
Identification and Characterization of Chemosensory Receptors in the Pheromone Gland-Ovipositor of Spodoptera frugiperda (J. E. Smith)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Tissue Collection
2.3. RNA Extraction, cDNA Library Construction, and Illumina Sequencing
2.4. De Novo Assembly of Short Reads
2.5. Gene Annotation and Identification of Chemosensory Receptors
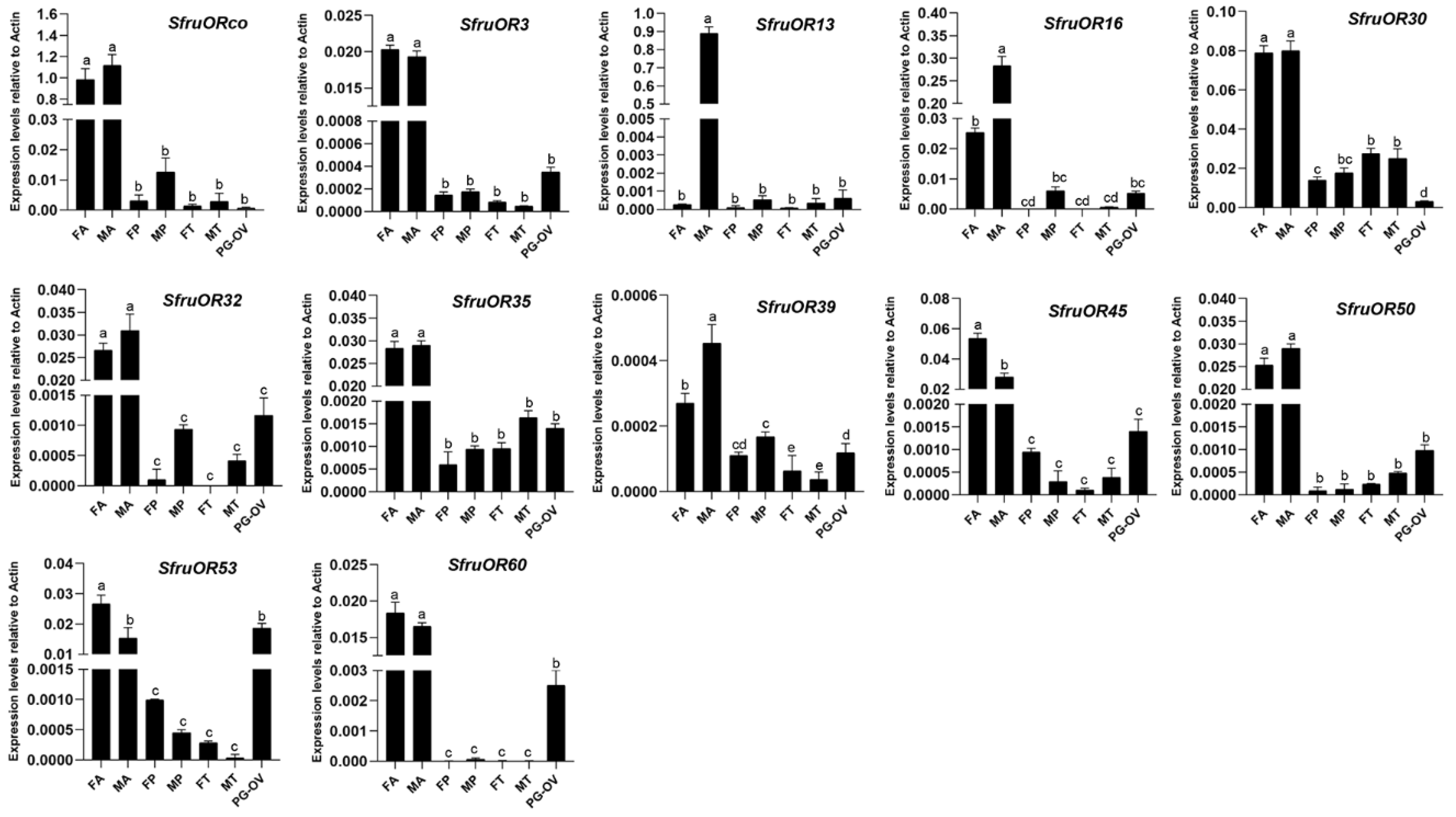
2.6. Expression Profiling by RT-qPCR
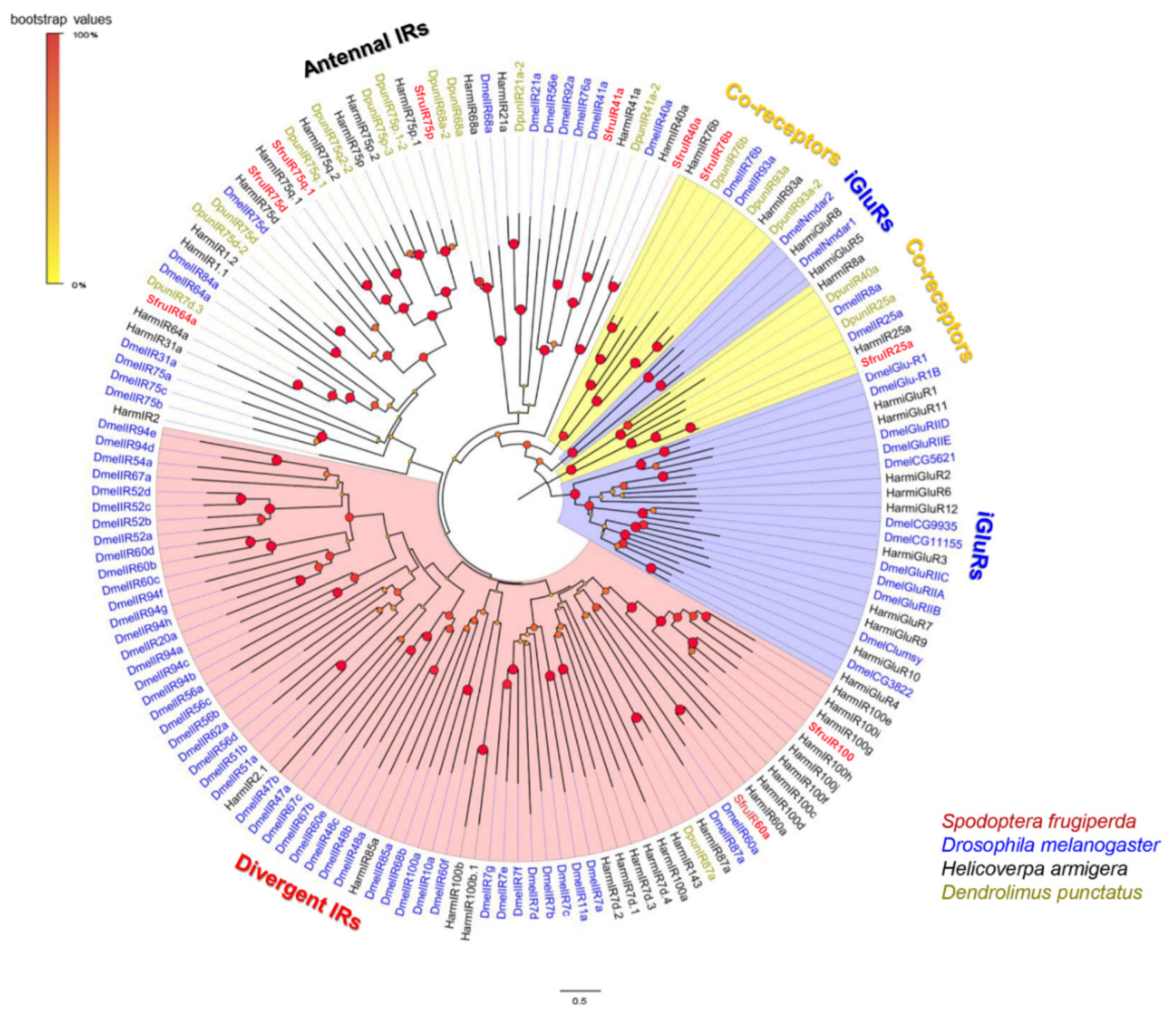
2.7. Phylogenetic Analyses
3. Results
3.1. Transcriptome Sequencing and Sequence Assembly
3.2. GO Annotation and Classification
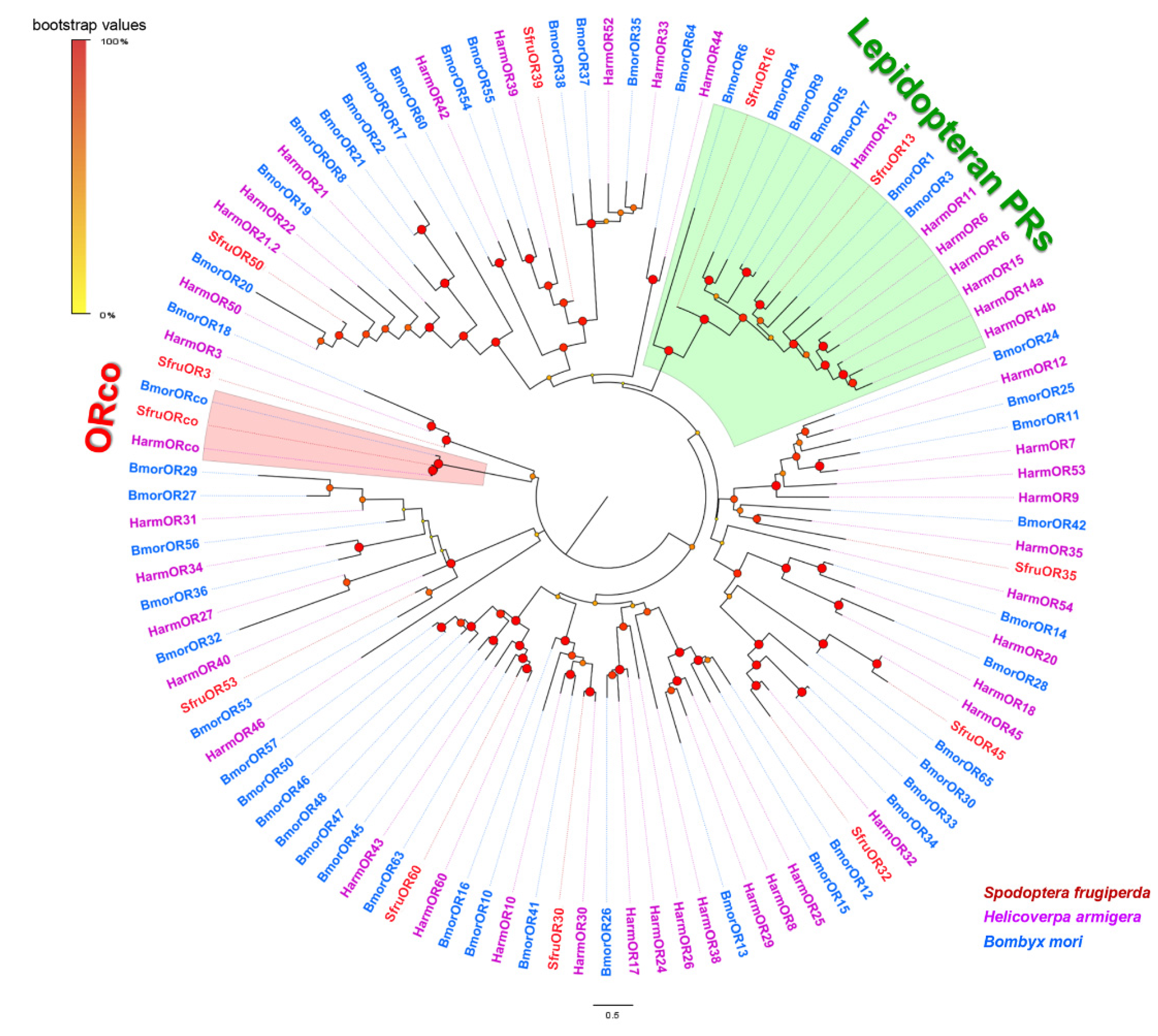
3.3. Analysis of Odorant Receptors
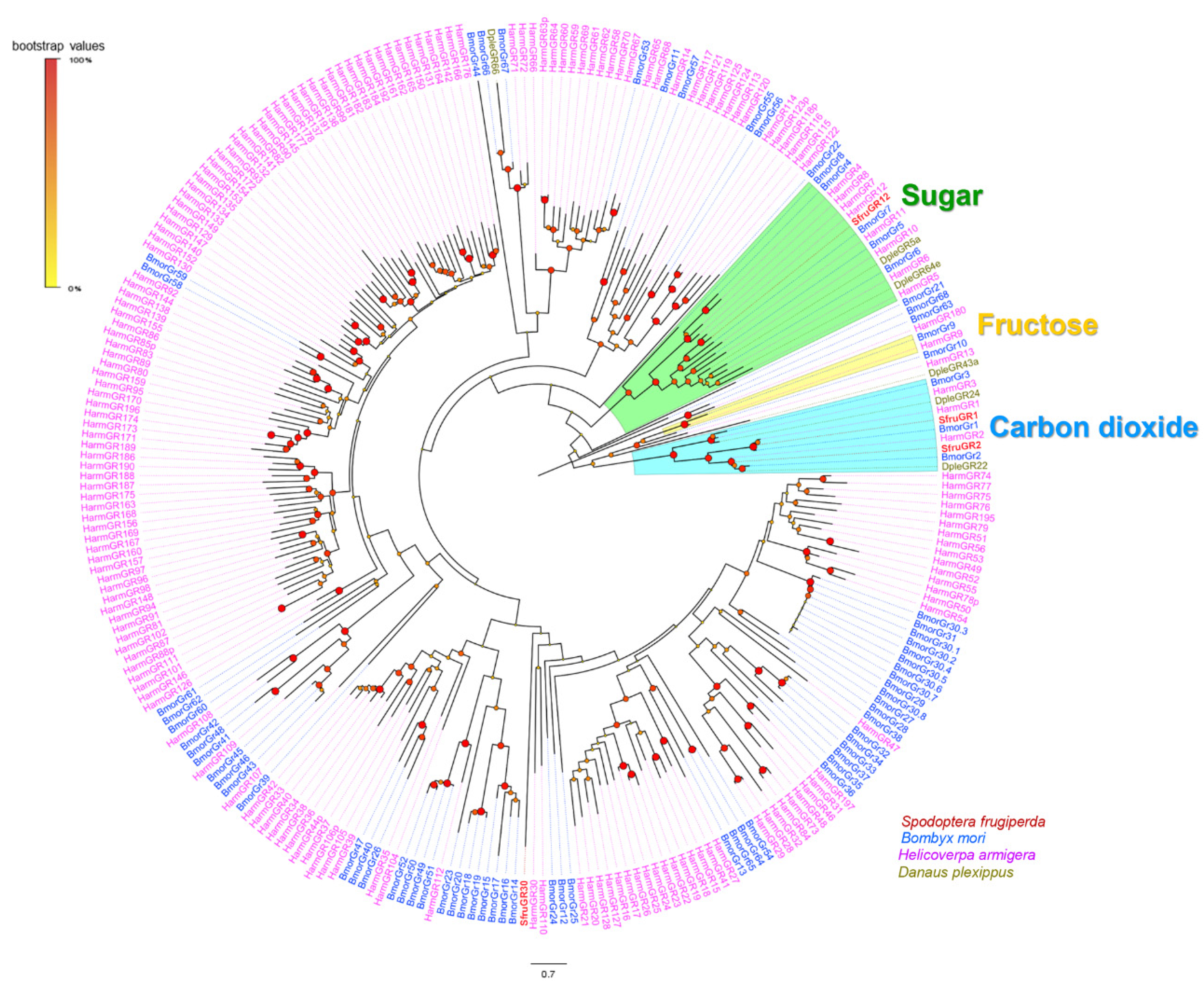
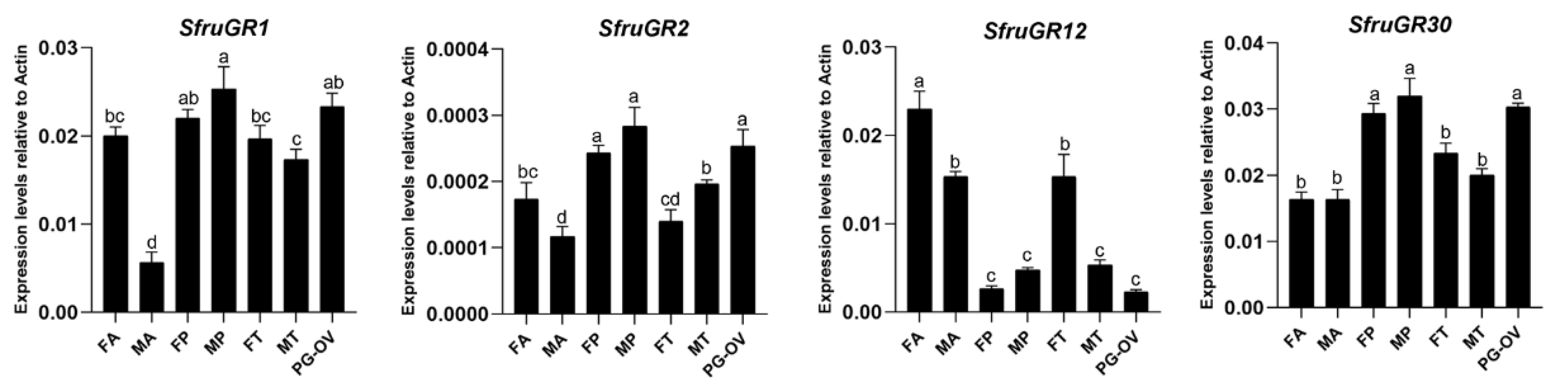
3.4. Analysis of Gustatory Receptors
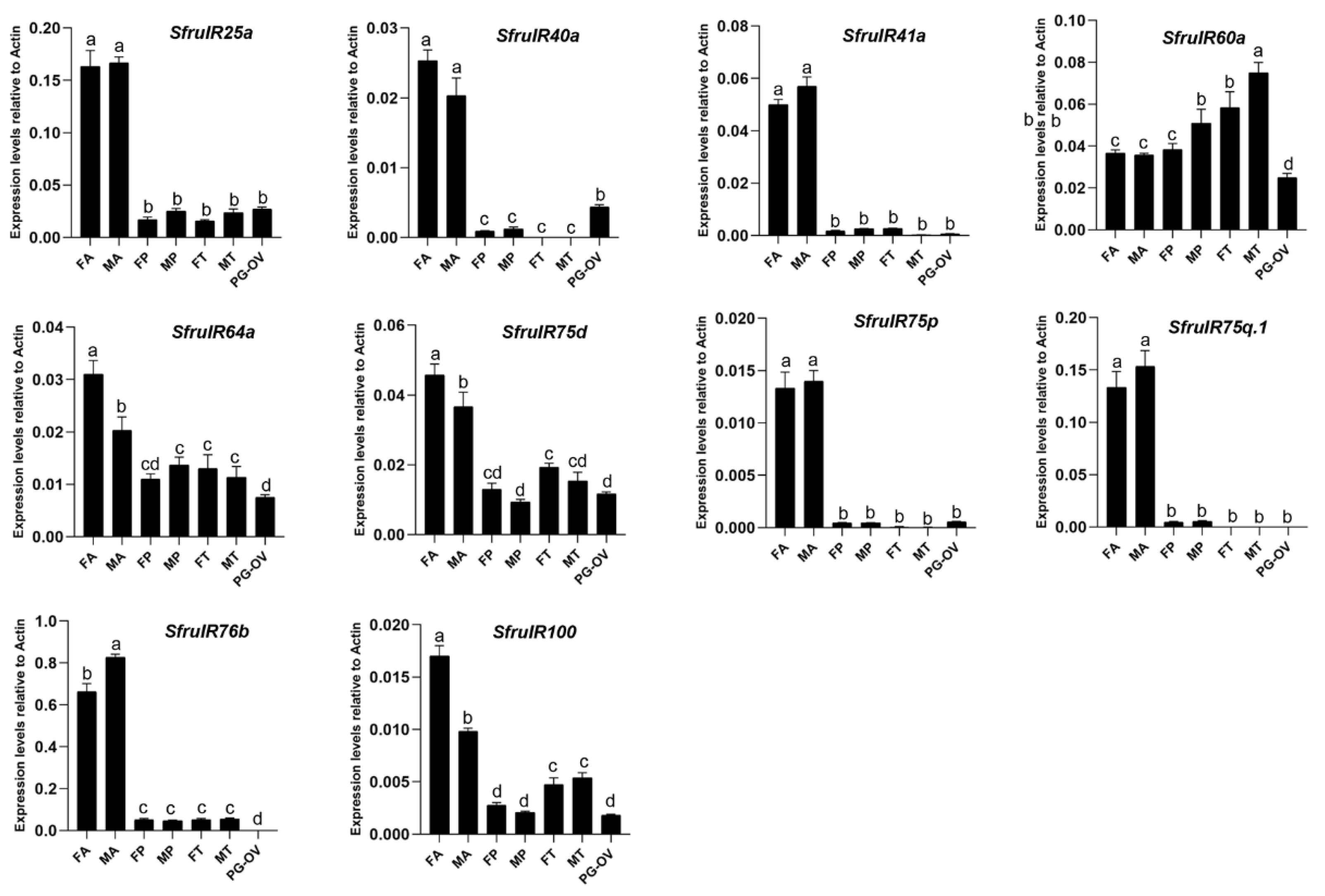
3.5. Analysis of Ionotropic Receptors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dahanukar, A.; Hallem, E.A.; Carlson, J.R. Insect chemoreception. Curr. Opin. Neurobiol. 2005, 15, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Hansson, B.S.; Stensmyr, M.C. Evolution of insect olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Qiu, Y.T.; Wang, G.R.; Kwon, J.Y.; Rutzler, M.; Kwon, H.W.; Pitts, R.J.; van Loon, J.J.; Takken, W.; Carlson, J.R.; et al. Odor coding in the maxillary palp of the malaria vector mosquito Anopheles gambiae. Curr. Biol. 2007, 17, 1533–1544. [Google Scholar] [CrossRef] [Green Version]
- Haverkamp, A.; Yon, F.; Keesey, I.W.; Missbach, C.; Koenig, C.; Hansson, B.S.; Baldwin, I.T.; Knaden, M.; Kessler, D. Hawkmoths evaluate scenting flowers with the tip of their proboscis. eLife 2016, 5, e15039. [Google Scholar] [CrossRef] [Green Version]
- Stocker, R.F. The organization of the chemosensory system in Drosophila melanogaster: A review. Cell Tissue Res. 1994, 275, 3–26. [Google Scholar] [CrossRef]
- Kaissling, K.E. Peripheral mechanisms of pheromone reception in moths. Chem. Sens. 1996, 21, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, H.R.; Benton, R. Molecular mechanisms of olfactory detection in insects: Beyond receptors. Open Biol. 2020, 10, 200252. [Google Scholar] [CrossRef]
- Clyne, P.J.; Warr, C.G.; Freeman, M.R.; Lessing, D.; Kim, J.; Carlson, J.R. A novel family of divergent seven-transmembrane proteins: Candidate odorant receptors in Drosophila. Neuron 1999, 22, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Chess, A. Identification of candidate Drosophila olfactory receptors from genomic DNA sequence. Genomics 1999, 60, 31–39. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Amrein, H.; Morozov, P.S.; Rzhetsky, A.; Axel, R. A spatial map of olfactory receptor expression in the Drosophila antenna. Cell 1999, 96, 725–736. [Google Scholar] [CrossRef] [Green Version]
- Clyne, P.J. Candidate taste receptors in Drosophila. Science 2000, 287, 1830–1834. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.; Brady, R., Jr.; Cravchik, A.; Morozov, P.; Rzhetsky, A.; Zuker, C.; Axel, R. A chemosensory gene family encoding candidate gustatory and olfactory receptor in Drosophila. Cell 2001, 104, 661–673. [Google Scholar] [CrossRef]
- Robertson, H.M. Molecular evolution of the major arthropod chemoreceptor gene families. Annu. Rev. Entomol. 2018, 64, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Benton, R.; Sachse, S.; Michnick, S.W.; Vosshall, L.B. Atypical membrane topology and heteromeric function of Drosophila odorant receptors in vivo. PLoS Biol. 2006, 4, e20. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Pellegrino, M.; Nakagawa, T.; Vosshall, L.B.; Touhara, K. Insect olfactory receptors are heteromeric ligand-gated ion channels. Nature 2008, 452, 1002–1006. [Google Scholar] [CrossRef] [PubMed]
- Wicher, D.; Schäfer, R.; Bauernfeind, R.; Stensmyr, M.C.; Heller, R.; Heinemann, S.H.; Hansson, B.S. Drosophila odorant receptors are both ligand-gated and cyclic-nucleotide-activated cation channels. Nature 2008, 452, 1007–1011. [Google Scholar] [CrossRef]
- Butterwick, J.A.; Del Mármol, J.; Kim, K.H.; Kahlson, M.A.; Rogow, J.A.; Walz, T.; Ruta, V. Cryo-EM structure of the insect olfactory receptor Orco. Nature 2018, 560, 447–452. [Google Scholar] [CrossRef]
- Hallem, E.A.; Carlson, J.R. Coding of odors by a receptor repertoire. Cell 2006, 125, 143–160. [Google Scholar] [CrossRef] [Green Version]
- Kirkness, E.F.; Haas, B.J.; Sun, W.; Braig, H.R.; Perotti, M.A.; Clark, J.M.; Lee, S.H.; Robertson, H.M.; Kennedy, R.C.; Elhaik, E.; et al. Genome sequences of the human body louse and its primary endosymbiont provide insights into the permanent parasitic lifestyle. Proc. Natl. Acad. Sci. USA 2010, 107, 12168–12173. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Rokas, A.; Berger, S.L.; Liebig, J.; Ray, A.; Zwiebel, L.J. Chemoreceptor evolution in Hymenoptera and its implications for the evolution of eusociality. Genome Biol. Evol. 2015, 7, 2407–2416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.L.; Dong, J.F.; Gu, N.; Wang, S.L. Identification of candidate chemosensory receptors in the antennae of the variegated cutworm, Peridroma saucia Hübner, based on a transcriptomic analysis. Front. Physiol. 2020, 11, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Walker, W.B.; Wang, G.R. Pheromone reception in moths: From molecules to behaviors. Prog. Mol. Biol. Transl. 2014, 130, 109–128. [Google Scholar]
- Yang, K.; Wang, C.Z. Review of pheromone receptors in heliothine species: Expression, function, and evolution. Entomol. Exp. Appl. 2021, 169, 156–171. [Google Scholar] [CrossRef]
- Agnihotri, A.R.; Roy, A.A.; Joshi, R.S. Gustatory receptors in Lepidoptera: Chemosensation and beyond. Insect Mol. Biol. 2016, 25, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.D.; Cayirlioglu, P.; Kadow, I.G.; Vosshall, L.B. Two chemosensory receptors together mediate carbon dioxide detection in Drosophila. Nature 2007, 445, 86–90. [Google Scholar] [CrossRef]
- Fujii, S.; Yavuz, A.; Slone, J.; Jagge, C.; Song, X.; Amrein, H. Drosophila sugar receptors in sweet taste perception, olfaction, and internal nutrient sensing. Curr. Biol. 2015, 25, 621–627. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Tanaka, K.; Touhara, K. Sugar-regulated cation channel formed by an insect gustatory receptor. Proc. Natl. Acad. Sci. USA 2011, 108, 11680–11685. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, T.; Slone, J.; Song, X.; Amrein, H. A fructose receptor functions as a nutrient sensor in the Drosophila brain. Cell 2012, 151, 1113–1125. [Google Scholar] [CrossRef] [Green Version]
- Delventhal, R.; Carlson, J.R. Bitter taste receptors confer diverse functions to neurons. eLife 2016, 5, e11181. [Google Scholar] [CrossRef]
- Xu, W.; Papanicolaou, A.; Zhang, H.J.; Anderson, A. Expansion of a bitter taste receptor family in a polyphagous insect herbivore. Sci. Rep. 2016, 6, 23666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benton, R.; Vannice, K.S.; Gomez-Diaz, C.; Vosshall, L.B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Buhl, E.; Xu, M.; Croset, V.; Rees, J.S.; Lilley, K.S.; Benton, R.; Hodge, J.J.L.; Stanewsky, R. Drosophila ionotropic receptor 25a mediates circadian clock resetting by temperature. Nature 2015, 527, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Enjin, A.; Zaharieva, E.E.; Frank, D.D.; Mansourian, S.; Suh, G.S.; Gallio, M.; Stensmyr, M.C. Humidity sensing in Drosophila. Curr. Biol. 2016, 26, 1352–1358. [Google Scholar] [CrossRef] [PubMed]
- Knecht, Z.A.; Silbering, A.F.; Ni, L.N.; Klein, M.; Budelli, G.; Bell, R.; Abuin, L.; Ferrer, A.J.; Samuel, A.D.T.; Benton, R.; et al. Distinct combinations of variant ionotropic glutamate receptors mediate thermosensation and hygrosensation in Drosophila. eLife 2016, 5, e17879. [Google Scholar] [CrossRef]
- Knecht, Z.A.; Silbering, A.F.; Cruz, J.; Yang, L.; Croset, V.; Benton, R.; Garrity, P.A. Ionotropic receptor-dependent moist and dry cells control hygrosensation in Drosophila. eLife 2017, 6, e26654. [Google Scholar] [CrossRef]
- Chen, H.L.; Stern, U.; Yang, C.H. Molecular control limiting sensitivity of sweet taste neurons in Drosophila. Proc. Natl. Acad. Sci. USA 2019, 116, 20158–20168. [Google Scholar] [CrossRef] [Green Version]
- Abuin, L.; Bargeton, B.; Ulbrich, M.H.; Isacoff, E.Y.; Kellenberger, S.; Benton, R. Functional architecture of olfactory ionotropic glutamate receptors. Neuron 2011, 69, 44–60. [Google Scholar] [CrossRef] [Green Version]
- Widmayer, P.; Heifetz, Y.; Breer, H. Expression of a pheromone receptor in ovipositor sensilla of the female moth (Heliothis virescens). Insect Mol. Biol. 2009, 18, 541–547. [Google Scholar] [CrossRef]
- Klinner, C.F.; König, C.; Missbach, C.; Werckenthin, A.; Daly, K.C.; Bisch-Knaden, S.; Stengl, M.; Hansson, B.S.; Große-Wilde, E. Functional olfactory sensory neurons housed in olfactory sensilla on the ovipositor of the hawkmoth Manduca sexta. Front. Ecol. Evol. 2016, 4, 130. [Google Scholar] [CrossRef] [Green Version]
- Li, R.T.; Huang, L.Q.; Dong, J.F.; Wang, C.Z. A moth odorant receptor highly expressed in the ovipositor is involved in detecting host-plant volatiles. eLife 2020, 9, e53706. [Google Scholar] [CrossRef] [PubMed]
- Sparks, A.N. A review of the biology of the fall armyworm. Flo. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Otim, M.H.; Tay, W.T.; Walsh, T.K.; Kanyesigye, D.; Adumo, S.; Abongosi, J.; Ochen, S.; Sserumaga, J.; Alibu, S.; Abalo, G.; et al. Detection of sister-species in invasive populations of the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) from Uganda. PLoS ONE 2018, 13, e0194571. [Google Scholar] [CrossRef] [Green Version]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamo, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [Green Version]
- Cock, M.J.W.; Beseh, P.K.; Buddie, A.G.; Cafá, G.; Crozier, J. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring the spread of invasive species in developing countries. Sci. Rep. 2017, 7, 4103. [Google Scholar] [CrossRef] [PubMed]
- Mallapur, C.P.; Naik, A.K.; Hagari, S.; Prabhu, S.T.; Patil, P.K. Status of alien pest fall armyworm, Spodoptera frugiperda (J.E. Smith) on maize in Northern Karnataka. J. Entomol. Zool. Stud. 2018, 6, 432–436. [Google Scholar]
- Wu, Q.L.; Jiang, Y.Y.; Wu, K.M. Analysis of migration routes of the fall armyworm Spodoptera frugiperda (J.E. Smith) from Myanmar to China. Plant Prot. 2019, 45, 1–6. [Google Scholar]
- Zhang, L.; Jin, M.H.; Zhang, D.D.; Jiang, Y.Y.; Liu, J.; Wu, K.M.; Xiao, Y.T. Molecular identification of invasive fall armyworm Spodoptera frugiperda in Yunnan province. Plant Protec. 2019, 45, 19–24. [Google Scholar]
- Sun, X.X.; Hu, C.X.; Jia, H.R.; Wu, Q.L.; Shen, X.J.; Zhao, S.Y.; Yang, Y.Y.; Wu, K.M. Case study on the first immigration of fall armyworm, Spodoptera frugiperda invading into China. J. Integr. Agr. 2021, 20, 664–672. [Google Scholar] [CrossRef]
- Sena Jr, D.G.; Pinto, F.A.C.; Queiroz, D.M.; Viana, P.A. Fall armyworm damaged maize plant identification using digital images. Biosyst. Eng. 2003, 85, 449–454. [Google Scholar] [CrossRef]
- Tumlinson, J.H.; Mitchell, E.R.; Teal, P.E.A.; Heath, R.R.; Mengelkoch, L.J. Sex pheromone of fall armyworm, Spodoptera frugiperda (J.E. Smith): Identification of components critical to attraction in the field. J. Chem. Ecol. 1986, 12, 1909–1926. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.J.; Mo, B.T.; Guo, H.; Yang, J.; Tang, R.; Wang, C.Z. Revisiting the sex pheromone of the fall armyworm Spodoptera frugiperda, a new invasive pest in South China. Insect Sci. 2021, in press . [Google Scholar] [CrossRef] [PubMed]
- Yactayo-Chang, J.P.; Mendoza, J.; Willms, S.; Rering, C.C.; Beck, J.J.; Block, A.K. Zea mays volatiles that influence oviposition and feeding behaviors of Spodoptera frugiperda. J. Chem. Ecol. 2021, 47, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; He, L.; Tan, X.; Zhang, Z.; Wang, Y.; Li, X.; He, H.; Ding, W.; Li, Y. Identification and phylogenetics of Spodoptera frugiperda chemosensory proteins based on antennal transcriptome data. Com. Biochem. Physiol. Part D Genom. Proteom. 2020, 34, 100680. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.M.; Liu, X.L.; Liu, S.R.; Wei, Z.Q.; Han, W.K.; Guo, Y.; Dong, S.L. Functional characterization of sex pheromone receptors in the fall armyworm (Spodoptera frugiperda). Insects 2020, 11, 193. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Gong, X.L.; Li, G.C.; Mo, B.T.; Jiang, N.J.; Huang, L.Q.; Wang, C.Z. Functional analysis of pheromone receptor repertoire in the fall armyworm, Spodoptera frugiperda. Pest Manag. Sci. 2022, in press . [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.L.; Dong, J.F.; Ning, C.; Ding, P.P.; Huang, L.Q.; Wang, C.Z. An odorant receptor mediates the attractiveness of cis-jasmone to Campoletis chlorideae, the endoparasitoid of Helicoverpa armigera. Insect Mol. Biol. 2019, 28, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Zhang, Y.; Cui, H.H.; Liu, J.W.; Wu, Y.Q.; Cheng, Y.; Xu, H.X.; Huang, X.X.; Li, S.T.; Zhou, A.; et al. WEGO 2.0: A web tool for analyzing and plotting GO annotations, 2018 update. Nucleic Acids Res. 2018, 46, W71–W75. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTALW: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Gouin, A.; Bretaudeau, A.; Nam, K.; Gimenez, S.; Aury, J.M.; Duvic, B.; Hilliou, F.; Durand, N.; Montagne, N.; Darboux, I.; et al. Two genomes of highly polyphagous lepidopteran pests (Spodoptera frugiperda, Noctuidae) with different host-plant ranges. Sci. Rep. 2017, 7, 11816. [Google Scholar] [CrossRef]
- Anderson, A.R.; Wanner, K.W.; Trowell, S.C.; Warr, C.G.; Jaquin-Joly, E.; Zagatti, P.; Robertson, H.; Newcomb, R.D. Molecular basis of female-specific odorant responses in Bombyx mori. Insect Biochem. Mol. Biol. 2009, 39, 189–197. [Google Scholar] [CrossRef]
- Ning, C.; Yang, K.; Xu, M.; Huang, L.Q.; Wang, C.Z. Functional validation of the carbon dioxide receptor in labial palps of Helicoverpa armigera moths. Insect Biochem. Mol. Biol. 2016, 73, 12–19. [Google Scholar] [CrossRef]
- Wanner, K.W.; Anderson, A.R.; Trowell, S.C.; Theilmann, D.A.; Robertson, H.M.; Newcomb, R.D. Female-biased expression of odorant receptor genes in the adult antennae of the silkworm, Bombyx mori. Insect Mol. Biol. 2007, 16, 107–119. [Google Scholar] [CrossRef]
- Faucheux, M.J. Multiporous sensilla on the ovipositor of Monopis krocicapitella Clem (Lepidoptera, Tineidae). Int. J. Insect Morphol. Embryol. 1988, 17, 473–475. [Google Scholar] [CrossRef]
- Anderson, P.; Hallberg, E. Structure and distribution of tactile and bimodal taste tactile sensilla on the ovipositor, tarsi and antennae of the flour moth, Ephestia kuehniella (Zeller) (Lepidoptera, Pyralidae). Int. J. Insect Morphol. Embryol. 1990, 19, 13–23. [Google Scholar] [CrossRef]
- Ha, T.S.; Smith, D.P. Odorant and pheromone receptors in insects. Front. Cell. Neurosci. 2009, 3, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glaser, N.; Gallot, A.; Legeai, F.; Montagné, N.; Poivet, E.; Harry, M.; Calatayud, P.A.; Jacquin-Joly, E. Candidate chemosensory genes in the stemborer Sesamia nonagrioides. Int. J. Biol. Sci. 2013, 9, 481–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.H.; Zhang, Y.N.; Hou, X.Q.; Li, F.; Dong, S.L. Large number of putative chemoreception and pheromone biosynthesis genes revealed by analyzing transcriptome from ovipositor-pheromone glands of Chilo suppressalis. Sci. Rep. 2015, 5, 7888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, C.; Ning, C.; Huang, L.Q.; Wang, C.Z. Design of larval chemical attractants based on odorant response spectra of odorant receptors in the cotton bollworm. Insect Biochem. Mol. Biol. 2017, 84, 48–62. [Google Scholar] [CrossRef]
- Rostelien, T.; Stranden, M.; Borg-Karlson, A.K.; Mustaparta, H. Olfactory receptor neurons in two heliothine moth species responding selectively to aliphatic green leaf volatiles, aromatic compounds, monoterpenes and sesquiterpenes of plant origin. Chem. Sens. 2005, 30, 443–461. [Google Scholar] [CrossRef]
- de Fouchier, A.; Walker, W.B.; Montagné, N.; Steiner, C.; Binyameen, M.; Schlyter, F.; Chertemps, T.; Maria, A.; François, M.-C.; Monsempes, C.; et al. Functional evolution of Lepidoptera olfactory receptors revealed by deorphanization of a moth repertoire. Nat. Commun. 2017, 30, 15709. [Google Scholar] [CrossRef]
- Guo, M.; Du, L.; Chen, Q.; Feng, Y.; Zhang, J.; Zhang, X.; Tian, K.; Cao, S.; Huang, T.; Jacquin-Joly, E.; et al. Odorant receptors for detecting flowering plant cues are functionally conserved across moths and butterflies. Mol. Biol. Evol. 2021, 38, 1413–1427. [Google Scholar] [CrossRef]
- Luo, Y.; Carlson, J.R. Sensory biology: Structure of an insect chemoreceptor. Curr. Biol. 2018, 28, 1202–1205. [Google Scholar] [CrossRef] [Green Version]
- Spehr, M.; Gisselmann, G.; Poplawski, A.; Riffell, J.A.; Wetzel, C.H.; Zimmer, R.K.; Hatt, H. Identification of a testicular odorant receptor mediating human sperm chemotaxis. Science 2003, 299, 2054–2058. [Google Scholar] [CrossRef]
- Pitts, R.J.; Liu, C.; Zhou, X.; Malpartida, J.C.; Zwiebel, L.J. Odorant receptor-mediated sperm activation in disease vector mosquitoes. Proc. Natl. Acad. Sci. USA 2014, 111, 2566–2571. [Google Scholar] [CrossRef] [Green Version]
- Vosshall, L.B.; Stocker, R.F. Molecular architecture of smell and taste in Drosophila. Annu. Rev. Neurosci. 2007, 30, 505–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, H.M.; Kent, L.B. Evolution of the gene lineage encoding the carbon dioxide receptor in insects. J. Insect Sci. 2009, 9, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W. How do moth and butterfly taste? —Molecular basis of gustatory receptors in Lepidoptera. Insect Sci. 2020, 27, 1148–1157. [Google Scholar] [CrossRef] [PubMed]
- Seada, M.A.; Ignell, R.; Anderson, P. Morphology and distribution of ovipositor sensilla of female cotton leaf worm Spodoptera littoralis (Lepidoptera: Noctuidae), and evidence for gustatory function. Entomol. Sci. 2016, 19, 9–19. [Google Scholar] [CrossRef]
- Thom, C.; Guerenstein, P.G.; Mechaber, W.L.; Hildebrand, J.G. Floral CO2 reveals flower profitability to moths. J. Chem. Ecol. 2004, 30, 1285–1288. [Google Scholar] [CrossRef]
- Xu, W.; Anderson, A. Carbon dioxide receptor genes in cotton bollworm Helicoverpa armigera. Naturwissenschaften 2015, 102, 11. [Google Scholar] [CrossRef]
- Senthilan, P.R.; Piepenbrock, D.; Ovezmyradov, G.; Nadrowski, B.; Bechstedt, S.; Pauls, S.; Winkler, M.; Möbius, W.; Howard, J.; Göpfert, M.C. Drosophila auditory organ genes and genetic hearing defects. Cell 2012, 150, 1042–1054. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.Y.; Xu, W.; Dong, S.L.; Zhu, J.Y.; Xu, Y.X.; Anderson, A. Genome-wide analysis of ionotropic receptor gene repertoire in Lepidoptera with an emphasis on its functions of Helicoverpa armigera. Insect Biochem. Mol. Biol. 2018, 99, 37–53. [Google Scholar] [CrossRef] [Green Version]
- Walker III, W.B.; Gonzalez, F.; Garczynski, S.F.; Witzgall, P. The chemosensory receptors of codling moth Cydia pomonella-expression in larvae and adults. Sci. Rep. 2016, 6, 23518. [Google Scholar] [CrossRef] [Green Version]
- Yuvaraj, J.K.; Andersson, M.N.; Zhang, D.D.; Löfstedt, C. Antennal transcriptome analysis of the chemosensory gene families from Trichoptera and basal Lepidoptera. Front. Physiol. 2018, 9, 1365. [Google Scholar] [CrossRef]
- Croset, V.; Rytz, R.; Cummins, S.F.; Budd, A.; Brawand, D.; Kaessmann, H.; Gibson, T.J.; Benton, R. Ancient protostome origin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction. PLoS Genet. 2010, 6, e1001064. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
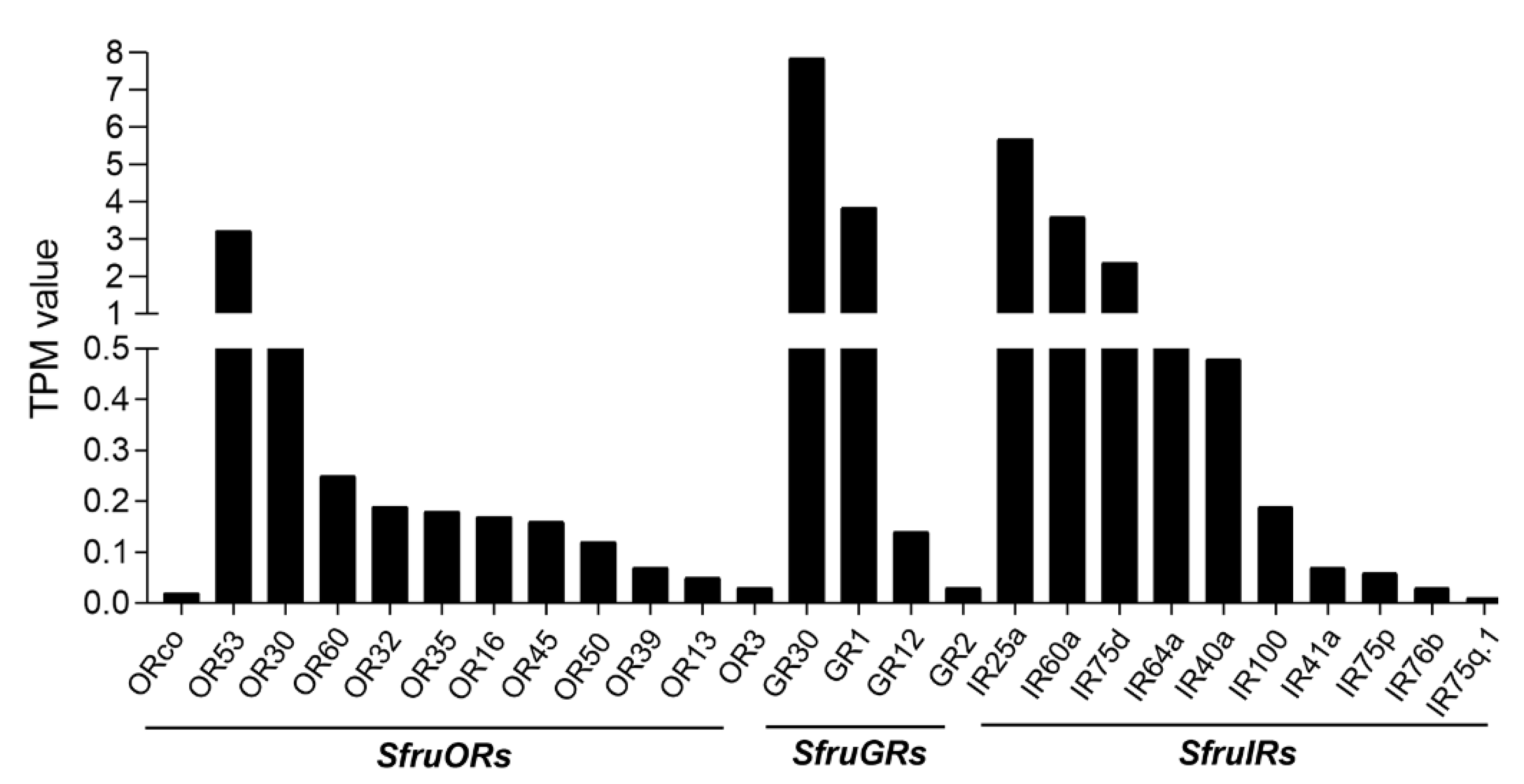
| Name | ID | ORF (aa) | TPM | BLASTx Best Hit (GenBank Accession/Name/Species) | Full Length | Identity (%) | E-Value |
|---|---|---|---|---|---|---|---|
| ORs | |||||||
| SfruORco | DN43518_c2_g2 | 473 | 0.01 | AAW52583.1|putative chemosensory receptor 2 [Spodoptera exigua] | Yes | 99 | 2.4 × 10−258 |
| SfruOR53 | DN34590_c0_g2 | 404 | 3.23 | ALM26238.1|odorant receptor 53 [Athetis dissimilis] | Yes | 78 | 1.0 × 10−187 |
| SfruOR30 | DN44157_c8_g3 | 387 | 0.55 | ALM26205.1|odorant receptor 16 [Athetis dissimilis] | Yes | 80 | 1.0 × 10−173 |
| SfruOR60 | DN36386_c0_g1 | 392 | 0.25 | ABQ82137.1|chemosensory receptor 2 [Spodoptera littoralis] | Yes | 98 | 0.0 |
| SfruOR32 | DN41457_c1_g1 | 397 | 0.19 | QEY02574.1|odorant receptor 5 [Spodoptera littoralis] | Yes | 72 | 0.0 |
| SfruOR35 | DN38262_c0_g2 | 453 | 0.18 | XP_022831643.1|odorant receptor 85c-like [Spodoptera litura] | Yes | 95 | 0.0 |
| SfruOR16 | DN42787_c4_g1 | 432 | 0.17 | ACL81182.1|putative olfactory receptor 16 [Spodoptera littoralis] | Yes | 93 | 0.0 |
| SfruOR45 | DN39388_c1_g1 | 429 | 0.16 | XP_022825109.1|odorant receptor 13a-like isoform X1 [Spodoptera litura] | Yes | 94 | 0.0 |
| SfruOR50 | DN34812_c1_g1 | 404 | 0.12 | QNS36220.1|olfactory receptor 23 [Mythimna separata] | Yes | 72 | 0.0 |
| SfruOR39 | DN33131_c1_g3 | 385 | 0.07 | QNS36227.1|olfactory receptor 36 [Mythimna separata] | Yes | 82 | 0.0 |
| SfruOR13 | DN37955_c1_g4 | 435 | 0.05 | AGI96750.1|olfactory receptor 13 [Spodoptera litura] | Yes | 90 | 0.0 |
| SfruOR3 | DN36417_c0_g1 | 379 | 0.03 | XP_022827581.1|odorant receptor 30a-like [Spodoptera litura] | Yes | 89 | 0.0 |
| GRs | |||||||
| SfruGR30 | DN41240_c0_g1 | 357 | 7.85 | QHB15310.1|gustatory receptor 10 [Peridroma saucia] | Yes | 92 | 0.0 |
| SfruGR1 | DN33391_c0_g2 | 464 | 3.85 | XP_022828173.1|gustatory and odorant receptor 22 [Spodoptera litura] | Yes | 99 | 0.0 |
| SfruGR12 | DN30079_c1_g1 | 396 | 0.14 | XP_022826955.1|gustatory receptor for sugar taste 64f-like [Spodoptera litura] | Yes | 93 | 0.0 |
| SfruGR2 | DN28721_c0_g1 | 413 | 0.03 | XP_022814066.1|gustatory and odorant receptor 22-like [Spodoptera litura] | Yes | 98 | 0.0 |
| IRs | |||||||
| SfruIR25a | DN42711_c2_g3 | 918 | 5.69 | XP_022828195.1|ionotropic receptor 25a [Spodoptera litura] | Yes | 99 | 0.0 |
| SfruIR60a | DN43581_c1_g1 | 585 | 3.59 | QHB15321.1|ionotropic receptor 60a [Peridroma saucia] | Yes | 80 | 0.0 |
| SfruIR75d | DN36075_c1_g1 | 314 | 2.37 | ALM24944.1|ionotropic receptor 75d [Athetis dissimilis] | No | 73 | 1.0 × 10−29 |
| SfruIR64a | DN38775_c1_g1 | 603 | 0.94 | ARB05666.1|ionization receptor 64a [Mythimna separata] | Yes | 80 | 0.0 |
| SfruIR40a | DN39774_c1_g2 | 712 | 0.48 | XP_022834254.1|ionotropic receptor 40a [Spodoptera litura] | Yes | 97 | 0.0 |
| SfruIR100 | DN42291_c0_g1 | 586 | 0.19 | QHB15322.1|ionotropic receptor 60a1b [Peridroma saucia] | Yes | 60 | 0.0 |
| SfruIR41a | DN41275_c0_g1 | 537 | 0.07 | ADR64681.1|putative chemosensory ionotropic receptor IR41a [Spodoptera littoralis] | No | 88 | 0.0 |
| SfruIR75p | DN38967_c0_g1 | 764 | 0.06 | XP_022816386.1|glutamate receptor 1-like [Spodoptera litura] | Yes | 90 | 0.0 |
| SfruIR76b | DN42735_c2_g1 | 542 | 0.03 | ADR64687.1|putative chemosensory ionotropic receptor IR76b [Spodoptera littoralis] | Yes | 95 | 0.0 |
| SfruIR75q.1 | DN44064_c7_g1 | 662 | 0.01 | ADR64686.1|putative chemosensory ionotropic receptor IR75q.1[Spodoptera littoralis] | Yes | 78 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.-L.; Dong, J.-F.; Yang, H.-B.; Li, D.-X.; Tian, C.-H. Identification and Characterization of Chemosensory Receptors in the Pheromone Gland-Ovipositor of Spodoptera frugiperda (J. E. Smith). Insects 2022, 13, 481. https://doi.org/10.3390/insects13050481
Sun Y-L, Dong J-F, Yang H-B, Li D-X, Tian C-H. Identification and Characterization of Chemosensory Receptors in the Pheromone Gland-Ovipositor of Spodoptera frugiperda (J. E. Smith). Insects. 2022; 13(5):481. https://doi.org/10.3390/insects13050481
Chicago/Turabian StyleSun, Ya-Lan, Jun-Feng Dong, Hai-Bo Yang, Ding-Xu Li, and Cai-Hong Tian. 2022. "Identification and Characterization of Chemosensory Receptors in the Pheromone Gland-Ovipositor of Spodoptera frugiperda (J. E. Smith)" Insects 13, no. 5: 481. https://doi.org/10.3390/insects13050481
APA StyleSun, Y. -L., Dong, J. -F., Yang, H. -B., Li, D. -X., & Tian, C. -H. (2022). Identification and Characterization of Chemosensory Receptors in the Pheromone Gland-Ovipositor of Spodoptera frugiperda (J. E. Smith). Insects, 13(5), 481. https://doi.org/10.3390/insects13050481