
Sympatric Populations of the Anopheles gambiae Complex in Southwest Burkina Faso Evolve Multiple Diverse Resistance Mechanisms in Response to Intense Selection Pressure with Pyrethroids

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
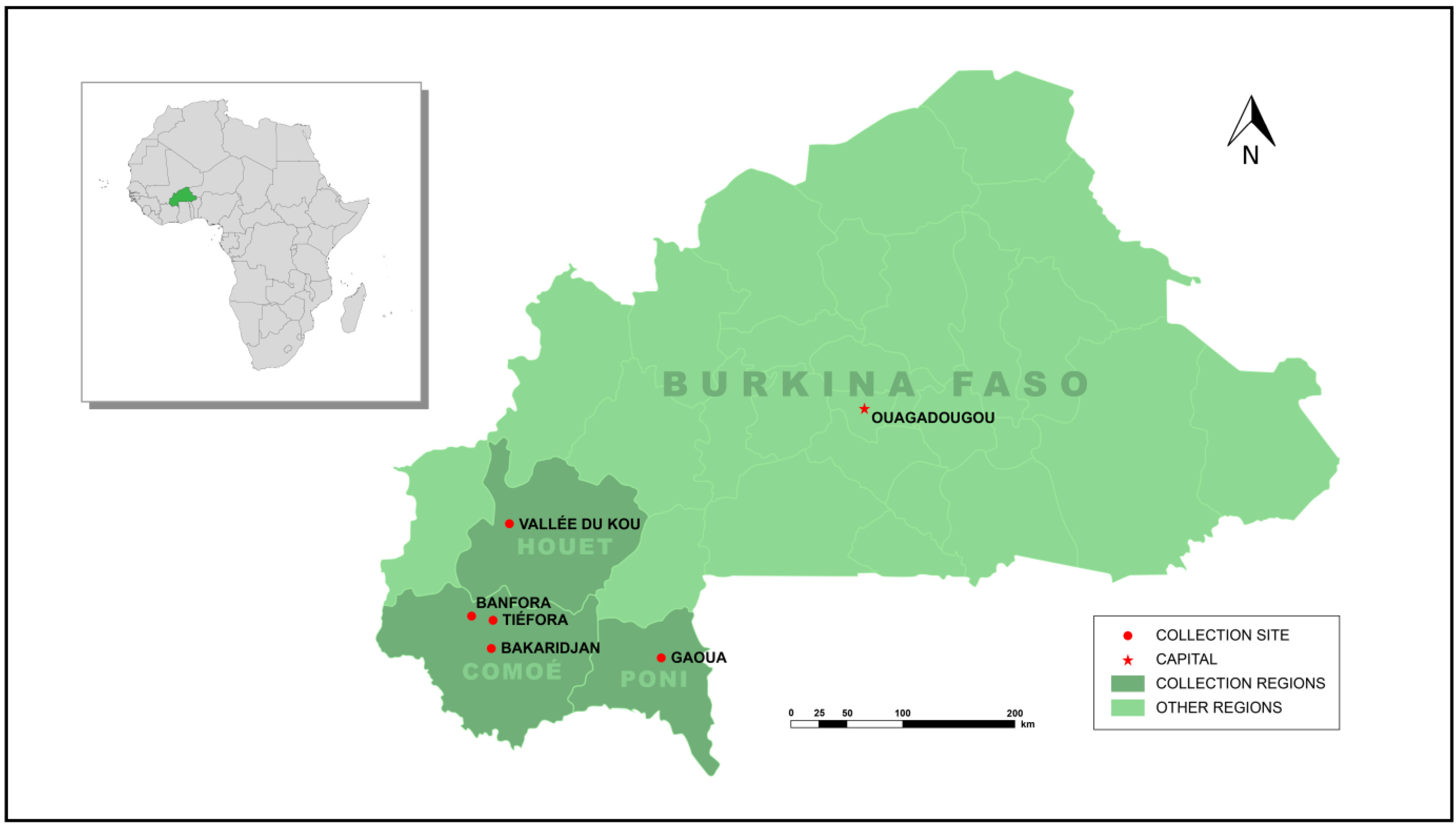
2.1. Establishment of Strains
2.2. Mosquito Rearing
2.3. Selection and Resistance Profiling
2.4. Synergist Bioassays
2.5. Cone Bioassays
2.6. Target Site Mutation Genotyping
2.7. RNAseq Transcriptomic Analysis
2.8. Metabolic Resistance—Detox Gene Expression Levels
3. Results
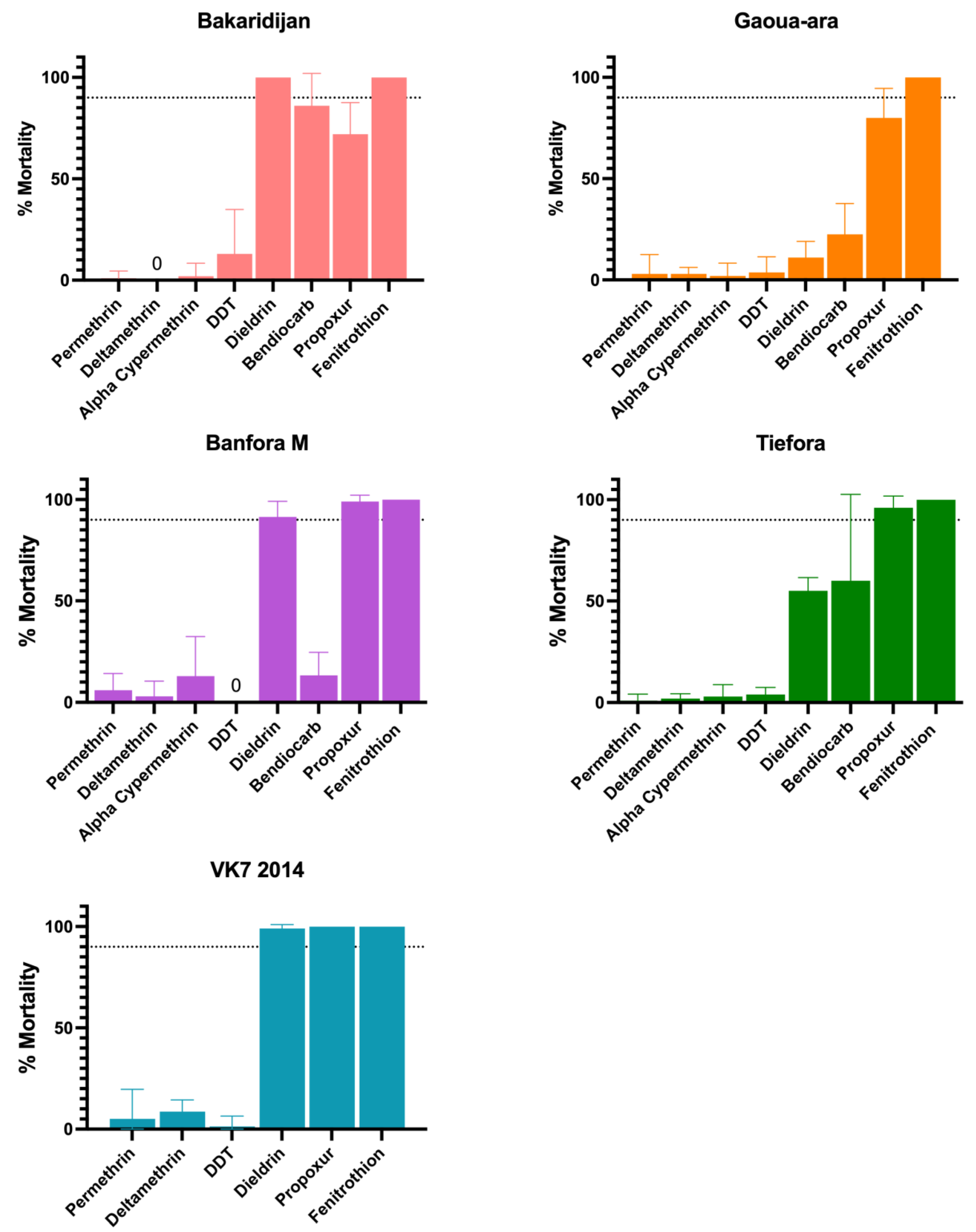
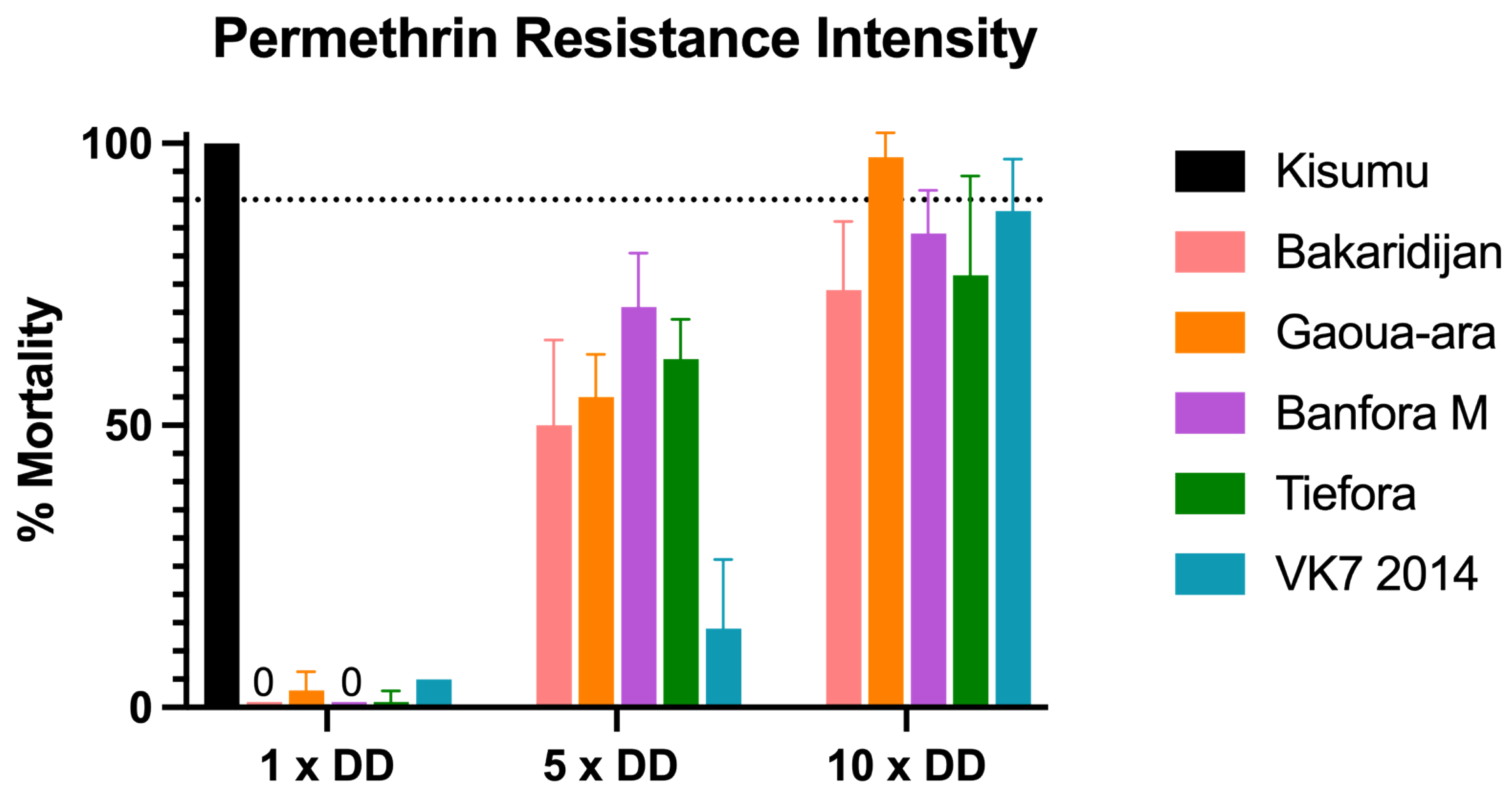
3.1. Discriminating Dose Assays
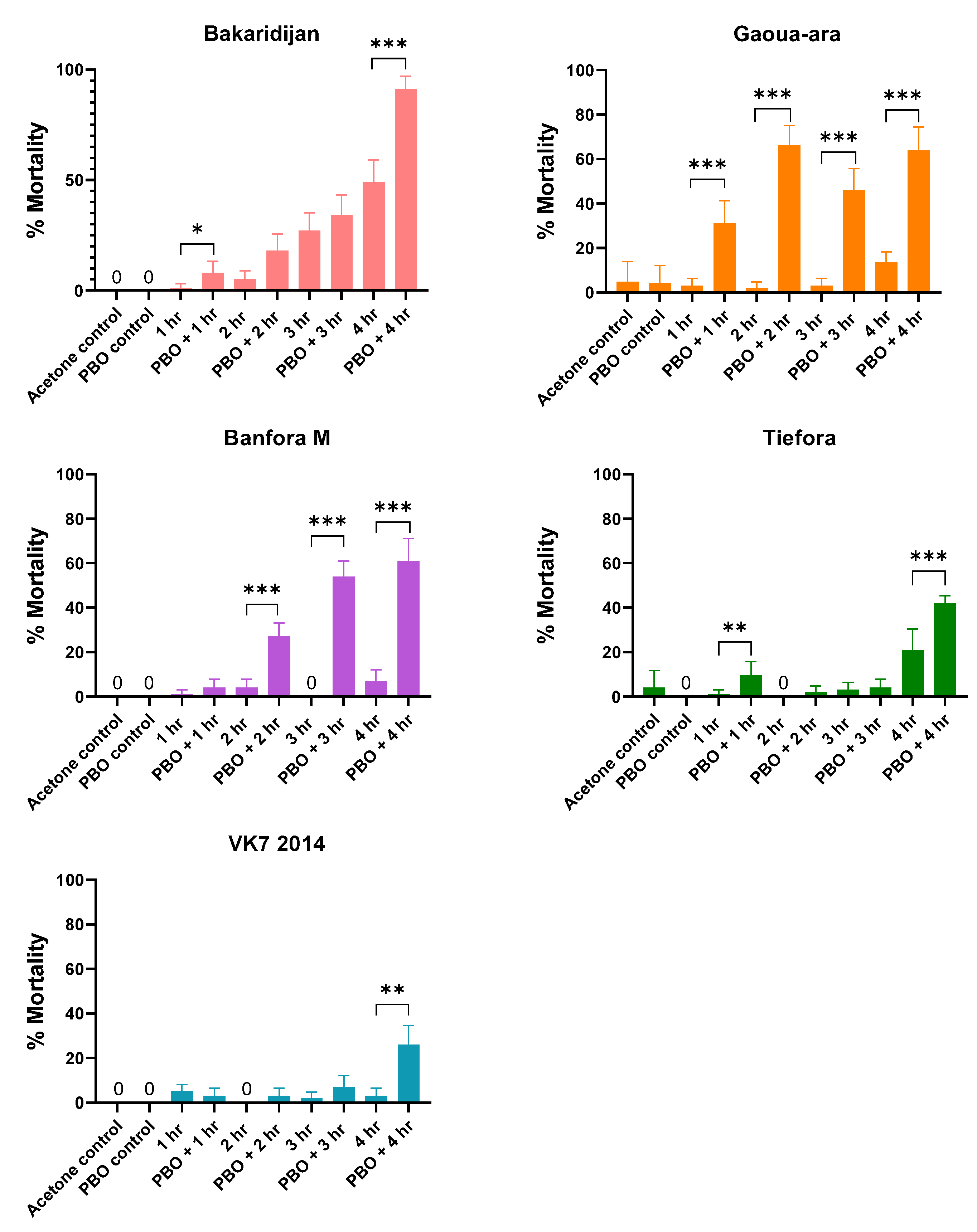
3.2. Impact of PBO on Pyrethroid Mortality
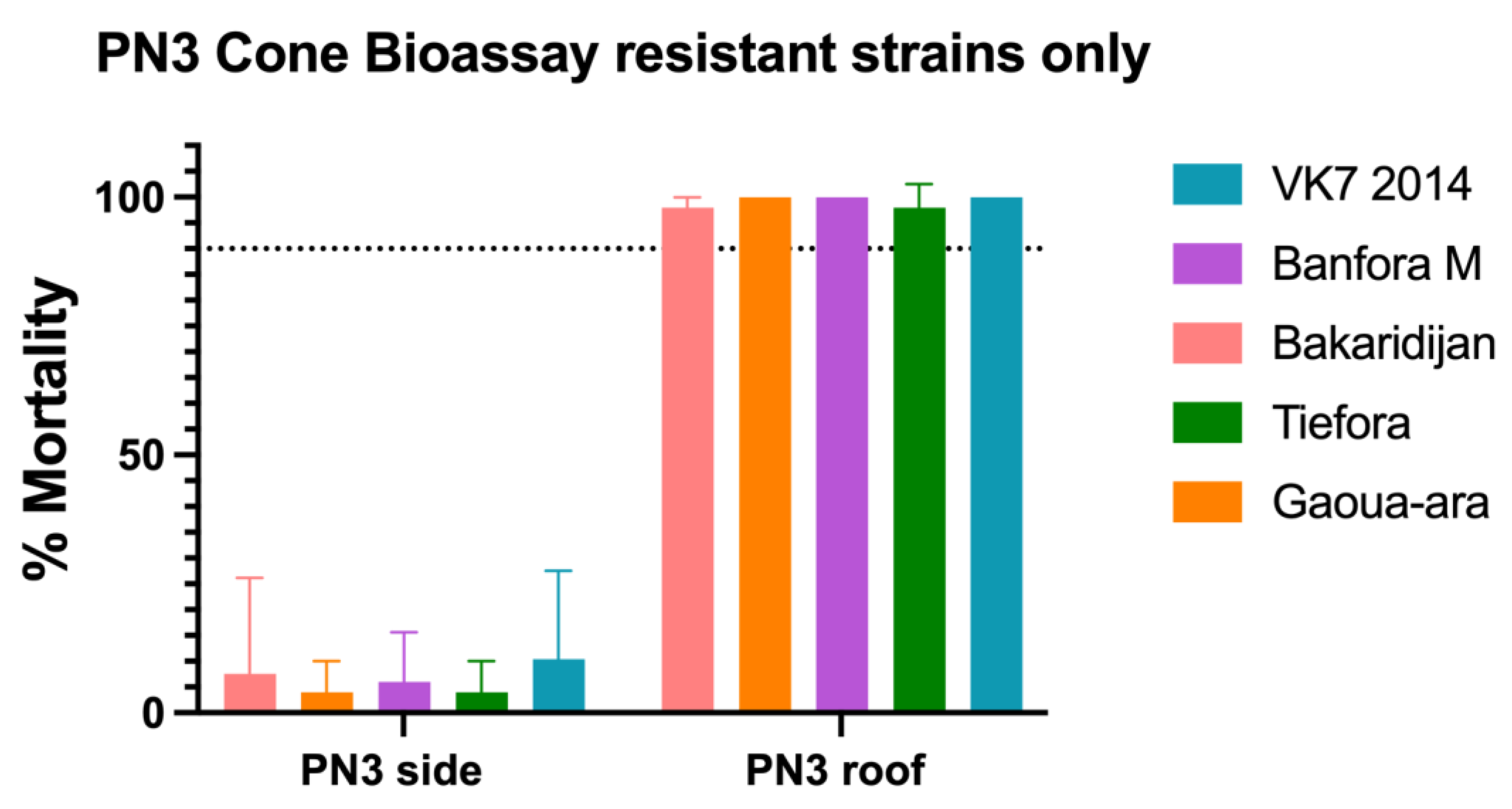
3.3. Cone Bioassays
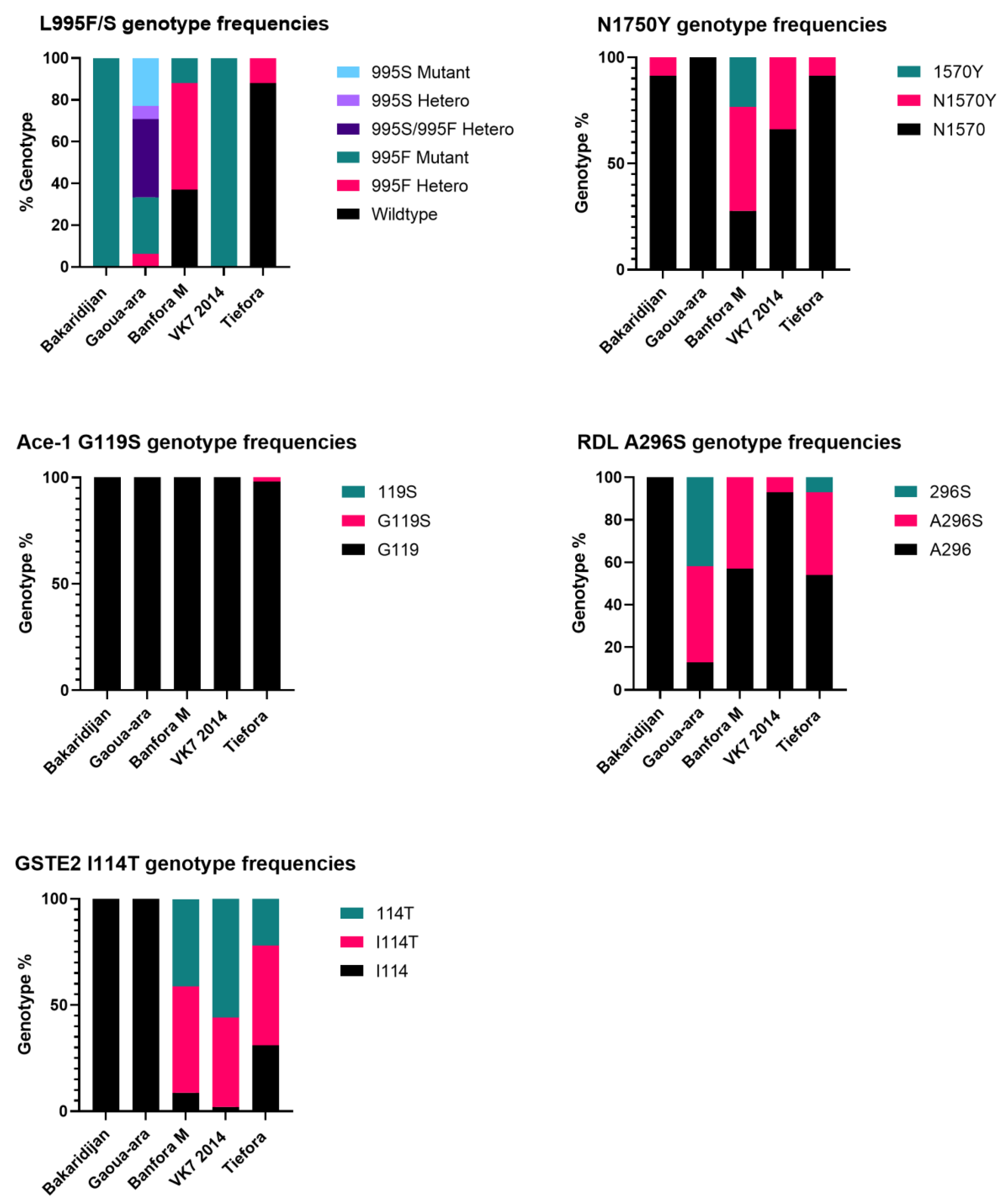
3.4. Target Site Mutation Genotyping
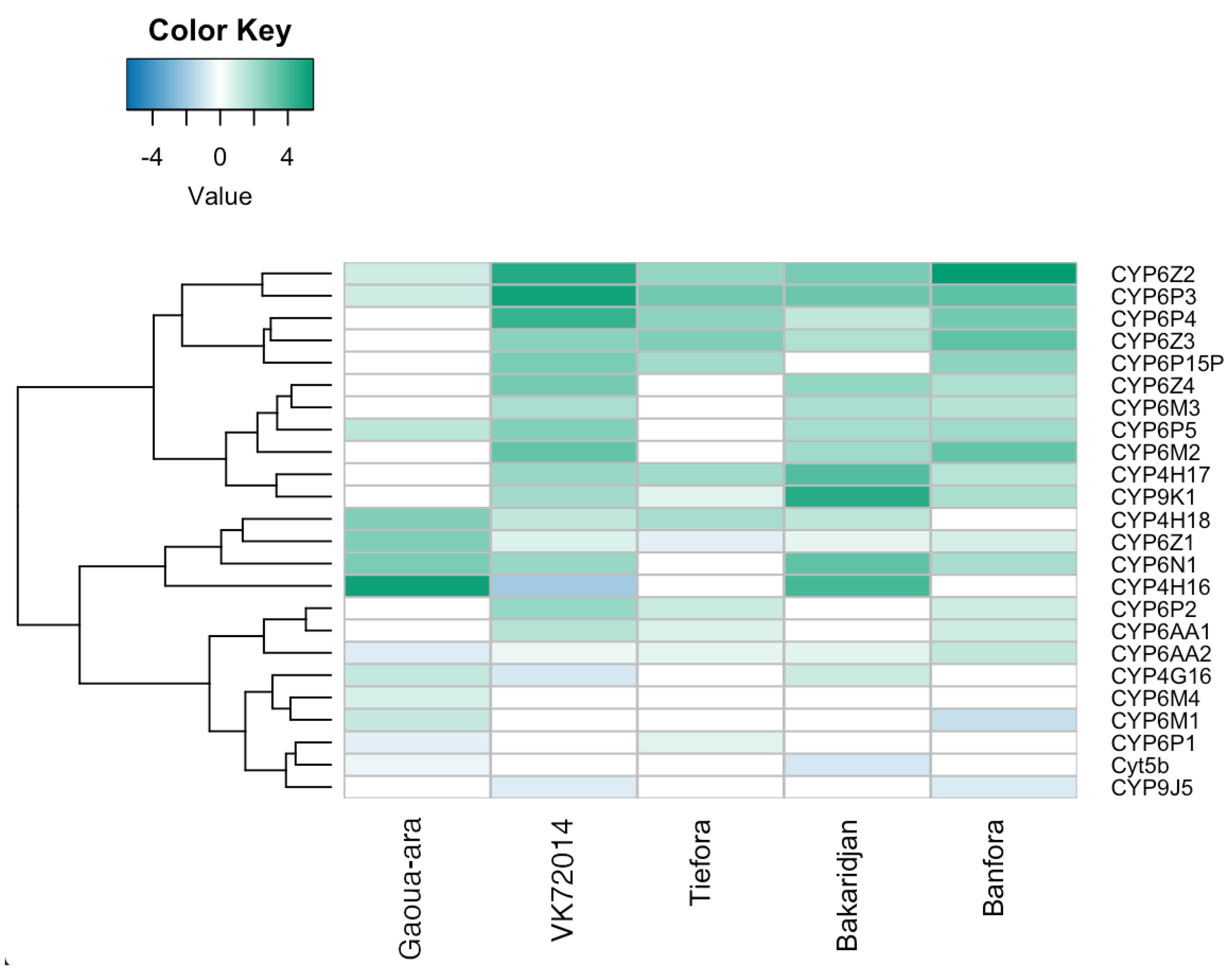
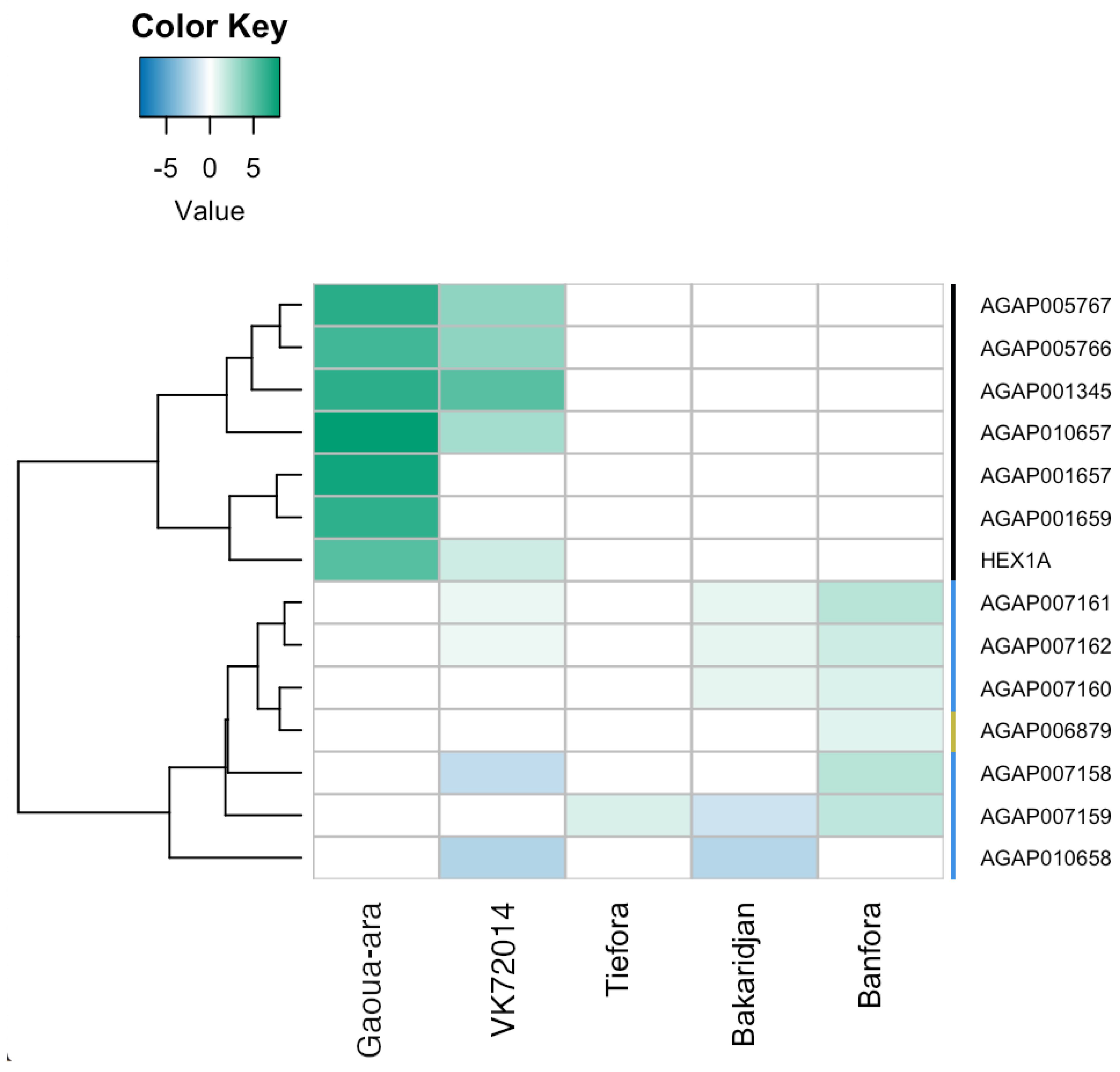
3.5. RNAseq Analysis
3.6. Similarities between Strains
3.7. Differences between Strains
3.8. Evaluation of a Multiplex Gene Expression Assay for Metabolic Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oxborough, R.M. Trends in US President’s Malaria Initiative-funded indoor residual spray coverage and insecticide choice in sub-Saharan Africa (2008–2015): Urgent need for affordable, long-lasting insecticides. Malar. J. 2016, 15, 146. [Google Scholar] [CrossRef] [Green Version]
- Nkya, T.E.; Akhouayri, I.; Kisinza, W.; David, J.P. Impact of environment on mosquito response to pyrethroid insecticides: Facts, evidences and prospects. Insect Biochem. Mol. Biol. 2013, 43, 407–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hien, A.S.; Soma, D.D.; Hema, O.; Bayili, B.; Namountougou, M.; Gnankiné, O.; Baldet, T.; Diabaté, A.; Dabiré, K.R. Evidence that agricultural use of pesticides selects pyrethroid resistance within Anopheles gambiae s.l. populations from cotton growing areas in Burkina Faso, West Africa. PLoS ONE 2017, 12, e0173098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.W. Insecticide resistance in mosquitoes: A pragmatic review. J. Am. Mosq. Control Assoc. 1986, 2, 123–140. [Google Scholar] [PubMed]
- Ranson, H.; Lissenden, N. Insecticide Resistance in African Anopheles Mosquitoes: A Worsening Situation that Needs Urgent Action to Maintain Malaria Control. Trends Parasitol. 2016, 32, 187–196. [Google Scholar] [CrossRef]
- Lees, R.; Praulins, G.; Davies, R.; Brown, F.; Parsons, G.; White, A.; Ranson, H.; Small, G.; Malone, D. A testing cascade to identify repurposed insecticides for next-generation vector control tools: Screening a panel of chemistries with novel modes of action against a malaria vector. Gates Open Res. 2019, 3, 1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J.A.; Ruscoe, C.N.; Perrior, T.R. Discovery to Development: Insecticides for Malaria Vector Control. Chimia 2016, 70, 684–693. [Google Scholar] [CrossRef]
- Lees, R.S.; Ismail, H.M.; Logan, R.A.E.; Malone, D.; Davies, R.; Anthousi, A.; Adolfi, A.; Lycett, G.J.; Paine, M.J.I. New insecticide screening platforms indicate that Mitochondrial Complex I inhibitors are susceptible to cross-resistance by mosquito P450s that metabolise pyrethroids. Sci. Rep. 2020, 10, 16232. [Google Scholar] [CrossRef]
- Williams, J.; Flood, L.; Praulins, G.; Ingham, V.A.; Morgan, J.; Lees, R.S.; Ranson, H. Characterisation of Anopheles strains used for laboratory screening of new vector control products. Parasites Vectors 2019, 12, 522. [Google Scholar] [CrossRef] [Green Version]
- Adams, K.L.; Sawadogo, S.P.; Nignan, C.; Niang, A.; Paton, D.G.; Robert Shaw, W.; South, A.; Wang, J.; Itoe, M.A.; Werling, K.; et al. Cuticular hydrocarbons are associated with mating success and insecticide resistance in malaria vectors. Commun. Biol. 2021, 4, 911. [Google Scholar] [CrossRef]
- Balabanidou, V.; Kampouraki, A.; MacLean, M.; Blomquist, G.J.; Tittiger, C.; Juárez, M.P.; Mijailovsky, S.J.; Chalepakis, G.; Anthousi, A.; Lynd, A.; et al. Cytochrome P450 associated with insecticide resistance catalyzes cuticular hydrocarbon production in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2016, 113, 9268–9273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingham, V.A.; Anthousi, A.; Douris, V.; Harding, N.J.; Lycett, G.; Morris, M.; Vontas, J.; Ranson, H. A sensory appendage protein protects malaria vectors from pyrethroids. Nature 2020, 577, 376–380. [Google Scholar] [CrossRef]
- Clarkson, C.S.; Miles, A.; Harding, N.J.; O’Reilly, A.O.; Weetman, D.; Kwiatkowski, D.; Donnelly, M.J. The genetic architecture of target-site resistance to pyrethroid insecticides in the African malaria vectors Anopheles gambiae and Anopheles coluzzii. Mol. Ecol. 2021, 30, 5303–5317. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.; Cowlishaw, R.; Sanou, A.; Ranson, H.; Grigoraki, L. In vivo functional validation of the V402L voltage gated sodium channel mutation in the malaria vector An. gambiae. Pest Manag. Sci. 2022, 78, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Namountougou, M.; Simard, F.; Baldet, T.; Diabaté, A.; Ouédraogo, J.B.; Martin, T.; Dabiré, R.K. Multiple Insecticide Resistance in Anopheles gambiae s.l. Populations from Burkina Faso, West Africa. PLoS ONE 2012, 7, e48412. [Google Scholar] [CrossRef] [Green Version]
- Grau-Bové, X.; Tomlinson, S.; O’Reilly, A.O.; Harding, N.J.; Miles, A.; Kwiatkowski, D.; Donnelly, M.J.; Weetman, D.; The Anopheles gambiae 1000 Genomes Consortium. Evolution of the Insecticide Target Rdl in African Anopheles Is Driven by Interspecific and Interkaryotypic Introgression. Mol. Biol. Evol. 2020, 37, 2900–2917. [Google Scholar] [CrossRef]
- Norris, L.C.; Main, B.J.; Lee, Y.; Collier, T.C.; Fofana, A.; Cornel, A.J.; Lanzaro, G.C. Adaptive introgression in an African malaria mosquito coincident with the increased usage of insecticide-treated bed nets. Proc. Natl. Acad. Sci. USA 2015, 112, 815–820. [Google Scholar] [CrossRef] [Green Version]
- Namountougou, M.; Soma, D.D.; Kientega, M.; Balboné, M.; Kaboré, D.P.A.; Drabo, S.F.; Coulibaly, A.Y.; Fournet, F.; Baldet, T.; Diabaté, A.; et al. Insecticide resistance mechanisms in Anopheles gambiae complex populations from Burkina Faso, West Africa. Acta Trop. 2019, 197, 105054. [Google Scholar] [CrossRef]
- Sanou, A.; Nelli, L.; Guelbéogo, W.M.; Cissé, F.; Tapsoba, M.; Ouédraogo, P.; Sagnon, N.; Ranson, H.; Matthiopoulos, J.; Ferguson, H.M. Insecticide resistance and behavioural adaptation as a response to long-lasting insecticidal net deployment in malaria vectors in the Cascades region of Burkina Faso. Sci. Rep. 2021, 11, 17569. [Google Scholar] [CrossRef]
- MR4. Anopheles Laboratory Biology and Culture. Methods in Anopheles Research; MR4, Atlanta Centers for Disease Control and Prevention: Manassas, VA, USA, 2014; pp. 1–8. [Google Scholar]
- Fanello, C.; Santolamazza, F.; della Torre, A. Simultaneous identification of species and molecular forms of the Anopheles gambiae complex by PCR-RFLP. Med. Vet. Entomol. 2002, 16, 461–464. [Google Scholar] [CrossRef]
- Harris, C.; Lambrechts, L.; Rousset, F.; Abate, L.; Nsango, S.E.; Fontenille, D.; Morlais, I.; Cohuet, A. Polymorphisms in Anopheles gambiae immune genes associated with natural resistance to Plasmodium falciparum. PLoS Pathog. 2010, 6, e1001112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witzig, C.; Parry, M.; Morgan, J.C.; Irving, H.; Steven, A.; Cuamba, N.; Kerah-Hinzoumbé, C.; Ranson, H.; Wondji, C.S. Genetic mapping identifies a major locus spanning P450 clusters associated with pyrethroid resistance in kdr-free Anopheles arabiensis from Chad. Heredity 2013, 110, 389–397. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Test Procedures for Insecticide Resistance Monitoring in Malaria Vector Mosquitoes; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- World Health Organization. Guidelines for Laboratory and Field-Testing of Long-Lasting Insecticidal Nets; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- Bass, C.; Nikou, D.; Donnelly, M.J.; Williamson, M.S.; Ranson, H.; Ball, A.; Vontas, J.; Field, L.M. Detection of knockdown resistance (kdr) mutations in Anopheles gambiae: A comparison of two new high-throughput assays with existing methods. Malar. J. 2007, 6, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, C.M.; Liyanapathirana, M.; Agossa, F.R.; Weetman, D.; Ranson, H.; Donnelly, M.J.; Wilding, C.S. Footprints of positive selection associated with a mutation (N1575Y) in the voltage-gated sodium channel of Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2012, 109, 6614–6619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabire, R.K.; Namountougou, M.; Diabate, A.; Soma, D.D.; Bado, J.; Toe, H.K.; Bass, C.; Combary, P. Distribution and frequency of kdr mutations within Anopheles gambiae s.l. populations and first report of the ace.1 G119S mutation in Anopheles arabiensis from Burkina Faso (West Africa). PLoS One 2014, 9, e101484. [Google Scholar] [CrossRef]
- Bass, C.; Nikou, D.; Vontas, J.; Donnelly, M.J.; Williamson, M.S.; Field, L.M. The Vector Population Monitoring Tool (VPMT): High-Throughput DNA-Based Diagnostics for the Monitoring of Mosquito Vector Populations. Malar. Res. Treat. 2010, 2010, 190434. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, S.N.; Rigden, D.J.; Dowd, A.J.; Lu, F.; Wilding, C.S.; Weetman, D.; Dadzie, S.; Jenkins, A.M.; Regna, K.; Boko, P.; et al. Metabolic and target-site mechanisms combine to confer strong DDT resistance in Anopheles gambiae. PLoS One 2014, 9, e92662. [Google Scholar] [CrossRef]
- Ingham, V.A.; Tennessen, J.A.; Lucas, E.R.; Elg, S.; Yates, H.C.; Carson, J.; Guelbeogo, W.M.; Sagnon, N.; Hughes, G.L.; Heinz, E.; et al. Integration of whole genome sequencing and transcriptomics reveals a complex picture of the reestablishment of insecticide resistance in the major malaria vector Anopheles coluzzii. PLoS Genet. 2021, 17, e1009970. [Google Scholar] [CrossRef]
- Mavridis, K.; Wipf, N.; Medves, S.; Erquiaga, I.; Müller, P.; Vontas, J. Rapid multiplex gene expression assays for monitoring metabolic resistance in the major malaria vector Anopheles gambiae. Parasite Vectors 2019, 12, 9. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Wondji, C.S.; Irving, H.; Morgan, J.; Lobo, N.F.; Collins, F.H.; Hunt, R.H.; Coetzee, M.; Hemingway, J.; Ranson, H. Two duplicated P450 genes are associated with pyrethroid resistance in Anopheles funestus, a major malaria vector. Genome Res. 2009, 19, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Ingham, V.A.; Jones, C.M.; Pignatelli, P.; Balabanidou, V.; Vontas, J.; Wagstaff, S.C.; Moore, J.D.; Ranson, H. Dissecting the organ specificity of insecticide resistance candidate genes in Anopheles gambiae: Known and novel candidate genes. BMC Genom. 2014, 15, 1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kefi, M.; Charamis, J.; Balabanidou, V.; Ioannidis, P.; Ranson, H.; Ingham, V.A.; Vontas, J. Transcriptomic analysis of resistance and short-term induction response to pyrethroids, in Anopheles coluzzii legs. BMC Genom. 2021, 22, 891. [Google Scholar] [CrossRef] [PubMed]
- Pignatelli, P.; Ingham, V.A.; Balabanidou, V.; Vontas, J.; Lycett, G.; Ranson, H. The Anopheles gambiae ATP-binding cassette transporter family: Phylogenetic analysis and tissue localization provide clues on function and role in insecticide resistance. Insect Mol. Biol. 2018, 27, 110–122. [Google Scholar] [CrossRef] [Green Version]
- Vontas, J.; Grigoraki, L.; Morgan, J.; Tsakireli, D.; Fuseini, G.; Segura, L.; Niemczura de Carvalho, J.; Nguema, R.; Weetman, D.; Slotman, M.A.; et al. Rapid selection of a pyrethroid metabolic enzyme CYP9K1 by operational malaria control activities. Proc. Natl. Acad. Sci. USA 2018, 115, 4619–4624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yunta, C.; Hemmings, K.; Stevenson, B.; Koekemoer, L.L.; Matambo, T.; Pignatelli, P.; Voice, M.; Nász, S.; Paine, M.J.I. Cross-resistance profiles of malaria mosquito P450s associated with pyrethroid resistance against WHO insecticides. Pestic. Biochem. Physiol. 2019, 161, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Ingham, V.A.; Wagstaff, S.; Ranson, H. Transcriptomic meta-signatures identified in Anopheles gambiae populations reveal previously undetected insecticide resistance mechanisms. Nat. Commun. 2018, 9, 5282. [Google Scholar] [CrossRef] [Green Version]
- Grigoraki, L.; Grau-Bové, X.; Carrington Yates, H.; Lycett, G.J.; Ranson, H. Isolation and transcriptomic analysis of Anopheles gambiae oenocytes enables the delineation of hydrocarbon biosynthesis. Elife 2020, 9. [Google Scholar] [CrossRef]
- Lynd, A.; Balabanidou, V.; Grosman, R.; Maas, J.; Lian, L.-Y.; Vontas, J.; Lycett, G.J. Development of a functional genetic tool for Anopheles gambiae oenocyte characterisation: Application to cuticular hydrocarbon synthesis. bioRxiv 2019, 742619. [Google Scholar]
- Hughes, A.; Lissenden, N.; Viana, M.; Toé, K.H.; Ranson, H. Anopheles gambiae populations from Burkina Faso show minimal delayed mortality after exposure to insecticide-treated nets. Parasites Vectors 2020, 13, 17. [Google Scholar] [CrossRef] [Green Version]
- Toé, K.H.; Jones, C.M.; N’Fale, S.; Ismail, H.M.; Dabiré, R.K.; Ranson, H. Increased pyrethroid resistance in malaria vectors and decreased bed net effectiveness, Burkina Faso. Emerg. Infect. Dis. 2014, 20, 1691–1696. [Google Scholar] [CrossRef] [PubMed]
- Toé, K.H.; N’Falé, S.; Dabiré, R.K.; Ranson, H.; Jones, C.M. The recent escalation in strength of pyrethroid resistance in Anopheles coluzzi in West Africa is linked to increased expression of multiple gene families. BMC Genom. 2015, 16, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badolo, A.; Traore, A.; Jones, C.M.; Sanou, A.; Flood, L.; Guelbeogo, W.M.; Ranson, H.; Sagnon, N.F. Three years of insecticide resistance monitoring in Anopheles gambiae in Burkina Faso: Resistance on the rise? Malar. J. 2012, 11, 232. [Google Scholar] [CrossRef] [Green Version]
- Gansané, A.; Candrinho, B.; Mbituyumuremyi, A.; Uhomoibhi, P.; Nfalé, S.; Mohammed, A.B.; Guelbeogo, W.M.; Sanou, A.; Kangoye, D.; Debe, S.; et al. Design and methods for a quasi-experimental pilot study to evaluate the impact of dual active ingredient insecticide-treated nets on malaria burden in five regions in sub-Saharan Africa. Malar. J. 2022, 21, 19. [Google Scholar] [CrossRef] [PubMed]
- Glynne-Jones, D.E. Piperonyl Butoxide: The Insecticide Synergist. Academic Press: San Diego, California, USA, 1998; p. 323. [Google Scholar]
- Toe, K.H.; Müller, P.; Badolo, A.; Traore, A.; Sagnon, N.; Dabiré, R.K.; Ranson, H. Do bednets including piperonyl butoxide offer additional protection against populations of Anopheles gambiae s.l. that are highly resistant to pyrethroids? An experimental hut evaluation in Burkina Fasov. Med. Vet. Entomol. 2018, 32, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Pinto, J.; Lynd, A.; Vicente, J.L.; Santolamazza, F.; Randle, N.P.; Gentile, G.; Moreno, M.; Simard, F.; Charlwood, J.D.; do Rosário, V.E.; et al. Multiple origins of knockdown resistance mutations in the Afrotropical mosquito vector Anopheles gambiae. PLoS One 2007, 2, e1243. [Google Scholar] [CrossRef] [PubMed]
- Edi, C.V.; Djogbénou, L.; Jenkins, A.M.; Regna, K.; Muskavitch, M.A.; Poupardin, R.; Jones, C.M.; Essandoh, J.; Kétoh, G.K.; Paine, M.J.; et al. CYP6 P450 enzymes and ACE-1 duplication produce extreme and multiple insecticide resistance in the malaria mosquito Anopheles gambiae. PLoS Genet. 2014, 10, e1004236. [Google Scholar] [CrossRef]
- Abdalla, H.; Wilding, C.S.; Nardini, L.; Pignatelli, P.; Koekemoer, L.L.; Ranson, H.; Coetzee, M. Insecticide resistance in Anopheles arabiensis in Sudan: Temporal trends and underlying mechanisms. Parasites Vectors 2014, 7, 213. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.M.; Toé, H.K.; Sanou, A.; Namountougou, M.; Hughes, A.; Diabaté, A.; Dabiré, R.; Simard, F.; Ranson, H. Additional selection for insecticide resistance in urban malaria vectors: DDT resistance in Anopheles arabiensis from Bobo-Dioulasso, Burkina Faso. PLoS One 2012, 7, e45995. [Google Scholar] [CrossRef]
- Balabanidou, V.; Kefi, M.; Aivaliotis, M.; Koidou, V.; Girotti, J.R.; Mijailovsky, S.J.; Juárez, M.P.; Papadogiorgaki, E.; Chalepakis, G.; Kampouraki, A.; et al. Mosquitoes cloak their legs to resist insecticides. Proc. Biol. Sci. 2019, 286, 20191091. [Google Scholar] [CrossRef] [Green Version]
- Yahouédo, G.A.; Chandre, F.; Rossignol, M.; Ginibre, C.; Balabanidou, V.; Mendez, N.G.A.; Pigeon, O.; Vontas, J.; Cornelie, S. Contributions of cuticle permeability and enzyme detoxification to pyrethroid resistance in the major malaria vector Anopheles gambiae. Sci. Rep. 2017, 7, 11091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reidenbach, K.R.; Cheng, C.; Liu, F.; Liu, C.; Besansky, N.J.; Syed, Z. Cuticular differences associated with aridity acclimation in African malaria vectors carrying alternative arrangements of inversion 2La. Parasites Vectors 2014, 7, 176. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Q.; Jiang, H.B.; Liu, Y.; Fan, J.Y.; Ma, Y.J.; Yuan, C.Y.; Lou, B.H.; Wang, J.J. Odorant binding protein 2 reduces imidacloprid susceptibility of Diaphorina citri. Pestic. Biochem. Physiol. 2020, 168, 104642. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Q.; Yan, Q.; Li, L.L.; Xu, J.W.; Mang, D.; Wang, X.L.; Hoh, H.H.; Ye, J.; Ju, Q.; Ma, Y.; et al. Different binding properties of two general-odorant binding proteins in Athetis lepigone with sex pheromones, host plant volatiles and insecticides. Pestic. Biochem. Physiol. 2020, 164, 173–182. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Species | Origin | Source | Year Colony Established |
|---|---|---|---|---|
| Kisumu (susceptible strain) | An. gambiae (s.s.) | Kenya | MR4 | 1975 |
| N’Gousso (susceptible strain) | An. coluzzii | Cameroon | CRID | 2006 |
| Moz (susceptible strain) | An. arabiensis | Chokwe, Southern Mozambique (24° 33′ 37″ S, 33° 1′ 20″ E) | Established in LSTM from field collections performed by JCM with assistance from National Institute of Health, Mozambique [23] | 2009 |
| VK7 2014 | An. coluzzii | Houet Province, Burkina Faso Valley de Kou 7 (11°24′29″ N, 4°24′37″ W) | Established from larval collections performed by LSTM (JCM) and CNRFP (KHT) | 2015 |
| Banfora M | An. coluzzii | Comoé Province Burkina Faso Banfora district (Tiefora, Pont Maurice, Sikane and Djomale (10° 38′ 0″ N, 4° 33′ 0″ W) and Bakaridjan (10°24′26.34″ N, 4°33′44.78″ W) villages) | Established from larval collections performed by LSTM (JCM) and CNRFP (KHT) | 2015 |
| Bakaridjan | An. gambiae (s.s) | Comoé Province Burkina Faso Banfora district (Tiefora, Pont Maurice, Sikane and Djomale (10° 38′ 0″ N, 4° 33′ 0″ W) and Bakaridjan (10°24′26.34″ N, 4°33′44.78″ W) villages) | Established from larval collections performed by LSTM (JCM) and CNRFP (KHT) | 2015 |
| Tiefora | An. coluzzii | Comoé Province, Burkina Faso Banfora district (10° 37.447’ N, 4° 33.201’ W) | Established from larval collections performed by LSTM (JCM) and CNRFP (KHT) | 2018 |
| Gaoua-ara | An. arabiensis | Poni Province Burkina Faso Gaoua district (10.3231° N, 3.1679° W) | Established from larval collections performed by IRSS (ASH) | 2018 |
| An. coluzzii | An. gambaie | An. arabiensis | |||
|---|---|---|---|---|---|
| VK72014 | Banfora | Tiefora | Bakaridjan | Gaoua-ara | |
| CYP4G16 | |||||
| CYP6M2 | |||||
| CYP6P1 | |||||
| CYP6P3 | |||||
| CYP6P4 | |||||
| CYP6Z1 | |||||
| CYP9K1 | |||||
| GSTE2 | |||||
| Genes up-regulated in both qPCR and RNAseq data set | |||||
| Genes up-regulated in qPCR data set only | |||||
| Genes up-regulated in RNAseq data set only | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, J.; Ingham, V.A.; Morris, M.; Toé, K.H.; Hien, A.S.; Morgan, J.C.; Dabiré, R.K.; Guelbéogo, W.M.; Sagnon, N.; Ranson, H. Sympatric Populations of the Anopheles gambiae Complex in Southwest Burkina Faso Evolve Multiple Diverse Resistance Mechanisms in Response to Intense Selection Pressure with Pyrethroids. Insects 2022, 13, 247. https://doi.org/10.3390/insects13030247
Williams J, Ingham VA, Morris M, Toé KH, Hien AS, Morgan JC, Dabiré RK, Guelbéogo WM, Sagnon N, Ranson H. Sympatric Populations of the Anopheles gambiae Complex in Southwest Burkina Faso Evolve Multiple Diverse Resistance Mechanisms in Response to Intense Selection Pressure with Pyrethroids. Insects. 2022; 13(3):247. https://doi.org/10.3390/insects13030247
Chicago/Turabian StyleWilliams, Jessica, Victoria A. Ingham, Marion Morris, Kobié Hyacinthe Toé, Aristide S. Hien, John C. Morgan, Roch K. Dabiré, Wamdagogo Moussa Guelbéogo, N’Falé Sagnon, and Hilary Ranson. 2022. "Sympatric Populations of the Anopheles gambiae Complex in Southwest Burkina Faso Evolve Multiple Diverse Resistance Mechanisms in Response to Intense Selection Pressure with Pyrethroids" Insects 13, no. 3: 247. https://doi.org/10.3390/insects13030247
APA StyleWilliams, J., Ingham, V. A., Morris, M., Toé, K. H., Hien, A. S., Morgan, J. C., Dabiré, R. K., Guelbéogo, W. M., Sagnon, N., & Ranson, H. (2022). Sympatric Populations of the Anopheles gambiae Complex in Southwest Burkina Faso Evolve Multiple Diverse Resistance Mechanisms in Response to Intense Selection Pressure with Pyrethroids. Insects, 13(3), 247. https://doi.org/10.3390/insects13030247