, M.; Goslee, S.C.; Douglas, M.R.; Tooker, J.F.; Grozinger, C.M. Wild bees as winners and losers: Relative impacts of landscape composition, quality, and climate. Glob. Chang. Biol. 2021, gcb.15485. [Google Scholar] [CrossRef] [PubMed]
- McKinney, M.L.; Lockwood, J.L. Biotic homogenization: A few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 1999, 14, 450–453. [Google Scholar] [CrossRef]
- Danforth, B.N. Emergence dynamics and bet hedging in a desert bee, Perdita portalis. Proc. R. Soc. Lond. B Biol. Sci. 1999, 266, 1985–1994. [Google Scholar] [CrossRef]
- Woodard, S.H.; Federman, S.; James, R.R.; Danforth, B.N.; Griswold, T.L.; Inouye, D.; McFrederick, Q.S.; Morandin, L.; Paul, D.L.; Sellers, E.; et al. Towards a U.S. national program for monitoring native bees. Biol. Conserv. 2020, 252, 108821. [Google Scholar] [CrossRef]
- Orr, M.C.; Hughes, A.C.; Chesters, D.; Pickering, J.; Zhu, C.-D.; Ascher, J.S. Global patterns and drivers of bee distribution. Curr. Biol. 2020, S0960982220315967. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison | Type | Test Statistic | R2 | p Value |
|---|---|---|---|---|
| 2011 versus 2015 | Pre-Post | F1,17 = 7.88 | 0.29 | 0.0015 |
| 2011 versus 2016 | Pre-Post | F1,17 = 5.60 | 0.23 | 0.0015 |
| 2012 versus 2015 | Pre-Post | F1,26 = 11.23 | 0.28 | 0.0015 |
| 2012 versus 2016 | Pre-Post | F1,26 = 7.87 | 0.21 | 0.0015 |
| 2011 versus 2012 | Pre-Pre | F1,22 = 2.54 | 0.09 | 0.011 |
| 2015 versus 2016 | Post-Post | F1,21 = 1.70 | 0.07 | 0.083 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hung, K.-L.J.; Sandoval, S.S.; Ascher, J.S.; Holway, D.A. Joint Impacts of Drought and Habitat Fragmentation on Native Bee Assemblages in a California Biodiversity Hotspot. Insects 2021, 12, 135. https://doi.org/10.3390/insects12020135
Hung K-LJ, Sandoval SS, Ascher JS, Holway DA. Joint Impacts of Drought and Habitat Fragmentation on Native Bee Assemblages in a California Biodiversity Hotspot. Insects. 2021; 12(2):135. https://doi.org/10.3390/insects12020135
Chicago/Turabian StyleHung, Keng-Lou James, Sara S. Sandoval, John S. Ascher, and David A. Holway. 2021. "Joint Impacts of Drought and Habitat Fragmentation on Native Bee Assemblages in a California Biodiversity Hotspot" Insects 12, no. 2: 135. https://doi.org/10.3390/insects12020135
APA StyleHung, K.-L. J., Sandoval, S. S., Ascher, J. S., & Holway, D. A. (2021). Joint Impacts of Drought and Habitat Fragmentation on Native Bee Assemblages in a California Biodiversity Hotspot. Insects, 12(2), 135. https://doi.org/10.3390/insects12020135