
Preliminary Report on the Acquisition, Persistence, and Potential Transmission of Citrus tristeza virus by Diaphorina citri

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. CTV Orchard Surveys
2.2. Plant and Insect Materials under Laboratory Conditions
2.3. Acquisition and Persistence of CTV in D. citri
2.4. Transmission of CTV by D. citri to Mandarin, Lemon, and Orange Jasmine Plants
2.5. RNA Extraction and cDNA Synthesis
2.6. CTV Detection
2.6.1. Conventional PCR
2.6.2. Reverse Transcription Real-Time Quantitative PCR (RT-qPCR)
2.6.3. CTV Genome Detection
2.6.4. Enzyme-Linked Immunosorbent Assays (ELISA)
2.7. Statistical Analyses
3. Results
3.1. CTV Orchard Surveys
3.2. Genetic Characterization of CTV in D. citri
3.3. CTV Persistence Period Inside D. citri
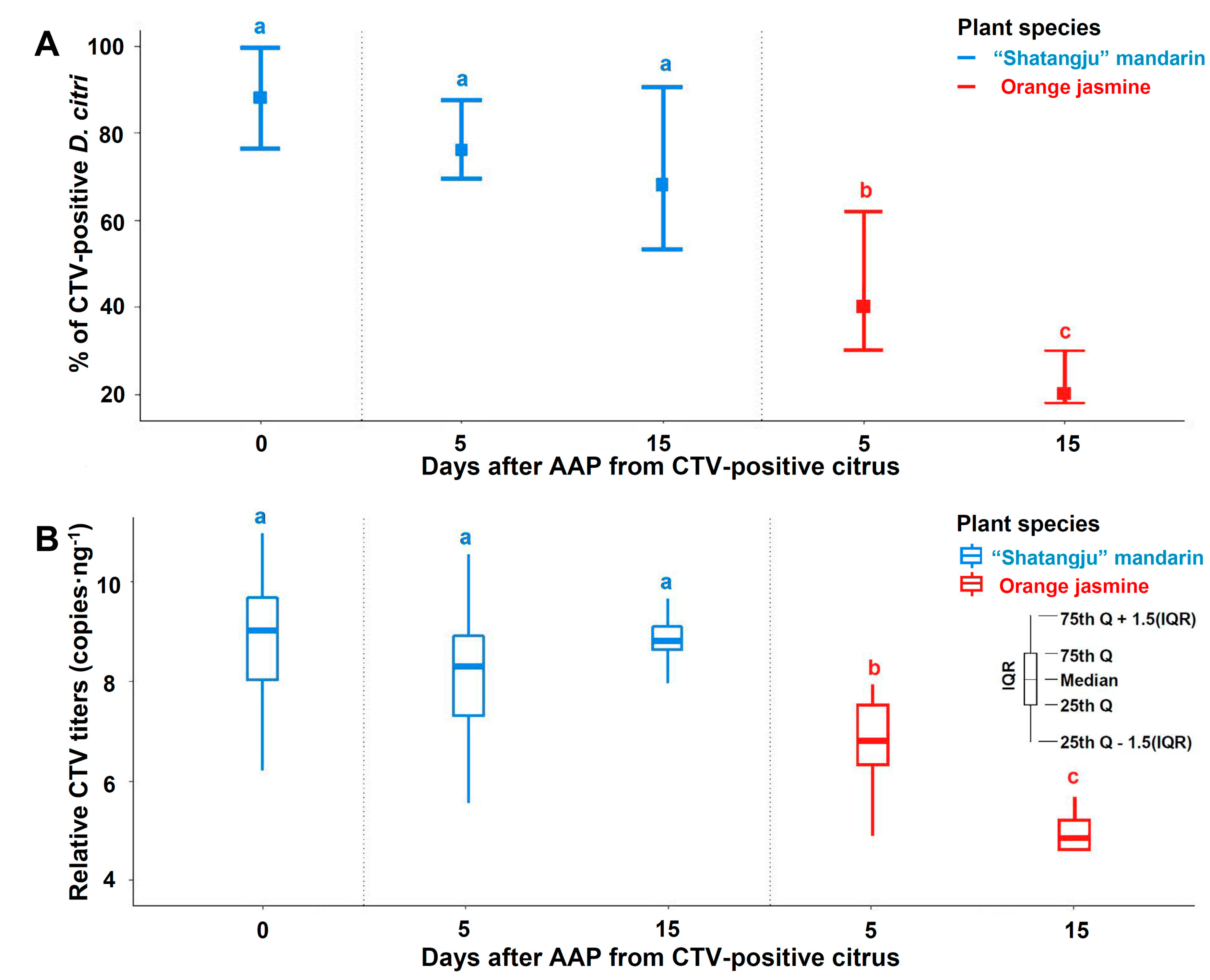
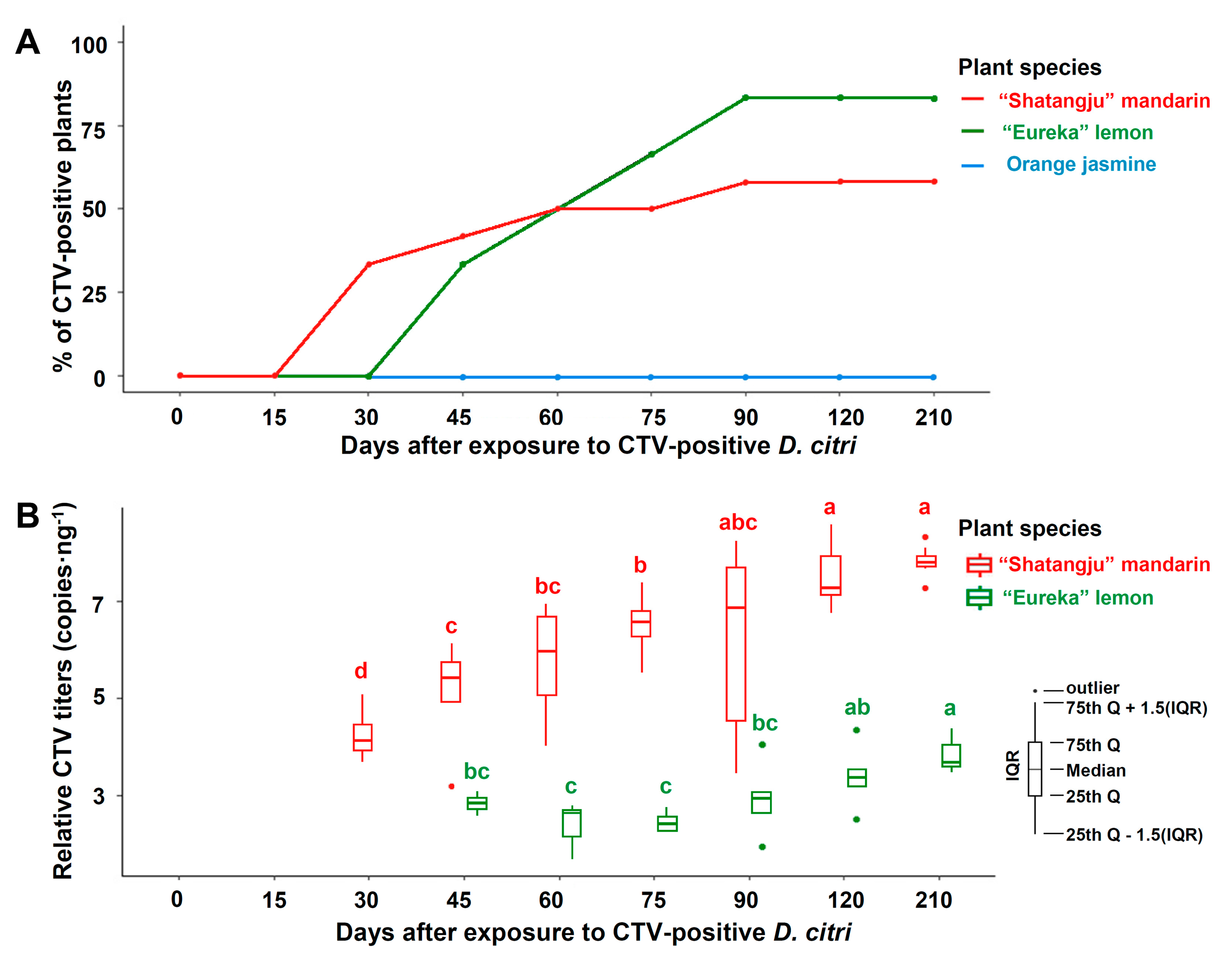
3.4. Transmission of CTV by D. citri to Mandarin, Lemon, and Orange Jasmine Plants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rocha-Peña, M.A.; Lee, R.F.; Lastra, R.; Niblett, C.L.; Ochoa-Corona, F.M.; Garnsey, S.M.; Yokomi, R.K. Citrus tristeza virus and its aphid vector Toxoptera citricida: Threats to citrus production in the Carbbean and central and North America. Plant Dis. 1995, 79, 437–445. [Google Scholar] [CrossRef]
- Moreno, P.; Ambrós, S.; Albiach-Marti, M.; Guerri, J.; Peña, L. Citrus tristeza virus: A pathogen that changed the course of the citrus industry. Mol. Plant Path. 2008, 9, 251–268. [Google Scholar] [CrossRef]
- Cambra, M.; Gorris, M.T.; Marroquı́n, C.; Román, M.P.; Olmos, A.; Martı́nez, M.C.; de Mendoza, A.H.; López, A.; Nacarro, L. Incidence and epidemiology of Citrus tristeza virus in the Valencian Community of Spain. Virus Res. 2000, 71, 85–95. [Google Scholar] [CrossRef]
- Atta, S.; Zhou, C.; Zhou, Y.; Cao, M.; Wang, X. Distribution and research advances of Citrus tristeza virus. J. Integr. Agric. 2012, 11, 346–358. [Google Scholar] [CrossRef]
- McClean, A.P.D. Tristeza and stem pitting diseases of citrus in South Africa. FAO. Plant Protect. Bull. 1956, 4, 88–94. [Google Scholar]
- Roistacher, C.N.; Bar-Joseph, M. Aphid transmission of Citrus tristeza virus: A review. Phytophylactica 1987, 19, 163–167. [Google Scholar]
- Roistacher, C.N.; Bar-Joseph, M. Transmission of Citrus tristeza virus (CTV) by Aphis gossypii and by graft inoculation to and from Passiflora spp. Phytophylactica 1987, 19, 179–182. [Google Scholar]
- Zhao, X.; Jiang, Y.; Zhang, Q.; Qiu, Z. Distribution of the seedling yellow tristeza and the tristeza susceptibility of 6 sour orange stocks. Acta Phytopathol. Sin. 1979, 9, 61–63. [Google Scholar]
- Zhou, Y.; Zhou, C.; Li, Z.; Wang, X.; Liu, K. Mild strains cross protection against stem-pitting tristeza of sweet orange. Sci. Agric. Sin. 2008, 41, 4085–4091. [Google Scholar]
- Chen, J.; Chen, Q.; Wu, Y.; Zhao, X.; Chen, Z. Investigation of Citrus tristeza virus with stempitting symptom among varrious citrus genotypes. China Citrus 1992, 21, 26–28. [Google Scholar]
- Zhou, C.; Zhao, X.; Jiang, Y.; Dai, S.; Tang, K.; Yang, F.; Huang, S.; Yang, Y.; Li, Z.; Wang, Z.; et al. Investigation and identification of pummelo dwarf disease. South China Fruits 1998, 27, 20–21. [Google Scholar]
- Lu, Z. The Influence of Various Citrus Genotype on Isolation of Citrus tristeza virus. Master’s Thesis, Southwest University, Chongqing, China, 2009. [Google Scholar]
- Tao, Z.; Yi, L.; Lu, Z.; Li, F. Investigation and research on the infection rate of Citrus tristeza virus for major production regions of Gannan’s navel oranges. Chin. Agric. Sci. Bull. 2011, 27, 297–300. [Google Scholar]
- Bar-Joseph, M.; Garnsey, S.M.; Gonsalves, D. The closteroviruses: A distinct group of enlong plant virus. Adv. Virus Res. 1979, 25, 93–168. [Google Scholar]
- Chen, A.Y.S.; Watanabe, S.; Yokomi, R.; Ng, J.C.K. Nucleotide heterogeneity at the terminal ends of the genomes of two California Citrus tristeza virus strains and their complete genome sequence analysis. Virol. J. 2018, 15, 141. [Google Scholar] [CrossRef]
- Karasev, A.V.; Boyko, V.P.; Gowda, S.; Nikolaeva, O.V.; Hilf, M.E.; Koonin, E.V.; Niblett, C.L.; Cline, K.; Gumpf, D.J.; Lee, R.F.; et al. Complete sequence of the Citrus tristeza virus RNA genome. Virology 1995, 208, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Harper, S.J. Citrus tristeza virus: Evolution of complex and varied genotypic groups. Front. Microbiol. 2013, 4, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Bar-Joseph, M.; Garnsey, S.M.; Gonsalves, D.; Moscovitz, M.; Purcifull, D.E.; Clark, M.F.; Loebenstein, G. The use of enzyme-linked immunosorbent assay for the detection of Citrus tristeza virus. Phytopathology 1979, 69, 190–194. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Z.; Cao, Y.; Xia, Y.; Yin, Y. Detection of Citrus tristeza virus using conventional and fluorescence quantitative RT-PCR assays. Acta. Phytopathol. Sin. 2008, 1, 12. [Google Scholar]
- Fawcett, H.S.; Wallace, J.M. Evidence of the virus nature of citrus quick decline. Calif. Citrogr. 1946, 32, 88–89. [Google Scholar]
- Bar-Joseph, M.; Raccah, B.; Loebenstein, G. Evaluation of the main variables that affect Citrus tristeza virus transmission by aphids. Proc. Int. Soc. Citric. 1977, 3, 958–961. [Google Scholar]
- Bar-Joseph, M.; Marcu, S.R.; Lee, R. The continuing challenge of Citrus tristeza virus control. Ann. Rev. Phytopath. 1989, 27, 291–316. [Google Scholar] [CrossRef]
- Yokomi, R.K.; Garnsey, S.M.; Civerolo, E.L.; Gumpf, D.J. Transmission of exotic Citrus tristeza virus isolates by a Florida colony. Plant. Dis. 1989, 73, 552–556. [Google Scholar] [CrossRef]
- Yokomi, R.K.; Lastra, R.; Stoetzel, M.B.; Damsteegt, V.D.; Lee, R.F.; Garnsey, S.M.; Gottwald, T.R.; Rocha-Pena, M.A.; Niblett, C.L. Establishment of the brown citrus aphid (Homoptera: Aphididae) in Central America and the Caribbean basin and transmission of Citrus tristeza virus. J. Econ. Entomol. 1994, 87, 1078–1085. [Google Scholar] [CrossRef]
- Bové, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Hall, D.G.; Richardson, M.L.; Ammar, E.-D.; Halbert, S.E. Asian citrus psyllid, Diaphorina citri, vector of citrus huanglongbing disease. Entomol. Exp. Appl. 2013, 146, 207–223. [Google Scholar] [CrossRef]
- Capoor, S.P.; Rao, S.P.; Viswanath, S.M. Diaphorina citri Kuway, a vector of the greening disease of citrus in India. Indian J. Agric. Sci. 1967, 37, 572–576. [Google Scholar]
- Martinez, A.L.; Wallace, J.M. Citrus leaf-mottle-yellows disease in the Philippines and transmission of the causal virus by a psyllid, Diaphorina citri. Philipp. J. Plant Indus. 1967, 51, 119–125. [Google Scholar]
- Guangdong Agriculture and Forestry College. Preliminary report of Huanglongbing transmission by citrus psyllid. Guangdong Agric. Sci. 1977, 6, 50–53. [Google Scholar]
- Guangxi Huanglongbing Research Group. Citrus psyllids and citrus Huanglongbing. Citrus Sci. Technol. Comm. 1977, 2, 23–24. [Google Scholar]
- Hung, T.H.; Hung, S.C.; Chen, C.N.; Hsu, M.H.; Su, H.J. Detection by PCR of Candidatus Liberibacter asiaticus, the bacterium causing citrus Huanglongbing in vector psyllids: Application to the study of vector–pathogen relationships. Plant Pathol. 2004, 53, 96–102. [Google Scholar] [CrossRef]
- Ammar, E.-D.; Shatters, R.G., Jr.; Hall, D.G. Localization of Candidatus Liberibacter asiaticus, associated with citrus Huanglongbing disease, in its psyllid vector using fluorescence in situ hybridization. J. Phytopathol. 2011, 159, 726–734. [Google Scholar] [CrossRef]
- Ammar, E.-D.; Ramos, J.E.; Hall, D.G.; Dawson, W.O.; Shatters Jr, R.G. Acquisition, replication and inoculation of Candidatus Liberibacter asiaticus following various acquisition periods on Huanglongbing-infected citrus by nymphs and adults of the Asian citrus psyllid. PLoS ONE 2016, 11, e0159594. [Google Scholar] [CrossRef]
- Zheng, Z.; Chen, J.; Deng, X. Historical perspectives, management, and current research of citrus HLB in Guangdong province of China, where the disease has been endemic for over a hundred years. Phytopathology 2018, 108, 1224–1236. [Google Scholar] [CrossRef] [Green Version]
- Beattie, G.A.C. Management of the Asian citrus psyllid in Asia. In Asian Citrus Psyllid: Biology, Ecology and Management of the Huanglongbing Vector; Qureshi, J.A., Stansly, P.A., Eds.; CAB International: Wallingford, UK, 2020; pp. 179–209. [Google Scholar]
- Beattie, G.A.C. Hosts of the Asian citrus psyllid. In Asian Citrus Psyllid: Biology, Ecology and Management of the Huanglongbing Vector; Qureshi, J.A., Stansly, P.A., Eds.; CAB International: Wallingford, UK, 2020; pp. 67–87. [Google Scholar]
- Halbert, S.E.; Manjunath, K.L. Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: A literature review and assessment of risk in Florida. Fla. Entomol. 2004, 87, 330–353. [Google Scholar] [CrossRef]
- Nouri, S.; Salem, N.; Nigg, J.C.; Falk, B.W. Diverse array of new viral sequences identified in worldwide populations of the Asian citrus psyllid (Diaphorina citri) using viral metagenomics. J. Virol. 2016, 90, 2434–2445. [Google Scholar] [CrossRef] [Green Version]
- Hajeri, S.; Killiny, N.; El-Mohtar, C.; Dawson, W.O.; Gowda, S. Citrus tristeza virus-based RNAi in citrus plants induces gene silencingin Diaphorina citri, a phloem-sap sucking insect vector of citrus greening disease (Huanglongbing). J. Biotechnol. 2014, 176, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Tian, M.; Wu, F.; Wang, Y.; Deng, X.; Cen, Y. Transcriptomic analyses of Diaphorina citri Kuwayama in response to the infection by Candidatus Liberibacter asiaticus. Chin. J. Appl. Entomol. 2018, 55, 574–585. [Google Scholar]
- Albiach-Marti, M.R.; Grosser, J.W.; Gowda, S.; Mawassi, M.; Satyanarayana, T.; Garnsey, S.M.; Dawson, W.O. Citrus tristeza virus replicates and forms infectious virions in protoplasts of resistant citrus relatives. Mol. Breed. 2004, 14, 117–128. [Google Scholar] [CrossRef]
- Mourão Filho, F.A.A.; Pio, R.; Mendes, B.M.J.; de Azevedo, F.A.; Schinor, E.H.; Entelmann, F.A.; Alves, A.S.R.; Cantuarias-Avilés, T.E. Evaluation of citrus somatic hybrids for tolerance to Phytophthora nicotianae and Citrus tristeza virus. Sci. Hortic. 2008, 115, 301–308. [Google Scholar] [CrossRef]
- Yoshida, T. Graft compatibility of citrus with plants in the Aurantioideae and their susceptibility to Citrus tristeza virus. Plant Dis. 1996, 80, 414–417. [Google Scholar] [CrossRef]
- Wu, F.; Huang, J.; Xu, M.; Fox, E.G.P.; Beattie, G.A.C.; Holford, P.; Cen, Y.; Deng, X. Host and environmental factors influencing “Candidatus Liberibacter asiaticus” acquisition in Diaphorina citri. Pest Manag. Sci. 2018, 74, 2738–2746. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer 3-new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Ruiz, S.; Moreno, P.; Guerri, J.; Ambrós, S. A real-time RT-PCR assay for detection and absolute quantitation of Citrus tristeza virus in different plant tissues. J. Virol. Meth. 2007, 145, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iracheta, C.M.M.; Arrieta, G.L.C.; Rocha, P.M.A. Detection of the citrus virus by antibodies against the recombinant capsid protein p25 under an immunoimpression system. Mex. J. Plant Pathol. 2012, 30, 31–42. [Google Scholar]
- Wang, C.; Wang, G.; Hong, N.; Jang, B.; Liu, H.; Wu, K. Production of polyclonal and monoclonal antibodies against Citrus tristeza virus and their efficiency for the detection of the virus. Chin. J. Biotechnol. 2006, 22, 629–634. [Google Scholar]
- Dombrovsky, A.; Luria, N. The Nerium oleander aphid Aphis nerii is tolerant to a local isolate of Aphid lethal paralysis virus (ALPV). Virus Genes 2013, 46, 354–361. [Google Scholar] [CrossRef]
- Killiny, N.; Harper, S.J.; Alfaress, S.; El Mohtar, C.; Dawson, W.O. Minor coat and heat-shock proteins are involved in binding of Citrus tristeza virus to the foregut of its aphid vector, Toxoptera citricida. Appl. Environ. Microbiol. 2016, 82, 6294–6302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, W.A.; Lister, C.A. Lime dieback in the Gold Coast, a virus disease of the lime, Citrus Aurantifolia (Christmann) Swingle. J. Hortic. Sci. 1953, 28, 131–140. [Google Scholar] [CrossRef]
- Broadbent, P.; Brlansky, R.H.; Indsto, J. Biological characterization of Australian isolates of Citrus tristeza virus and separation of subisolates by single aphid transmission. Plant Dis. 1996, 80, 329. [Google Scholar] [CrossRef]
- Balaramen, K.; Ramakrishnan, K. Transmission studies with strains of tristeza virus on acid lime. J. Plant Dis. Protec. 1979, 86, 653–661. [Google Scholar]
- Nickel, O.; Klingauf, F.; Fischer, H. Transmission of Citrus tristeza virus (CTV) from “declinamiento”- diseased sweet orange tree on trifoliate orange and its epidemiology to the flight activity of the vector, Toxoptera citricida Kirkc (Homoptera: Aphididae) in Missiones, Argentina. IOCV 1984, 9, 28–32. [Google Scholar]
- Britt, K.; Gebben, S.; Levy, A.; Al Rwahnih, M.; Batuman, O. The detection and surveillance of Asian Citrus Psyllid (Diaphorina citri)—associated viruses in Florida citrus groves. Front. Plant Sci. 2020, 10, 1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boissot, N.; Thomas, S.; Sauvion, N.; Marchal, C.; Pavis, C.; Dogimont, C. Mapping and validation of QTLs for resistance to aphids and whiteflies in melon. Theor. Appl. Genet. 2010, 121, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, L.; Fontana, A.; Sciarroni, R.; Schimio, R.; Lcconsole, G.; Albanese, G.; Saponari, M. Molecular and biological evidence for a severe seedling yellows strain of Citrus tristeza virus spreading in southern Italy. Phytopathol. Mediterr. 2014, 53, 3–13. [Google Scholar]
- Licciardello, G.; Scuderi, G.; Ferraro, R.; Giampetruzzi, A.; Russo, M.; Lombardo, A.; Raspagliesi, D.; Bar-Joseph, M.; Catara, A. Deep sequencing and analysis of small RNAs in sweet orange grafted on sour orange infected with two Citrus tristeza virus isolates prevalent in Sicily. Arch. Virol. 2015, 160, 2583–2589. [Google Scholar] [CrossRef]
- Bar-Joseph, M.; Loebenstein, G. Effects of strain, source plant, and temperature on the transmissibility of Citrus tristeza virus by the melon aphid. Phytopathology 1973, 63, 716–720. [Google Scholar] [CrossRef]
- Fraser, L. Seedling yellows, an unreported virus disease of citrus. Agric. Gaz. NS Wales 1952, 63, 125–131. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, F.; Huang, M.; Fox, E.G.P.; Huang, J.; Cen, Y.; Deng, X.; Xu, M. Preliminary Report on the Acquisition, Persistence, and Potential Transmission of Citrus tristeza virus by Diaphorina citri. Insects 2021, 12, 735. https://doi.org/10.3390/insects12080735
Wu F, Huang M, Fox EGP, Huang J, Cen Y, Deng X, Xu M. Preliminary Report on the Acquisition, Persistence, and Potential Transmission of Citrus tristeza virus by Diaphorina citri. Insects. 2021; 12(8):735. https://doi.org/10.3390/insects12080735
Chicago/Turabian StyleWu, Fengnian, Mochi Huang, Eduardo G. P. Fox, Jiaquan Huang, Yijing Cen, Xiaoling Deng, and Meirong Xu. 2021. "Preliminary Report on the Acquisition, Persistence, and Potential Transmission of Citrus tristeza virus by Diaphorina citri" Insects 12, no. 8: 735. https://doi.org/10.3390/insects12080735
APA StyleWu, F., Huang, M., Fox, E. G. P., Huang, J., Cen, Y., Deng, X., & Xu, M. (2021). Preliminary Report on the Acquisition, Persistence, and Potential Transmission of Citrus tristeza virus by Diaphorina citri. Insects, 12(8), 735. https://doi.org/10.3390/insects12080735