

Effects of a Mealworm (Tenebrio molitor) Extract on Metabolic Syndrome-Related Pathologies: In Vitro Insulin Sensitivity, Inflammatory Response, Hypolipidemic Activity and Oxidative Stress

 , ,
, ,  , , , and
, , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Raw Materials and Chemicals
2.2. Production of the Mealworm Extract by Ultrasound-Assisted Extraction
2.3. Analysis of the Mealworm Extract by Gas Chromatography-Mass Spectrometry (GC-MS)
2.4. Pancreatic Lipase Inhibition Assay
2.5. Effect of the Mealworm Extract on the Intestinal Bioaccessibility of Cholesterol
2.6. Evaluation of the Anti-Inflammatory Activity of Mealworm Extract in LPS-Stimulated Murine Microglial Cells
2.7. Evaluation of the Insulin-Sensitizing Activity of Mealworm Extract in Insulin-Stimulated Primary Mouse Hepatocytes
2.8. Western Blot Analysis
2.9. Evaluation of the Stimulatory Effect on Mitochondrial Respiration of the Mealworm Extract in Primary Bovine Aortic Endothelial Cells (BAECs)
2.10. Evaluation of the Stimulatory Effect on Mitochondrial Superoxide Production of the Mealworm Extract in Primary BAECs
2.11. Antioxidant Activity of the Mealworm Extract by DPPH Assay
2.12. Statistical Analysis
3. Results and Discussion
3.1. Chemical Characterization of the Mealworm Extract
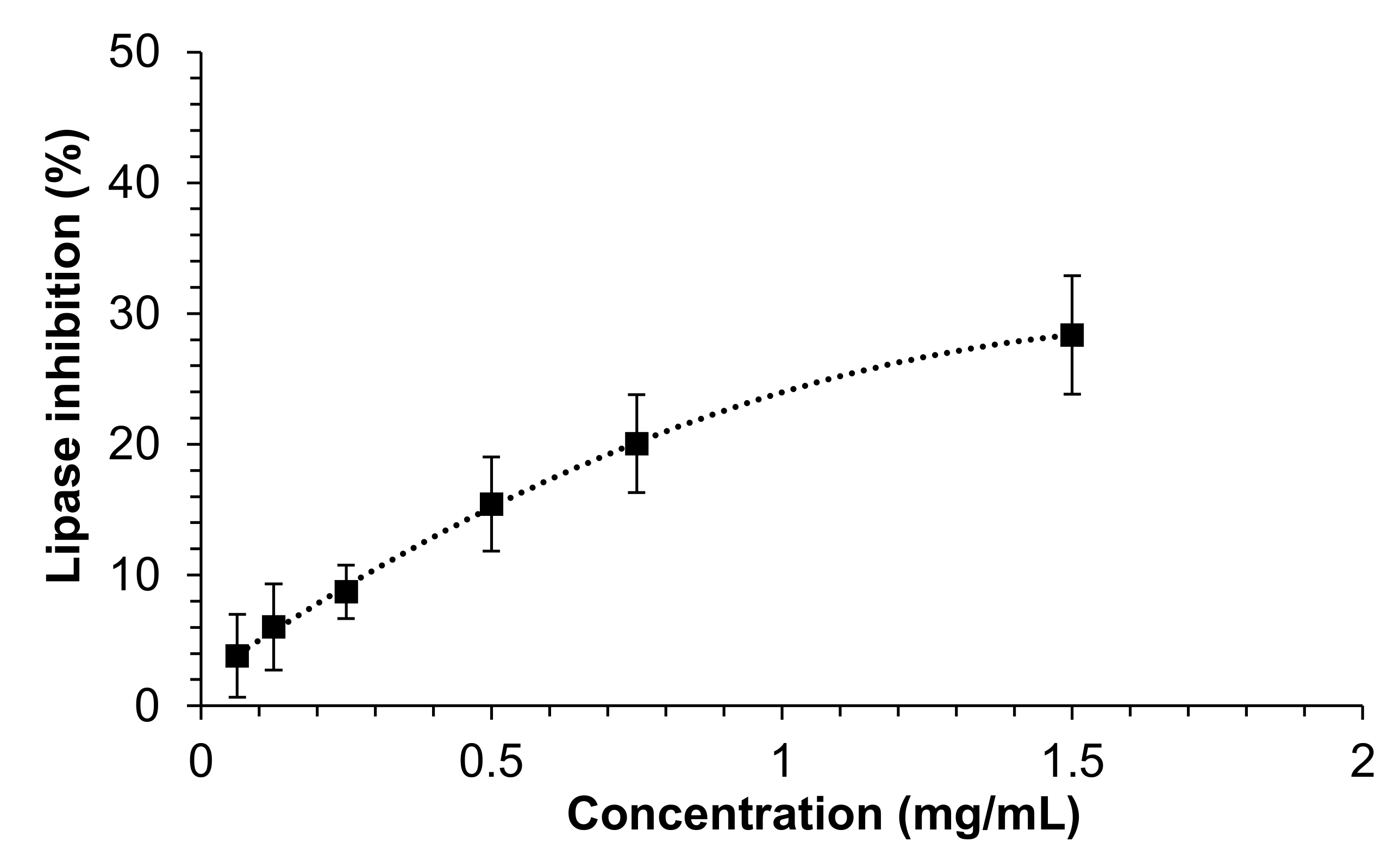
3.2. Inhibitory Activity against Pancreatic Lipase of Mealworm Extract
3.3. Effect of Mealworm Extract on the Bioaccessibility of Cholesterol
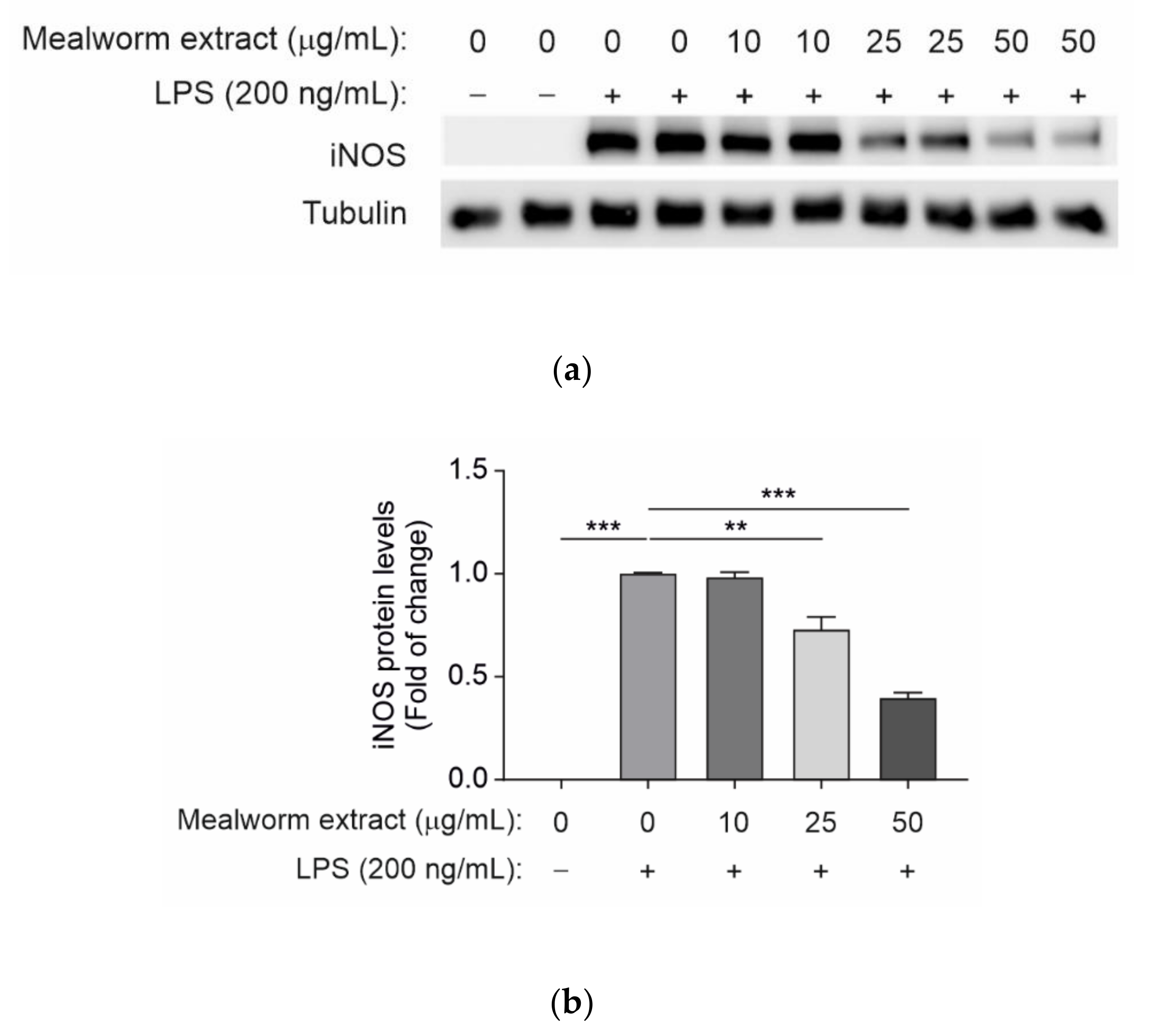
3.4. Effect of Mealworm Extract in Attenuating the Inflammatory Response Induced by LPS in Microglial Cells
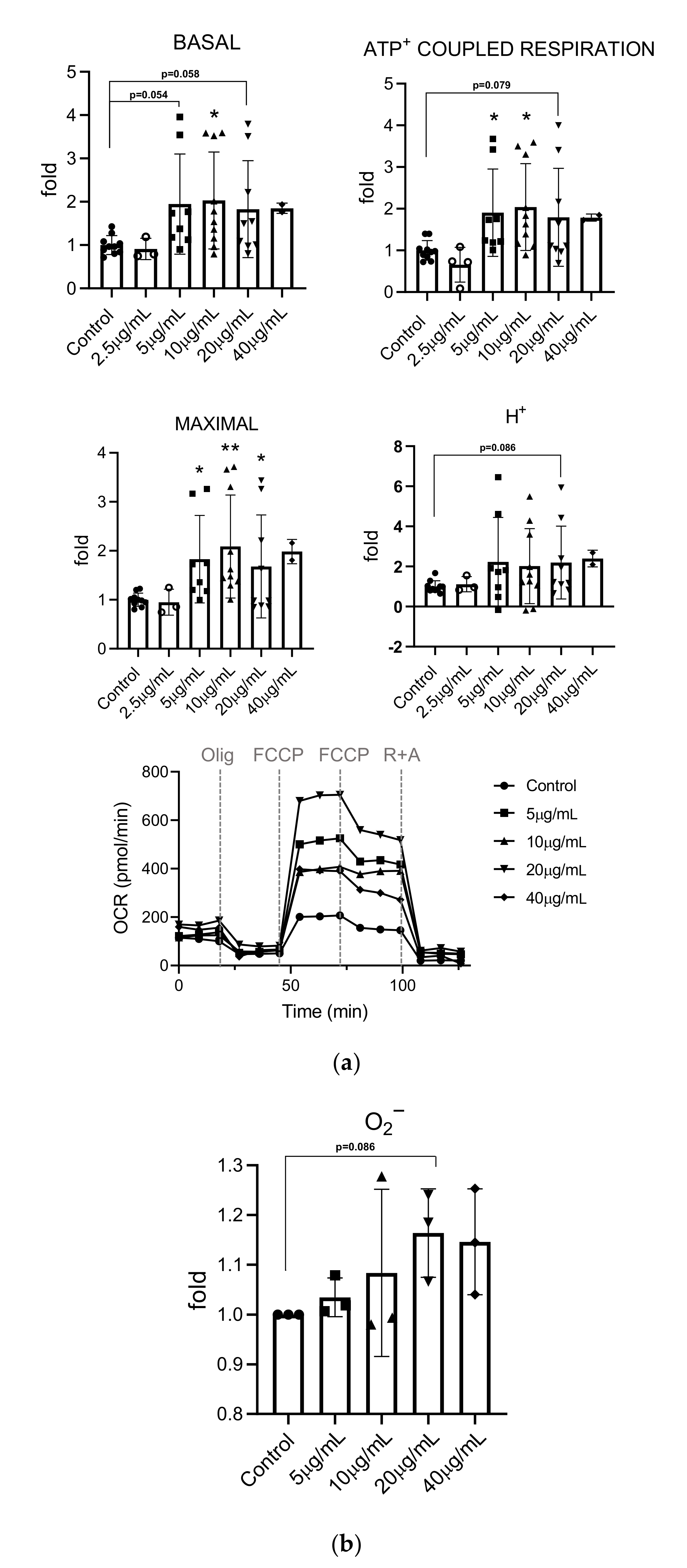
3.5. Effects of Mealworm Extract on Mitochondrial Respiration and Superoxide Production
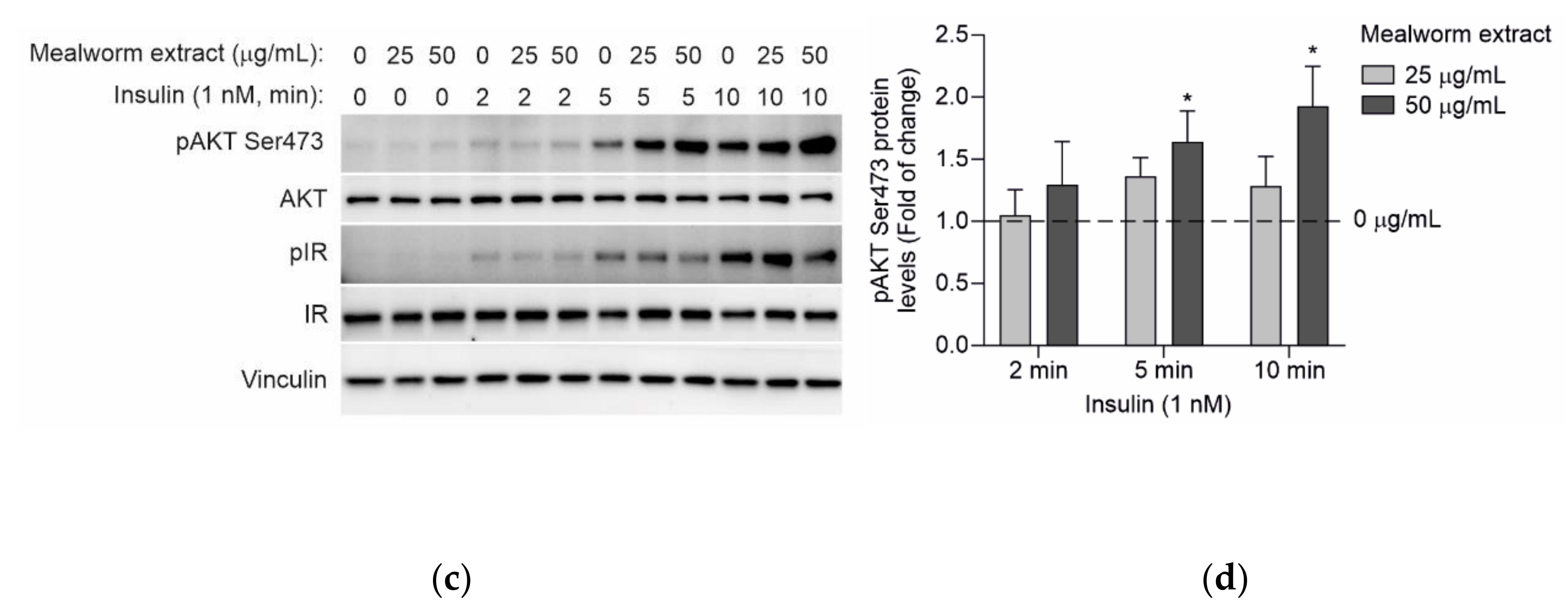
3.6. Effect of the Mealworm Extract in the Early Steps of the Insulin Signaling Cascade in Primary Mouse Hepatocytes
3.7. Antioxidant Activity of the Mealworm Extract
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scuteri, A.; Laurent, S.; Cucca, F.; Cockcroft, J.; Cunha, P.G.; Mañas, L.R.; Raso, F.U.M.; Muiesan, M.L.; Ryliskyte, L.; Rietzschel, E.; et al. Metabolic syndrome across Europe: Different clusters of risk factors. Eur. J. Prev. Cardiol. 2015, 22, 486–491. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Ryu, B.; Ahn, G.; Yeo, I.-K.; Jeon, Y.-J. Therapeutic potential of algal natural products against metabolic syndrome: A review of recent developments. Trends Food Sci. Technol. 2020, 97, 286–299. [Google Scholar] [CrossRef]
- Tung, Y.-T.; Pan, C.-H.; Chien, Y.-W.; Huang, H.-Y. Edible mushrooms: Novel medicinal agents to combat metabolic syndrome and associated diseases. Curr. Pharm. Des. 2020, 26, 4970–4981. [Google Scholar] [CrossRef]
- Francini, F.-P.; Spinella, P.; Calò, L.A. Potential role of phytochemicals in metabolic syndrome prevention and therapy. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 1987–2002. [Google Scholar] [CrossRef]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013; ISBN 9789251075968. [Google Scholar]
- Van Huis, A. Insects as food and feed, a new emerging agricultural sector: A review. J. Insects Food Feed 2020, 6, 27–44. [Google Scholar] [CrossRef]
- Van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef]
- European Parliament and Council of the European Union Regulation (EU) 2015/2283 of the European Parliament and of the Council of 25 November 2015 on novel foods, amending Regulation (EU) No 1169/2011 of the European Parliament and of the Council and repealing Regulation (EC) No 258/97 of the European Parliam. Off. J. Eur. Union 2015, L 327, 1–22.
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Pelaez, C.; et al. Safety of dried yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06343. [Google Scholar] [CrossRef] [PubMed]
- Navarro del Hierro, J.; Hernández-Ledesma, B.; Martin, D. Potential of edible insects as a new source of bioactive compounds against metabolic syndrome. In Current Advances for Development of Functional Foods Modulating Inflammation and Oxidative Stress; Hernández-Ledesma, B., Martínez-Villaluenga, C.B.T.-C.A., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 331–364. ISBN 978-0-12-823482-2. [Google Scholar]
- D’Antonio, V.; Serafini, M.; Battista, N. Dietary modulation of oxidative stress from edible insects: A mini-review. Front. Nutr. 2021, 8, 642551. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Lucas, A.J.; Menegon de Oliveira, L.; da Rocha, M.; Prentice, C. Edible insects: An alternative of nutritional, functional and bioactive compounds. Food Chem. 2020, 311, 126022. [Google Scholar] [CrossRef]
- Dutta, P.; Sahu, R.K.; Dey, T.; Lahkar, M.D.; Manna, P.; Kalita, J. Beneficial role of insect-derived bioactive components against inflammation and its associated complications (colitis and arthritis) and cancer. Chem. Biol. Interact. 2019, 313, 108824. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. Unlocking the biological potential of proteins from edible insects through enzymatic hydrolysis: A review. Innov. Food Sci. Emerg. Technol. 2017, 43, 239–252. [Google Scholar] [CrossRef]
- Nowakowski, A.C.; Miller, A.C.; Miller, M.E.; Xiao, H.; Wu, X. Potential health benefits of edible insects. Crit. Rev. Food Sci. Nutr. 2021, 62, 3499–3508. [Google Scholar] [CrossRef]
- Navarro del Hierro, J.; Gutiérrez-Docio, A.; Otero, P.; Reglero, G.; Martin, D. Characterization, antioxidant activity, and inhibitory effect on pancreatic lipase of extracts from the edible insects Acheta domesticus and Tenebrio molitor. Food Chem. 2020, 309, 125742. [Google Scholar] [CrossRef]
- Navarro del Hierro, J.; Cantero-Bahillo, E.; Fornari, T.; Martin, D. Effect of defatting and extraction solvent on the antioxidant and pancreatic lipase inhibitory activities of extracts from Hermetia illucens and Tenebrio molitor. Insects 2021, 12, 789. [Google Scholar] [CrossRef]
- Cantero-Bahillo, E.; Navarro del Hierro, J.; Hernández, D.M.; Fernández-Felipe, M.T.; Fornari, T.; Martin, D. Supercritical-CO2 for defatting and production of bioactive extracts from black soldier fly (Hermetia illucens) larvae. J. Insects Food Feed. 2022. In press. [Google Scholar] [CrossRef]
- Ham, J.R.; Choi, R.Y.; Lee, Y.; Lee, M.K. Effects of edible insect Tenebrio molitor larva fermentation extract as a substitute protein on hepatosteatogenesis and proteomic changes in obese mice induced by high-fat diet. Int. J. Mol. Sci. 2021, 22, 3615. [Google Scholar] [CrossRef] [PubMed]
- Herrera, T.; Navarro del Hierro, J.; Fornari, T.; Reglero, G.; Martin, D. Inhibitory effect of quinoa and fenugreek extracts on pancreatic lipase and α-amylase under in vitro traditional conditions or intestinal simulated conditions. Food Chem. 2019, 270, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Navarro del Hierro, J.; Casado-Hidalgo, G.; Reglero, G.; Martin, D. The hydrolysis of saponin-rich extracts from fenugreek and quinoa improves their pancreatic lipase inhibitory activity and hypocholesterolemic effect. Food Chem. 2021, 338, 128113. [Google Scholar] [CrossRef]
- Kolarič, L.; Šimko, P. The comparison of HPLC and spectrophotometric method for cholesterol determination. Potravin. Slovak J. Food Sci. 2020, 14, 118–124. [Google Scholar] [CrossRef]
- Benveniste, R.; Danoff, T.M.; Ilekis, J.; Craig, H.R. Epidermal growth factor receptor numbers in male and female mouse primary hepatocyte cultures. Cell Biochem. Funct. 1988, 6, 231–235. [Google Scholar] [CrossRef]
- Blois, M. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Mazzocchi, A.; De Cosmi, V.; Risé, P.; Milani, G.P.; Turolo, S.; Syrén, M.L.; Sala, A.; Agostoni, C. Bioactive compounds in edible oils and their role in oxidative stress and inflammation. Front. Physiol. 2021, 12, 598. [Google Scholar] [CrossRef]
- Marangoni, F.; Agostoni, C.; Borghi, C.; Catapano, A.L.; Cena, H.; Ghiselli, A.; La Vecchia, C.; Lercker, G.; Manzato, E.; Pirillo, A.; et al. Dietary linoleic acid and human health: Focus on cardiovascular and cardiometabolic effects. Atherosclerosis 2020, 292, 90–98. [Google Scholar] [CrossRef]
- Krishnan, N.; Dickman, M.B.; Becker, D.F. Proline modulates the intracellular redox environment and protects mammalian cells against oxidative stress. Free Radic. Biol. Med. 2008, 44, 671–681. [Google Scholar] [CrossRef]
- Obayashi, Y.; Arisaka, H.; Yoshida, S.; Mori, M.; Takahashi, M. The protection mechanism of proline from D-galactosamine hepatitis involves the early activation of ROS-eliminating pathway in the liver. Springerplus 2015, 4, 199. [Google Scholar] [CrossRef] [PubMed]
- Andrade, V.S.; Rojas, D.B.; de Andrade, R.B.; Kim, T.D.H.; Vizuete, A.F.; Zanatta, Â.; Wajner, M.; Gonçalves, C.A.S.; Wannmacher, C.M.D. A possible anti-inflammatory effect of proline in the brain cortex and cerebellum of rats. Mol. Neurobiol. 2018, 55, 4068–4077. [Google Scholar] [CrossRef] [PubMed]
- Kosmadaki, M.; Katsambas, A. Topical treatments for acne. Clin. Dermatol. 2017, 35, 173–178. [Google Scholar] [CrossRef]
- Kumar, A.; Rao, R.; Yadav, P. Azelaic acid: A promising agent for dermatological applications. Curr. Drug Ther. 2019, 15, 181–193. [Google Scholar] [CrossRef]
- Kamzolova, S.V.; Samoilenko, V.A.; Lunina, J.N.; Morgunov, I.G. Isocitric acid production from ethanol industry waste by Yarrowia lipolytica. Fermentation 2021, 7, 146. [Google Scholar] [CrossRef]
- Lunagariya, N.A.; Patel, N.K.; Jagtap, S.C.; Bhutani, K.K. Inhibitors of pancreatic lipase: State of the art and clinical perspectives. EXCLI J. 2014, 13, 897. [Google Scholar] [CrossRef] [PubMed]
- Patch, C.S.; Tapsell, L.C.; Williams, P.G.; Gordon, M. Plant sterols as dietary adjuvants in the reduction of cardiovascular risk: Theory and evidence. Vasc. Health Risk Manag. 2006, 2, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, Y.; Pastrana, I.; Rebollo-Hernanz, M.; Benitez, V.; Álvarez-Rivera, G.; Viejo, J.L.; Martín-Cabrejas, M.A. Investigating edible insects as a sustainable food source: Nutritional value and techno-functional and physiological properties. Food Funct. 2021, 12, 6309–6322. [Google Scholar] [CrossRef] [PubMed]
- Zadeh, Z.S.; Kheiri, F.; Faghani, M. Use of yellow mealworm (Tenebrio molitor) as a protein source on growth performance, carcass traits, meat quality and intestinal morphology of Japanese quails (Coturnix japonica). Vet. Anim. Sci. 2019, 8, 100066. [Google Scholar] [CrossRef]
- Sarica, S.; Kanoglu, B.; Yildirim, U. Defatted yellow mealworm larvae (Tenebrio molitor L.) meal as possible alternative to fish meal in quail diets. S. Afr. J. Anim. Sci. 2020, 50, 481–491. [Google Scholar] [CrossRef]
- Sankian, Z.; Khosravi, S.; Kim, Y.-O.; Lee, S.-M. Effects of dietary inclusion of yellow mealworm (Tenebrio molitor) meal on growth performance, feed utilization, body composition, plasma biochemical indices, selected immune parameters and antioxidant enzyme activities of mandarin fish (Siniperca scherze). Aquaculture 2018, 496, 79–87. [Google Scholar] [CrossRef]
- AL-Dalaeen, A.; AL-Domi, H. Does obesity put your brain at risk? Diabetes Metab. Syndr. Clin. Res. Rev. 2022, 16, 102444. [Google Scholar] [CrossRef] [PubMed]
- Mou, Y.; Du, Y.; Zhou, L.; Yue, J.; Hu, X.; Liu, Y.; Chen, S.; Lin, X.; Zhang, G.; Xiao, H.; et al. Gut microbiota interact with the brain through systemic chronic inflammation: Implications on neuroinflammation, neurodegeneration, and aging. Front. Immunol. 2022, 13, 1046. [Google Scholar] [CrossRef]
- Yu, J.-M.; Jang, J.-Y.; Kim, H.-J.; Cho, Y.-H.; Kim, D.; Kwon, O.; Cho, Y.-J.; An, B.-J. Antioxidant capacity and Raw 264.7 macrophage anti-inflammatory effect of the Tenebrio Molitor. Korean J. Food Preserv. 2016, 23, 890–898. [Google Scholar] [CrossRef]
- Seo, M.; Lee, H.J.; Lee, H.; Baek, M.; Kim, I.-W.; Kim, S.Y.; Hwang, J.-S.; Kim, M.-A. A study of the anti-inflammatory effect of protein derived from Tenebrio molitor larvae. J. Life Sci. 2019, 29, 854–860. [Google Scholar] [CrossRef]
- Fabregat-Andrés, Ó.; Tierrez, A.; Mata, M.; Estornell-Erill, J.; Ridocci-Soriano, F.; Monsalve, M. Induction of PGC-1α expression can be detected in blood samples of patients with ST-segment elevation acute myocardial infarction. PLoS ONE 2011, 6, e26913. [Google Scholar] [CrossRef] [PubMed]
- White, M.F.; Kahn, C.R. Insulin action at a molecular level–100 years of progress. Mol. Metab. 2021, 52, 101304. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Park, J.E.; Han, J.S. Tenebrio molitor (mealworm) extract improves insulin sensitivity and alleviates hyperglycemia in C57BL/Ksj-db/db mice. J. Life Sci. 2019, 29, 570–579. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rt (min) | Compound | Area | % |
|---|---|---|---|
| LIPIDS | |||
| Fatty acids | |||
| 12.485 | Dodecanoic acid | 17,480 | 0.20 |
| 13.87 | Tetradecanoic acid | 78,440 | 0.88 |
| 15.02 | Fatty acid n.i. | 49,241 | 0.55 |
| 15.139 | Hexadecanoic acid | 438,621 | 4.93 |
| 16.143 | 9,12-Octadecadienoic acid | 668,487 | 7.52 |
| 16.169 | (9Z)-Octadecenoic acid | 589,525 | 6.63 |
| 16.309 | Octadecanoic acid | 236,122 | 2.66 |
| Monoglycerides | |||
| 17.204 | 1-Monomyristin | 41,992 | 0.47 |
| 18.029 | 2-Monopalmitin | 143,652 | 1.62 |
| 18.202 | 1-Monopalmitin | 1,113,769 | 12.53 |
| 18.958 | 2-Monostearin | 106,180 | 1.19 |
| 19.131 | Monoglyceride n.i. + sugar n.i. | 941,077 | 10.59 |
| NITROGEN COMPOUNDS | |||
| Amino acids and derivatives | |||
| 8.908 | Valine + amino acid n.i. | 115,807 | 1.30 |
| 9.424 | Leucine | 55,652 | 0.63 |
| 9.625 | Isoleucine | 72,854 | 0.82 |
| 9.651 | Proline | 737,659 | 8.30 |
| 10.228 | Serine | 36,408 | 0.41 |
| 10.468 | Threonine | 30,762 | 0.35 |
| 10.719 | Aspartic acid | 12,740 | 0.14 |
| 11.55 | Pyroglutamic acid | 294,332 | 3.31 |
| 11.588 | Amino acid n.i. | 54,628 | 0.61 |
| 12.314 | Amino acid n.i. | 18,931 | 0.21 |
| 12.371 | Amino acid n.i. | 8630 | 0.10 |
| 14.569 | Tyrosine | 289,818 | 3.26 |
| 16.199 | Tryptophan | 145,443 | 1.64 |
| Non-protein nitrogen compounds | |||
| 9.112 | Urea | 33,290 | 0.37 |
| 15.637 | Uric acid | 35,977 | 0.40 |
| ACIDS | |||
| 9.461 | Phosphoric acid | 1,115,488 | 12.55 |
| 9.758 | Butanedioic acid | 21,627 | 0.24 |
| 13.42 | Glycerophosphoric acid | 83,584 | 0.94 |
| 13.537 | Nonanedioic acid | 59,739 | 0.67 |
| 13.811 | Isocitric acid | 158,515 | 1.78 |
| CARBOHYDRATES | |||
| 18.263 | Disaccharide n.i. | 297,470 | 3.35 |
| 18.483 | Disaccharide n.i. | 191,649 | 2.16 |
| 18.522 | Disaccharide n.i. | 180,864 | 2.03 |
| 18.672 | Disaccharide n.i. | 45,385 | 0.51 |
| 19.012 | Sugar n.i. | 92,244 | 1.04 |
| 19.209 | Sugar n.i. | 129,436 | 1.46 |
| STEROLS | |||
| 20.897 | Cholesterol | 108,886 | 1.22 |
| HYDROCARBONS | |||
| 17.692 | Alcane n.i. | 37,742 | 0.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro del Hierro, J.; Cantero-Bahillo, E.; Fernández-Felipe, M.T.; García-Risco, M.R.; Fornari, T.; Rada, P.; Doblado, L.; Ferreira, V.; Hitos, A.B.; Valverde, Á.M.; et al. Effects of a Mealworm (Tenebrio molitor) Extract on Metabolic Syndrome-Related Pathologies: In Vitro Insulin Sensitivity, Inflammatory Response, Hypolipidemic Activity and Oxidative Stress. Insects 2022, 13, 896. https://doi.org/10.3390/insects13100896
Navarro del Hierro J, Cantero-Bahillo E, Fernández-Felipe MT, García-Risco MR, Fornari T, Rada P, Doblado L, Ferreira V, Hitos AB, Valverde ÁM, et al. Effects of a Mealworm (Tenebrio molitor) Extract on Metabolic Syndrome-Related Pathologies: In Vitro Insulin Sensitivity, Inflammatory Response, Hypolipidemic Activity and Oxidative Stress. Insects. 2022; 13(10):896. https://doi.org/10.3390/insects13100896
Chicago/Turabian StyleNavarro del Hierro, Joaquín, Emma Cantero-Bahillo, M. Teresa Fernández-Felipe, Mónica R. García-Risco, Tiziana Fornari, Patricia Rada, Laura Doblado, Vitor Ferreira, Ana B. Hitos, Ángela M. Valverde, and et al. 2022. "Effects of a Mealworm (Tenebrio molitor) Extract on Metabolic Syndrome-Related Pathologies: In Vitro Insulin Sensitivity, Inflammatory Response, Hypolipidemic Activity and Oxidative Stress" Insects 13, no. 10: 896. https://doi.org/10.3390/insects13100896
APA StyleNavarro del Hierro, J., Cantero-Bahillo, E., Fernández-Felipe, M. T., García-Risco, M. R., Fornari, T., Rada, P., Doblado, L., Ferreira, V., Hitos, A. B., Valverde, Á. M., Monsalve, M., & Martin, D. (2022). Effects of a Mealworm (Tenebrio molitor) Extract on Metabolic Syndrome-Related Pathologies: In Vitro Insulin Sensitivity, Inflammatory Response, Hypolipidemic Activity and Oxidative Stress. Insects, 13(10), 896. https://doi.org/10.3390/insects13100896