

Predicting Culex pipiens/restuans Population Dynamics Using a Weather-Driven Dynamic Compartmental Population Model



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
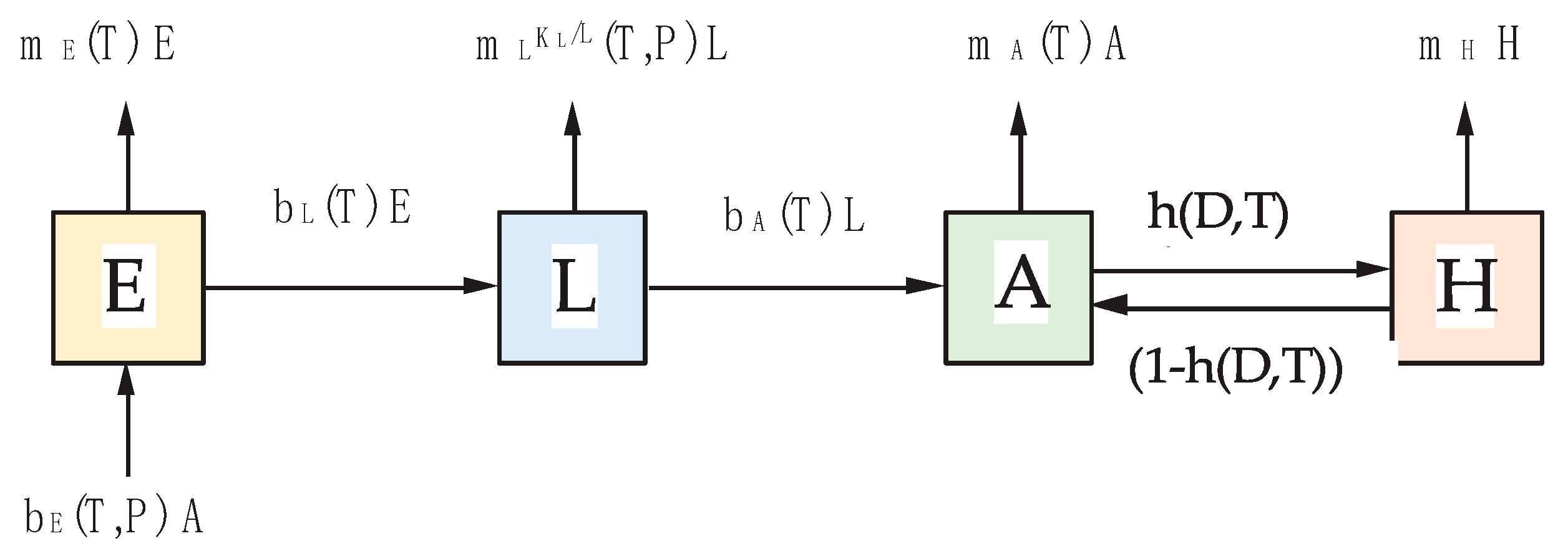
2.1. Cx. pipiens/restuans Population Model
2.1.1. Egg Stage—E
2.1.2. Larval Stage—L
2.1.3. Adult Stages—A and H
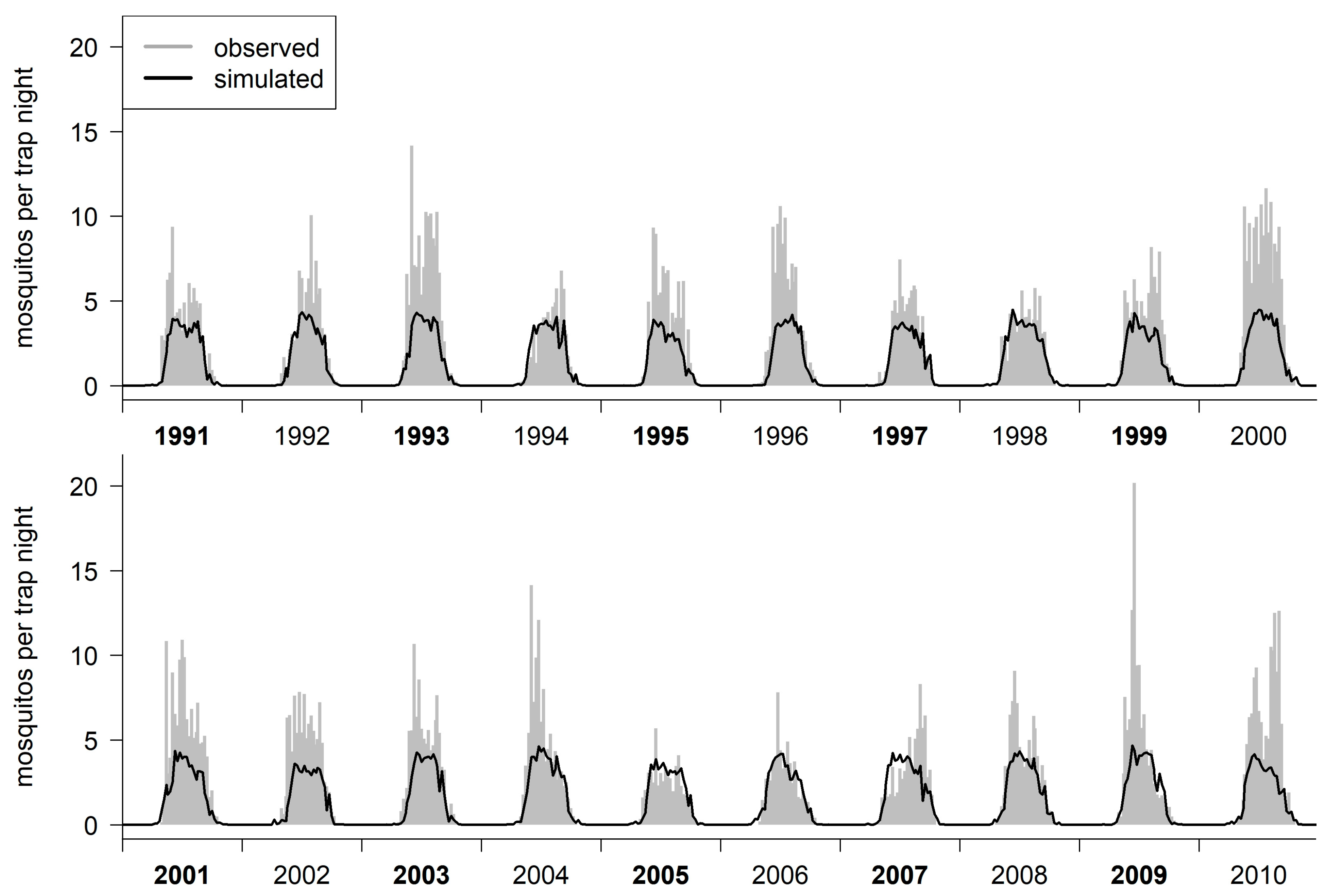
2.2. Model Evaluation
2.2.1. Mosquito Count Data
2.2.2. Environmental Data
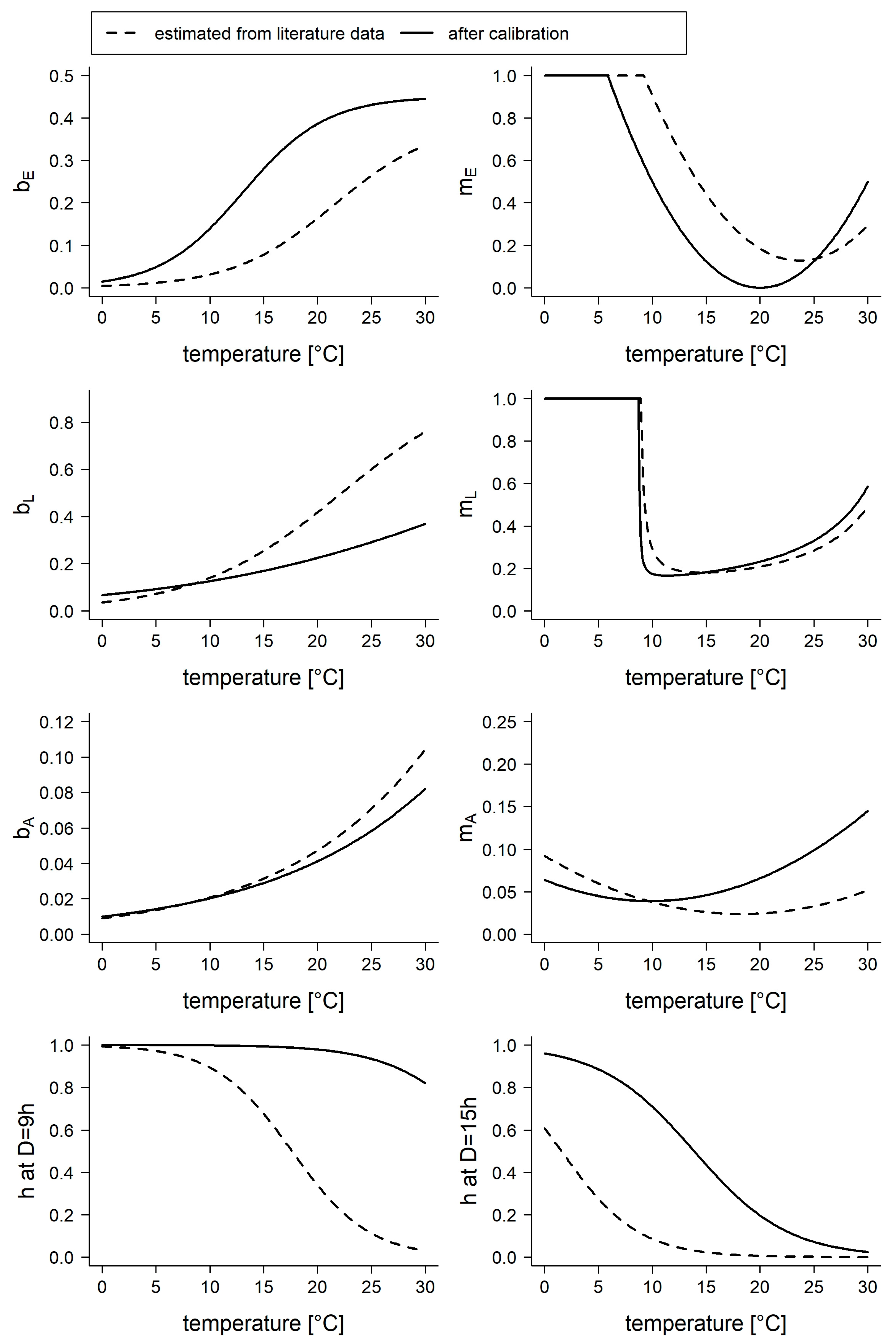
2.2.3. Model Calibration and Validation
2.2.4. Estimating the Effect of Mosquito Control Strategies
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calisher, C.H. Medically important arboviruses of the United States and Canada. Clin. Microbiol. Rev. 1994, 7, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Dobler, G.; Aspöck, H. Krank Durch Arthropoden. Denisia 30., Ch. Durch Stechmücken übertragene Arboviren als Erreger von Infektionskrakheiten des Menschen; Land Oberosterreich, Biologiezentrum/Oberösterreichische Landesmuseen: Linz, Austria, 2010; pp. 501–553. [Google Scholar]
- Farajollahi, A.; Fonseca, D.M.; Kramer, L.D.; Marm Kilpatrick, A. ”Bird biting” mosquitoes and human disease: A review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect. Genet. Evol. 2011, 11, 1577–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, L.; Ebel, G. Dynamics of flavivirus infection in mosquitoes. Adv. Virus Res. 2003, 60, 187–232. [Google Scholar] [PubMed]
- Hayes, E.B.; Komar, N.; Nasci, R.S.; Montgomery, S.P.; O’Leary, D.R.; Campbell, G.L. Epidemiology and transmission dynamics of West Nile virus disease. Emerg. Infect. Dis. 2005, 11, 1167–1173. [Google Scholar] [CrossRef]
- Barr, A.R. The distribution of Culex p. pipiens and C. p. quinquefasciatus in North America. Am. J. Trop. Med. 1957, 6, 153–165. [Google Scholar] [CrossRef]
- Darsie, R.F.; Ward, R.A. Identification and Geographical Distribution of the Mosquitoes of North America, North of Mexico; University Press of Florida: Gainesville, FL, USA, 2004. [Google Scholar]
- Bolling, B.G.; Kennedy, J.H.; Zimmerman, E.G. Seasonal dynamics of four potential West Nile vector species in north-central texas. J. Vector Ecol. 2005, 30, 186–194. [Google Scholar]
- Chuang, T.W.; Hildreth, M.B.; Vanroekel, D.L.; Wimberly, M.C. Weather and land cover influences on mosquito populations in Sioux Falls, South Dakota. J. Med. Entomol. 2011, 48, 669–679. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ogden, N.H.; Zhu, H. The impact of weather conditions on Culex pipiens and Culex restuans (Diptera: Culicidae) Abundance: A case study in Peel Region. J. Med. Entomol. 2011, 48, 468–475. [Google Scholar] [CrossRef]
- Madder, D.J.; Surgeoner, G.A.; Helson, B.V. Number of generations, egg production, and developmental time of Culex pipiens and Culex restauns (Diptera: Culicidae) in southern Ontario. J. Med. Entomol. 1983, 20, 275–287. [Google Scholar] [CrossRef]
- Reisen, W.K. Effect of Temperature on Culex tarsalis (Diptera: Culicidae) from the Coachella and San Joaquin Valleys of California. J. Med. Entomol. 1995, 32, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Rueda, L.M.; Patel, K.J.; Axtell, R.C.; Stinner, R.E. Temperature-dependent development and survival rates of Culex quinquefasciatus and Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 1990, 27, 892–898. [Google Scholar] [CrossRef]
- Suman, D.S.; Tikar, S.N.; Mendki, M.J.; Sukumaran, D.; Agrawal, O.P.; Parashar, B.D.; Prakash, S. Variations in life tables of geographically isolated strains of the mosquito Culex quinquefasciatus. Med. Vet. Entomol. 2011, 525, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Vinogradova, E.B. Mosquitoes Culex pipiens Pipiens: Taxonomy, Distribution, Ecology, Physiology, Genetics and Control; PenSoft: Sofia, Bulgaria, 2000; 280p. [Google Scholar]
- Spielman, A.; Wong, J. Environmental control of ovarian diapause in Culex pipiens. Ann. Entomol. Soc. Am. 1973, 66, 905–907. [Google Scholar] [CrossRef]
- Wilton, D.P.; Smith, G.C. Ovarian diapause in three geographic strains of Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 1985, 22, 524–528. [Google Scholar] [CrossRef]
- Pecoraro, H.L.; Day, H.L.; Reineke, R.; Stevens, N.; Withey, J.C.; Marzluff, J.M.; Meschke, J.S. Climatic and landscape correlates for potential West Nile virus mosquito vectors in the Seattle region. J. Vector Ecol. 2007, 32, 22–28. [Google Scholar] [CrossRef]
- Trawinski, P.R.; MacKay, D.S. Identification of environmental covariates of West Nile virus vector mosquito population abundance. Vector Borne Zoonotic Dis. 2010, 10, 515–526. [Google Scholar] [CrossRef]
- Diuk-Wasser, M.A.; Brown, H.D.; Andreadis, T.G.; Fish, D. Modeling the spatial distribution of mosquito vectors West Nile virus in Connecticut, USA. Vector Borne Zoonotic Dis. 2006, 6, 283–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trawinski, P.R.; MacKay, D.S. Meteorologically conditioned time-series predictions of West Nile virus vector mosquitoes. Vector Borne Zoonotic Dis. 2008, 8, 505–521. [Google Scholar] [CrossRef] [Green Version]
- Lebl, K.; Brugger, K.; Rubel, F. Predicting Culex pipiens/restuans population dynamics by interval lagged weather data. Parasites Vectors 2013, 6, 129. [Google Scholar] [CrossRef] [Green Version]
- Ahumada, J.A.; Lapointe, D.; Samuel, M.D. Modeling the population dynamics of Culex quinquefasciatus (Diptera: Culicidae), along an elevational gradient in Hawaii. J. Med. Entomol. 2004, 41, 1157–1170. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, J.N.; Reisen, W.; Spear, R.C. Dynamic Model Comparing the Binoimics of Two Isolated Culex tarsalis Populations: Model Development. J. Med. Entomol. 1995, 32, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.W.; Comrie, A.C. Modeled response of the West Nile virus vector Culex quinquefasciatus to changing climate using the dynamic mosquito simulation model. Int. J. Biometeorol. 2010, 54, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Laperriere, V.; Brugger, K.; Rubel, F. Simulation of the seasonal cycles of bird, equine and human West Nile virus cases. Prev. Vet. Med. 2011, 98, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Rubel, F.; Brugger, K.; Hantel, M.; Chvala-Mannsberger, S.; Bakonyi, T.; Weissenböck, H.; Nowotny, N. Explaining Usutu virus dynamics in Austria: Model development and calibration. Prev. Vet. Med. 2008, 85, 166–186. [Google Scholar] [CrossRef]
- Wonham, M.J.; de Camino-Beck, T.; Lewis, M.A. An epidemiological model for West Nile virus: Invasion analysis and control applications. Proc. Royal Soc. B 2004, 271, 501–507. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Madras, N.; Zhu, H. Temperature-driven population abundance model for Culex pipiens and Culex restuans (Diptera: Culicidae). J. Theor. Biol. 2018, 443, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Ballenger-Browning, K.; Elder, J. Multi-modal aedes aegypti mosquito reduction interventions and dengue fever prevention. Trop. Med. Int. Health. 2009, 14, 1542–1551. [Google Scholar] [CrossRef] [PubMed]
- Alphey, L.; Benedict, M.; Bellini, R.; Clark, G.; Dame, D.; Service, M.; Dobson, S. Sterile-insect methods for control of mosquito-borne diseases: An analysis. Vector Borne Zoonotic Dis. 2010, 10, 295–311. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing, Version 4.1.2; R Foundation for Statistical Computing: Vienna, Austria, 2022.
- Elizondo-Quiroga, A.; Flores-Suarez, A.; Elizondo-Quiroga, D.; Ponce-Garcia, G.; Blitvich, B.J.; Contreras-Cordero, J.F.; Gonzalez-Rojas, J.I.; Mercado-Hernandez, R.; Beaty, B.J.; Fernandez-Salas, I. Gonotrophic cycle and survivorship of Culex quinquefasciatus (Diptera: Culicidae) using sticky ovitraps in Monterrey, northeastern Mexico. J. Am. Mosq. Control Assoc. 2006, 22, 10–14. [Google Scholar]
- Reisen, W.K.; Milby, M.M. Studies on autogeny in Culex tarsalis: 3. Life table attributes of autogenous and anautogenous strains under laboratory conditions. J. Am. Mosq. Control Assoc. 1987, 3, 619–625. [Google Scholar]
- Oda, T.; Ueda, M. Seasonal changes in the oviposition activity of Culex pipiens molestus Forskal in the above-ground water. Trop. Med. 1979, 21, 139–144. [Google Scholar]
- Oda, T.; Mori, A.; Ueda, M.; Kurokawa, K. Effects of temperatures on the oviposition and hatching of eggs in Culex pipiens molestus and Culex pipiens quinquefasciatus. Trop. Med. 1980, 22, 167–172. [Google Scholar]
- Shriver, D.; Bickley, W.E. The effect of temperature on hatching eggs of the mosquito, culex pipiens quinquefasciatus say. Mosquito News 1964, 24, 137–140. [Google Scholar]
- Almir’on, W.R.; Brewer, M.E. Winter biology of Culex pipiens quinquefasciatus Say, (Diptera: Culicidae) from Cord´oba, Argentina. Mem. Do Inst. Oswaldo Cruz 1996, 91, 649–654. [Google Scholar] [CrossRef] [Green Version]
- Kirkpatrick, T.W. The Mosquitoes of Egypt; Government Press: Cairo, Egypt, 1925; p. 224.
- Kramer, S.D. The effect of temperature on the life cycle of Musca domestica and Culex pipiens. Science 1915, 41, 874–877. [Google Scholar] [CrossRef] [PubMed]
- Olejníček, J.; Gelbič, I. Differences in response to temperature and density between two strains of the mosquito, Culex pipiens molestus Forskal. J. Vector Ecol. 2000, 25, 136–145. [Google Scholar]
- Tamarina, N.A. The developmental cycle of culex pipiens molestus forsk. Vestn. MGU (Biol.) 1966, 2, 65–75. [Google Scholar]
- Bailey, S.F.; Gieke, P.A. A study of the effect of water temperatures on rice field mosquito development. In Proceedings and Papers of the Annual Conference of the California Mosquito Control Association; CMCA Press: Rockland, ME, USA, 1968; pp. 53–61. [Google Scholar]
- Mogi, M. Temperature and photoperiod effects on larval and ovarian development of New Zealand strains of Culex quinquefasciatus (Diptera: Culicidae). Ann. Entomol. Soc. Am. 1992, 85, 58–66. [Google Scholar] [CrossRef]
- Mori, A.; Oda, T.; Zaitsu, M.; Ueda, M.; Kurokawa, K. Studies on the developing period of larval stage of the Culex pipiens complex in Japan. Trop. Med. 1988, 30, 155–161. [Google Scholar]
- Shelton, R.M. The effect of temperatures on development of eight mosquito species. Mosq. News 1973, 33, 1–12. [Google Scholar]
- Reisen, W.K.; Meyer, R.P.; Shields, J.; Arbolante, C. Population ecology of preimaginal Culex tarsalis (Diptera: Culicidae) in Kern County, California. J. Med. Entomol. 1989, 26, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Buth, J.L.; Brust RAEllis, R.A. Development time, oviposition activity and onset of diapause in Culex tarsalis, Culex restuans and Culiseta inornata in southern Manitoba. J. Am. Mosq. Control Assoc. 1990, 6, 55–63. [Google Scholar] [PubMed]
- Kalchenko, E.I. The biology of Culex pipiens molestus Forsk. (Diptera, Culicidae). Entomol. Obozr. 1962, 41, 94–98. [Google Scholar]
- Gunay, F.; Alten, B.; Ozsoy, E.D. Estimating reaction norms for predictive population parameters, age specific mortality, and mean longevity in temperature-dependent cohorts of Culex quinquefasciatus Say (Diptera: Culicidae). J. Vector Ecol. 2010, 35, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Eshita, Y.; Uchida, K.; Mine, M.; Kurokawa, K.; Ogawa, Y.; Tahara, H.K. Reproductive activity and survival of Culex pipiens pallens and Culex quinquefasciatus (Diptera: Culicidae) in Japan at high temperature. J. Med. Entomol. 2002, 39, 185–190. [Google Scholar] [CrossRef]
- Eldridge, B.F. Environmental Control of Ovarian Development in Mosquitoes of the Culex pipiens complex. Science 1966, 151, 826–828. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.M.; Faran, T.; Gargan, I.I.; Hayes, D. Winter survival of blood-fed and nonblood-fed Culex pipiens L. Am. J. Trop. Med. 1982, 31, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- NCDC, 2012: NNCD Climate Data Online. Available online: https://www.ncei.noaa.gov/cdo-web/ (accessed on 1 December 2022).
- Forsythe, W.C.; Rykiel, E.J.; Stahl, R.S.; Wu, H.; Schoolfield, R.M. A model comparison for daylength as a function of latitude and day of year. Ecol Modell. 1995, 80, 87–95. [Google Scholar] [CrossRef]
- Katoch, S.; Chauhan, S.S.; Kumar, V. A review on genetic algorithm: Past, present, and future. Multimed. Tools Appl. 2021, 80, 8091–8126. [Google Scholar] [CrossRef]
- Willighagen, E. Genalg: R Based Genetic Algorithm; version 0.1.1; 2005. Available online: http://cran.rproject.org/web/packages/genalg/index.html (accessed on 27 February 2023).
- Erguler, K.; Mendel, J.; Petrić, D.V.; Petrić, M.; Kavran, M.; Demirok, M.C.; Gunay, F.; Georgiades, P.; Alten, B.; Lelieveld, J. A dynamically structured matrix population model for insect life histories observed under variable environmental conditions. Sci. Rep. 2022, 12, 11587. [Google Scholar] [CrossRef]
- Becker, N.; Petrić, D.; Zgomba, M. Mosquitoes-Identification, Ecology and Control, 3rd ed.; Springer: Cham, Switzerland, 2020; p. 570. [Google Scholar]
- Valdez, L.; Sibona, G.; Díaz LContigiani, M.; Condat, C. Effects of rainfall on Culex mosquito population dynamics. J. Theor. Biol. 2017, 421, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, L.; Styer, L.M.; Ebel, G.A. Global perspective on the epidemiology of the west nile virus. Annu. Rev. Entomol. 2008, 53, 61–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinior, B.; Lebl, K.; Firth, C.; Rubel, F.; Fuchs, R.; Stockreiter, S.; Loitsch, A.; Köfer, J. Cost analysis of bluetongue virus serotype 8 surveillance and vaccination programmes in Austria from 2005 to 2013. Vet. J. 2015, 206, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Pinior, B.; Brugger, K.; Köfer, J.; Schwermer, H.; Stockreiter, S.; Loitsch, A.; Rubel, F. Economic comparison of the monitoring programmes for bluetongue vectors in Austria and Switzerland. Vet. Rec. 2015, 176, 464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zittra, C.; Flechl, E.; Kothmayer, M.; Vitecek, S.; Rossiter, H.; Zechmeister, T.; Fuehrer, H.-P. Ecological characterization and molecular differentiation of Culex pipiens complex taxa and Culex torrentium in eastern Austria. Parasites Vectors 2016, 9, 197. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter 1 | Description |
|---|---|
| bE (T, P) | birth rate eggs—reciprocal value of the length of the gonotrophic cycle multiplied with the number of female eggs laid per egg raft |
| mE (T) | mortality rate of eggs |
| bL (T) | birth rate larvae—reciprocal value of the length of the egg stag |
| mL (T, KL) | mortality rate of larvae and pupae combined |
| KL (P) | carrying capacity of the larvae |
| bA (T) | birth rate adults—reciprocal value of the length of the larval and pupal stage combined |
| mA (T) | mortality rate of active adult mosquitoes |
| h (T, D) | diapausing rate—rate of adult mosquito leaving the active stage and entering the diapause stage |
| mH | mortality rate of adult mosquitoes during winter diapause |
| Func. | Par. | Estimate | Lower | Upper | Calib | Min | Max | Mean | SD | RSD |
|---|---|---|---|---|---|---|---|---|---|---|
| bE | a | 3.9 × 10−1 | 3.4 × 10−1 | 4.5 × 10−1 | 4.5 × 10−1 | 4.2 × 10−1 | 4.5 × 10−1 | 4.4 × 10−1 | 7.9 × 10−3 | 1.8 × 10−2 |
| b | 9.5 × 101 | 2.9 × 101 | 1.6 × 102 | 3.0 × 101 | 2.9 × 101 | 3.2 × 101 | 3.0 × 101 | 8.6 × 10−1 | 2.9 × 10−2 | |
| c | −2.1 × 10−1 | −2.6 × 10−1 | −1.7 × 10−1 | −2.6 × 10−1 | −2.6 × 10−1 | −2.4 × 10−1 | −2.5 × 10−1 | 5.3 × 10−3 | −2.1 × 10−2 | |
| d * | 0.0 × 100 | 0.0 × 100 | 1.0 × 10−1 | 1.0 × 10−1 | 5.0 × 10−2 | 1.0 × 10−1 | 9.7 × 10−2 | 9.1 × 10−3 | 9.4 × 10−2 | |
| bE, KL | k * | 0.0 × 100 | 0.0 × 100 | 2.0 × 101 | 1.1 × 100 | 8.4 × 10−1 | 1.8 × 100 | 1.2 × 100 | 2.5 × 10−1 | 2.1 × 10−1 |
| mE | a | 2.5 × 100 | 2.0 × 100 | 2.9 × 100 | 2.0 × 100 | 2.0 × 100 | 2.5 × 100 | 2.1 × 100 | 1.1 × 10−1 | 5.4 × 10−2 |
| b | −2.0 × 10−1 | −2.4 × 10−1 | −1.6 × 10−1 | −2.0 × 10−1 | −2.2 × 10−1 | −1.8 × 10−1 | −2.0 × 10−1 | 1.1 × 10−2 | −5.6 × 10−2 | |
| c | 4.2 × 10−3 | 3.3 × 10−3 | 5.0 × 10−3 | 5.0 × 10−3 | 4.1 × 10−3 | 5.0 × 10−3 | 4.9 × 10−3 | 2.4 × 10−4 | 5.0 × 10−2 | |
| bL | a | 2.7 × 101 | 1.4 × 101 | 4.0 × 101 | 1.4 × 101 | 1.3 × 101 | 3.3 × 101 | 1.6 × 101 | 4.2 × 100 | 2.7 × 10−1 |
| b | −1.5 × 10−1 | −1.7 × 10−1 | −1.3 × 10−1 | −7.0 × 10−2 | −1.7 × 10−1 | −5.9 × 10−2 | −1.2 × 10−1 | 3.2 × 10−2 | −2.7 × 10−1 | |
| mL | a | 1.9 × 101 | −3.6 × 100 | 4.1 × 101 | 3.1 × 101 | 2.5 × 101 | 3.3 × 101 | 2.8 × 101 | 2.3 × 100 | 8.1 × 10−2 |
| b | −2.9 × 100 | −3.8 × 100 | −2.0 × 100 | −3.7 × 100 | −3.8 × 100 | −3.7 × 100 | −3.7 × 100 | 4.1 × 10−2 | −1.1 × 10−2 | |
| KL | a * | 1.0 × 100 | 1.0 × 100 | 5.0 × 101 | 1.0 × 100 | 1.0 × 100 | 1.8 × 100 | 1.2 × 100 | 2.2 × 10−1 | 1.8 × 10−1 |
| b * | 1.0 × 100 | 1.0 × 100 | 7.0 × 101 | 3.6 × 101 | 3.6 × 101 | 3.7 × 101 | 3.6 × 101 | 5.7 × 10−1 | 1.6 × 10−2 | |
| bA | a | 1.1 × 102 | 7.3 × 101 | 1.5 × 102 | 1.0 × 102 | 7.5 × 101 | 1.5 × 102 | 1.1 × 102 | 2.2 × 101 | 2.1 × 10−1 |
| b | −8.5 × 10−2 | −9.8 × 10−2 | −7.3 × 10−2 | −7.3 × 10−2 | −7.4 × 10−2 | −7.3 × 10−2 | −7.3 × 10−2 | 3.9 × 10−4 | −5.2 × 10−3 | |
| mA | a | 9.2 × 10−2 | 6.4 × 10−2 | 1.2 × 10−1 | 6.4 × 10−2 | 6.4 × 10−2 | 8.5 × 10−2 | 6.8 × 10−2 | 5.0 × 10−3 | 7.3 × 10−2 |
| b | −7.5 × 10−3 | −1.0 × 10−2 | −5.0 × 10−3 | −5.1 × 10−3 | −5.3 × 10−3 | −4.9 × 10−3 | −5.0 × 10−3 | 1.0 × 10−4 | −2.1 × 10−2 | |
| c | 2.0 × 10−4 | 1.5 × 10−4 | 2.6 × 10−4 | 2.6 × 10−4 | 2.6 × 10−4 | 2.6 × 10−4 | 2.6 × 10−4 | 8.7 × 10−7 | 3.4 × 10−3 | |
| mH | a | 1.8 × 10−2 | 1.7 × 10−2 | 2.0 × 10−2 | 1.7 × 10−2 | 1.7 × 10−2 | 2.0 × 10−2 | 1.8 × 10−2 | 8.1 × 10−4 | 4.5 × 10−2 |
| h | a | 8.4 × 10−6 | −6.7 × 10−6 | 2.4 × 10−5 | 8.8 × 10−8 | 8.0 × 10−8 | 3.2 × 10−7 | 1.2 × 10−7 | 5.6 × 10−8 | 4.6 × 10−1 |
| h | b | 2.8 × 10−1 | 2.3 × 10−1 | 3.3 × 10−1 | 2.3 × 10−1 | 2.3 × 10−1 | 2.4 × 10−1 | 2.4 × 10−1 | 1.9 × 10−3 | 8.0 × 10−3 |
| h | c | 7.5 × 10−1 | 6.2 × 10−1 | 8.7 × 10−1 | 8.7 × 10−1 | 7.8 × 10−1 | 8.7 × 10−1 | 8.5 × 10−1 | 2.4 × 10−2 | 2.9 × 10−2 |
| 5% | 10% | 25% | 50% | 75% | |
|---|---|---|---|---|---|
| decrease bE | 99.6 | 99.2 | 97.8 | 94.2 | 86.5 |
| increase mE | 99.7 | 99.5 | 98.6 | 97.4 | 96.3 |
| increase mL | 96.2 | 92.6 | 82.6 | 68.4 | 56.5 |
| increase mA | 96.2 | 92.7 | 83.6 | 71.9 | 63.1 |
| increase mH | 99.1 | 98.3 | 95.9 | 92.3 | 89.1 |
| decrease bE + increase mE | 99.3 | 98.7 | 96.4 | 91.4 | 82.2 |
| decrease bE + increase mL | 95.8 | 91.8 | 80.7 | 64.2 | 48.0 |
| decrease bE + increase mA | 95.8 | 92.0 | 81.6 | 67.0 | 52.6 |
| decrease bE + increase mH | 98.7 | 97.5 | 93.6 | 85.9 | 73.8 |
| increase mE + increase mL | 95.9 | 92.1 | 81.5 | 66.5 | 54.2 |
| increase mE + increase mA | 95.9 | 92.2 | 82.4 | 69.8 | 60.4 |
| increase mE + increase mH | 98.8 | 97.7 | 94.6 | 89.8 | 85.6 |
| increase mL + increase mA | 92.6 | 85.8 | 69.1 | 49.3 | 35.8 |
| increase mL + increase mH | 95.3 | 91.0 | 79.1 | 62.9 | 49.7 |
| increase mA + increase mH | 95.4 | 91.1 | 80.4 | 66.8 | 56.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakran-Lebl, K.; Kjær, L.J.; Conrady, B. Predicting Culex pipiens/restuans Population Dynamics Using a Weather-Driven Dynamic Compartmental Population Model. Insects 2023, 14, 293. https://doi.org/10.3390/insects14030293
Bakran-Lebl K, Kjær LJ, Conrady B. Predicting Culex pipiens/restuans Population Dynamics Using a Weather-Driven Dynamic Compartmental Population Model. Insects. 2023; 14(3):293. https://doi.org/10.3390/insects14030293
Chicago/Turabian StyleBakran-Lebl, Karin, Lene Jung Kjær, and Beate Conrady. 2023. "Predicting Culex pipiens/restuans Population Dynamics Using a Weather-Driven Dynamic Compartmental Population Model" Insects 14, no. 3: 293. https://doi.org/10.3390/insects14030293
APA StyleBakran-Lebl, K., Kjær, L. J., & Conrady, B. (2023). Predicting Culex pipiens/restuans Population Dynamics Using a Weather-Driven Dynamic Compartmental Population Model. Insects, 14(3), 293. https://doi.org/10.3390/insects14030293