

The Adaptive Value of Chromosomal Inversions and Climatic Change—Studies on the Natural Populations of Drosophila subobscura from the Balkans


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Drosophila subobscura Sampling and Chromosome Preparation
2.2. Climatic Information
2.3. Statistical Analyses
3. Results
3.1. The Chromosomal Inversion Polymorphism of the Petnica Population
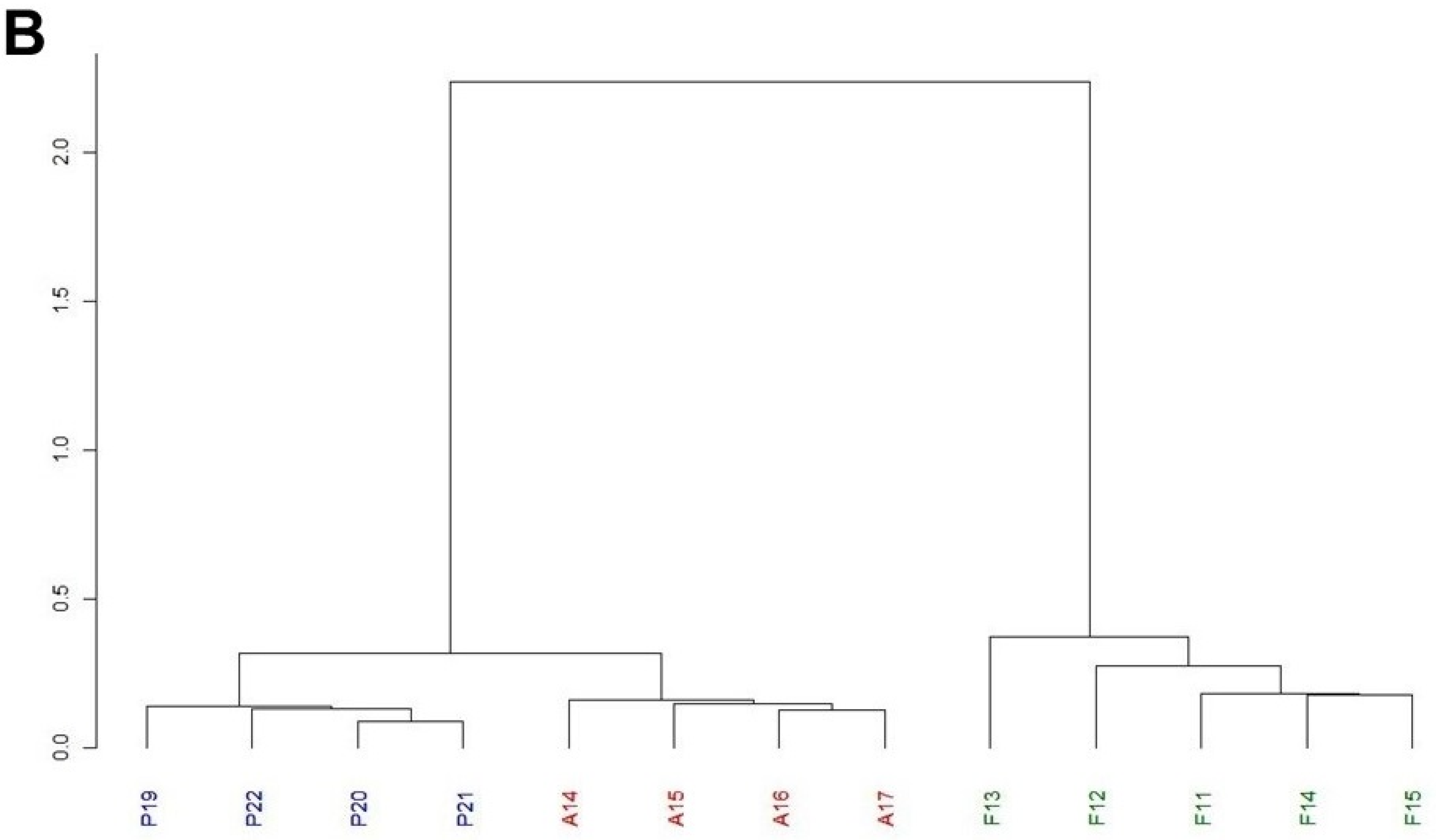
3.2. Comparison between the Inversion Chromosomal Polymorphism of Petnica, Avala, and Font Groga
3.3. O Chromosome Inversion Polymorphism from the Balkan Populations
3.4. Relationship between Chromosomal Inversions and Meteorological Variables
3.5. Changes over Time in the Chromosomal Polymorphism from Petnica
3.6. CTI in the Balkan Populations of D. subobscura
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2021: The Physical Science Basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; Volume 3, pp. 154–196. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2022: Mitigation of Climate Change. Contribution of Working Group III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Shukla, P.R., Skea, J., Slade, R., Al Khourdajie, A., van Diemen, R., McCollum, D., Pathak, M., Some, S., Vyas, P., Fradera, R., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022. [Google Scholar] [CrossRef]
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Galetti, M.; Alamgir, M.; Crist, E.; Mahmoud, M.I.; Laurance, W.F. World scientists’ warning to humanity: A second notice. BioScience 2017, 67, 1026–1028. [Google Scholar] [CrossRef] [Green Version]
- Ripple, W.I.; Wolf, C.; Newsome, T.M.; Barnard, P.; Moomaw, W.R.; Grandcolas, P. World scientists’ warning of a climate emergency. BioScience 2020, 70, 8–12. [Google Scholar] [CrossRef]
- Ripple, W.J.; Wolf, C.; Gregg, J.W.; Levin, K.; Rockström, J.; Newsome, T.M.; Betts, M.G.; Huq, S.; Law, B.E.; Kemp, L.; et al. World scientists’ warning of a climate emergency 2022. BioScience 2022, 72, 1149–1155. [Google Scholar] [CrossRef]
- Georgian, S.; Hameed, S.; Morgan, L.; Amon, D.J.; Sumaila, U.R.; Johns, D.; Ripple, W.J. Scientists’ warning of an imperiled ocean. Biol. Conserv. 2022, 272, 109595. [Google Scholar] [CrossRef]
- Feeley, K.J.; Rehm, E.M.; Machovina, B. The responses of tropical forest species to global climate change: Acclimate, adapt, migrate or go extinct? Front. Biogeogr. 2012, 4, 69–84. [Google Scholar] [CrossRef] [Green Version]
- Darwin, C.R. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life; Murray: London, UK, 1859. [Google Scholar]
- Dobzhansky, T. Genetics of the Evolutionary Process; Columbia University Press: New York, NY, USA, 1970. [Google Scholar]
- Lewontin, R.C.; Moore, J.A.; Provine, W.B.; Wallace, B. Dobzhanky’s Genetics of Natural Populations I–XLIII; Columbia University Press: New York, NY, USA, 1981. [Google Scholar]
- Levitan, M. Climatic factors and increased frequencies of ‘southern’ chromosome forms in natural populations of Drosophila robusta. Evol. Ecol. Res. 2003, 5, 597–604. [Google Scholar]
- Levitan, M.; Etges, W.J. Climate change and recent genetic flux in populations of Drosophila robusta. BMC Evol. Biol. 2005, 5, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umina, P.A.; Weeks, A.R.; Kearney, M.R.; McKechnie, S.W.; Hoffmann, A.A. A rapid shift in a classic clinal pattern in Drosophila reflecting climate change. Science 2005, 308, 691–693. [Google Scholar] [CrossRef]
- Heerwaarden, B.V.; Hoffmann, A.A. Global warming: Fly populations are responding rapidly to climate change. Curr. Biol. 2007, 17, R16–R18. [Google Scholar] [CrossRef] [Green Version]
- Rane, R.V.; Rako, L.; Kapun, M.; Lee, S.F.; Hoffmann, A.A. Genomic evidence for role of inversion 3RP of Drosophila melanogaster in facilitating climate change adaptation. Mol. Ecol. 2015, 24, 2423–2432. [Google Scholar] [CrossRef]
- Krimbas, C.B. The Inversion Polymorphism of Drosophila subobscura. In Drosophila Inversion Polymorphism; Krimbas, C.B., Powell, J.R., Eds.; CRC Press: Boca Raton, FL, USA, 1992; pp. 127–220. [Google Scholar]
- Krimbas, C.B. Drosophila subobscura. Biology, Genetics and Inversion Polymorphism; Verlag Dr. Kovac: Hamburg, Germany, 1993. [Google Scholar]
- Krimbas, C.B.; Loukas, M. The inversion polymorphism of Drosophila subobscura. Evol. Ecol. 1980, 12, 163–234. [Google Scholar]
- Brncic, D.; Prevosti, A.; Budnik, M.; Monclus, M.; Ocaña, J. Colonization of Drosophila subobscura in Chile I. First population and cytogenetic studies. Genetica 1981, 56, 3–9. [Google Scholar]
- Beckenbach, A.T.; Prevosti, A. Colonization of North America by the European species, Drosophila subobscura and D. ambigua. Am. Midl. Natl. 1986, 115, 10–18. [Google Scholar] [CrossRef]
- Prevosti, A.; Ribo, G.; Serra, L.; Aguade, M.; Balaña, J.; Monclus, M.; Mestres, F. Colonization of America by Drosophila subobscura: Experiment in natural populations that supports the adaptive role of chromosomal-inversion polymorphism. Proc. Natl. Acad. Sci. USA 1988, 85, 5597–5600. [Google Scholar] [CrossRef] [PubMed]
- Menozzi, P.; Krimbas, C.B. The inversion polymorphism of D. subobscura revisited: Synthetic maps of gene arrangement frequencies and their interpretation. J. Evol. Biol. 1992, 5, 625–641. [Google Scholar] [CrossRef]
- Orengo, D.J.; Prevosti, A. Temporal changes in chromosomal polymorphism of Drosophila subobscura related to climatic changes. Evolution 1996, 50, 1346–1350. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.; Céspedes, W.; Balanya, J.; Trotta, V.; Calboli, F.C.; Fontdevila, A.; Serra, L. Temperature-related genetic changes in laboratory populations of Drosophila subobscura: Evidence against simple climatic-based explanations for latitudinal clines. Am. Nat. 2005, 165, 258–273. [Google Scholar] [CrossRef]
- Santos, M. Evolution of total net fitness in thermal lines: Drosophila subobscura likes it ‘warm’. J. Evol. Biol. 2007, 20, 2361–2370. [Google Scholar] [CrossRef] [PubMed]
- Rego, C.; Balanya, J.; Fragata, I.; Matos, M.; Rezende, E.L.; Santos, M. Clinal patterns of chromosomal inversion polymorphisms in Drosophila subobscura are partly associated with thermal preferences and heat stress resistance. Evolution 2010, 64, 385–397. [Google Scholar] [CrossRef]
- Dolgova, O.; Rego, C.; Calabria, G.; Balanyà, J.; Pascual, M.; Rezende, E.L.; Santos, M. Genetic constraints for thermal coadaptation in Drosophila subobscura. BMC Evol. Biol. 2010, 10, 363. [Google Scholar] [CrossRef] [Green Version]
- Calabria, G.; Dolgova, O.; Rego, C.; Castañeda, L.E.; Rezende, E.L.; Balanyà, J.; Pascual, M.; Sørensen, J.G.; Loeschcke, V.; Santos, M. Hsp70 protein levels and thermotolerance in Drosophila subobscura: A reassessment of the thermal co-adaptation hypothesis. J. Evol. Biol. 2012, 25, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Trelles, F.; Rodríguez, M.A. Rapid micro-evolution and loss of chromosomal diversity in Drosophila in response to climate warming. Evol. Ecol. 1998, 12, 829–838. [Google Scholar] [CrossRef]
- Solé, E.; Balanyà, J.; Sperlich, D.; Serra, L. Long-term changes in the chromosomal inversion polymorphism of Drosophila subobscura. I. Mediterranean populations from Southwestern Europe. Evolution 2002, 56, 830–835. [Google Scholar] [CrossRef] [PubMed]
- Balanyà, J.; Solé, E.; Oller, J.M.; Sperlich, D.; Serra, L. Long-term changes in the chromosomal inversion polymorphism of Drosophila subobscura. II. European populations. J. Zool. Syst. Evol. Res. 2004, 42, 191–201. [Google Scholar] [CrossRef]
- Rodríguez-Trelles, F.; Tarrío, R.; Santos, M. 2013 Genome-wide evolutionary response to a heat wave in Drosophila. Biol. Lett. 2013, 9, 20130228. [Google Scholar] [CrossRef] [Green Version]
- Orengo, D.J.; Puerma, E.; Aguadé, M. Monitoring chromosomal polymorphism in Drosophila subobscura over 40 years. Entomol. Sci. 2016, 19, 215–221. [Google Scholar] [CrossRef]
- Balanyà, J.; Oller, J.M.; Huey, R.B.; Gilchrist, G.W.; Serra, L. Global genetic change tracks global climate warming in Drosophila subobscura. Science 2006, 313, 1773–1775. [Google Scholar] [CrossRef]
- Balanyà, J.; Huey, R.B.; Gilchrist, G.W.; Serra, L. The chromosomal polymorphism of Drosophila subobscura: A microevolutionary weapon to monitor global change. Heredity 2009, 103, 364–367. [Google Scholar] [CrossRef] [Green Version]
- Madrenas, R.; Balanyà, J.; Arenas, C.; Khadem, M.; Mestres, F. Global warming and chromosomal inversion adaptation in isolated islands: Drosophila subobscura populations from Madeira. Entomol. Sci. 2020, 23, 74–85. [Google Scholar] [CrossRef] [Green Version]
- Khadem, M.; Arenas, C.; Balanyà, J.; Mestres, F. Long-term changes in the inversion chromosomal polymorphism: Drosophila subobscura population from Rasht (north of Iran). J. Genet. 2022, 101, 45. [Google Scholar] [CrossRef] [PubMed]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Rubel, F.; Kottek, M. Comments on: ‘‘The thermal zones of the Earth’’ by Wladimir Köppen (1884). Meteorol. Z. 2011, 20, 361–365. [Google Scholar] [CrossRef]
- Zivanovic, G.; Arenas, C.; Mestres, F. Adaptation of Drosophila subobscura chromosomal inversions to climatic variables: The Balkan natural population of Avala. Genetica 2021, 149, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Galludo, M.; Canals, J.; Pineda-Cirera, L.; Esteve, C.; Rosselló, M.; Balanyà, J.; Arenas, C.; Mestres, F. Climatic adaptation of chromosomal inversions in Drosophila subobscura. Genetica 2018, 146, 433–441. [Google Scholar] [CrossRef]
- Zivanovic, G.; Andjelkovic, M.; Marinkovic, D. Chromosomal inversion polymorphism of Drosophila subobscura from southeastern part of Europe. J. Zool. Syst. Evol. Res. 2002, 40, 201–204. [Google Scholar] [CrossRef]
- Zivanovic, G.; Arenas, C.; Mestres, F. Short- and long-term changes in chromosomal inversion polymorphism and global warming: Drosophila subobscura from the Balkans. Isr. J. Ecol. Evol. 2012, 58, 289–311. [Google Scholar] [CrossRef]
- Zivanovic, G.; Milanovic, M.; Andjelkovic, M. Chromosomal inversion polymorphism of Drosophila subobscura populations from Jastrebac Mountain shows temporal and habitat-related changes. J. Zool. Syst. Evol. Res. 1995, 33, 81–83. [Google Scholar] [CrossRef]
- Zivanovic, G. Seasonal changes in chromosomal inversion polymorphism in a Drosophila subobscura natural population from a south-eastern European continental refugium of the last glaciation period. Russ. J. Genet. 2007, 43, 1344–1349. [Google Scholar] [CrossRef]
- Zivanovic, G.; Arenas, C.; Mestres, F. Rate of change for the thermal adapted inversions in Drosophila subobscura. Genetica 2019, 147, 401–409. [Google Scholar] [CrossRef]
- Kunze-Mühl, E.; Müller, E. Weitere Untersuchungen über die chromosomale Struktur und die natürlichen Strukturtypen von Drosophila subobscura. Chromosoma 1958, 9, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [Green Version]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at high resolution for the earth’s land surface areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Carson, H.L. The genetic characteristics of marginal populations of Drosophila. Cold Spring Harb. Symp. Quant. Biol. 1955, 20, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Arenas, C.; Zivanovic, G.; Mestres, F. Chromosomal Thermal Index: A comprehensive way to integrate the thermal adaptation of Drosophila subobscura whole karyotype. Genome 2018, 61, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, A. On a measure of divergence between two multinomial populations. Sankhya 1946, 7, 401–406. [Google Scholar]
- Andjelkovic, M.; Sperlich, D. Inversion polymorphism in a Pannonian population of Drosophila subobscura. Egypt. J. Genet. Cytol. 1973, 2, 144–147. [Google Scholar]
- Araúz, P.A.; Mestres, F.; Pegueroles, C.; Arenas, C.; Tzannidakis, G.; Krimbas, C.B.; Serra, L. Tracking the origin of the American colonization by Drosophila subobscura: Genetic comparison between Eastern and Western Mediterranean populations. J. Zool. Syst. Evol. Res. 2009, 47, 25–34. [Google Scholar] [CrossRef]
- Greuter, M.W. Vergleich der Ausbreitungsaktivitaten von Drosophila subobscura und Drosophila obscura. Rev. Suisse Zool. 1963, 70, 759–856. [Google Scholar] [CrossRef]
- Begon, M. Dispersal, density and microdistribution in Drosophila subobscura Collin. J. Anim. Ecol. 1976, 45, 441–456. [Google Scholar] [CrossRef]
- Loukas, M.; Krimbas, C.B. The genetics of Drosophila subobscura populations. X. A study of dispersal. Genetica 1979, 50, 127–134. [Google Scholar] [CrossRef]
- Mestres, F.; Balanya, J.; Pascual, M.; Arenas, C.; Gilchrist, G.W.; Huey, R.B.; Serra, L. Evolution of Chilean colonizing populations of Drosophila subobscura: Lethal genes and chromosomal arrangements. Genetica 2009, 136, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Ayala, F.J.; Serra, L.; Prevosti, A. A grand experiment in evolution: The Drosophila subobscura colonization of the Americas. Genome 1989, 31, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, A.A.; Sgrò, C.M.; Weeks, A.R. Chromosomal inversion polymorphisms and adaptation. Trends Ecol. Evol. 2004, 19, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Sun, C.; Zhu, J.; Zhang, R.; Li, W. Increased European heat waves in recent decades in response to shrinking Arctic sea ice and Eurasian snow cover. Npj Clim. Atmos. Sci. 2020, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Lhotka, O.; Kyselý, J. The 2021 European heat wave in the context of past major heat waves. Earth Space Sci. 2022, 9, e2022EA002567. [Google Scholar] [CrossRef]
- Fontdevila, A.; Zapata, C.; Alvarez, G.; Sanchez, L.; Méndez, J.; Enriquez, I. Genetic coadaptation in the chromosomal polymorphism of Drosophila subobscura. I. Seasonal changes of gametic disequilibrium in a natural population. Genetics 1983, 105, 935–955. [Google Scholar] [CrossRef]
- Rodríguez-Trelles, F.; Alvarez, G.; Zapata, C. Time-series analysis of seasonal changes of the O inversion polymorphism of Drosophila subobscura. Genetics 1996, 142, 179–187. [Google Scholar] [CrossRef]
- Powell, J.R. Progress and Prospects in Evolutionary Biology. The Drosophila Model; Oxford University Press: New York, NY, USA, 1997. [Google Scholar]
- Kirkpatrick, M.; Barton, N. Chromosome inversions, local adaptation and speciation. Genetics 2006, 173, 419–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, A.A.; Rieseberg, L.H. Revisiting the impact of inversions in evolution: From population genetic markers to drivers of adaptive shifts and speciation? Annu. Rev. Ecol. Evol. Syst. 2008, 39, 21–42. [Google Scholar] [CrossRef] [Green Version]
- Götz, W. Chromosomaler Polymorphismus in einem Muster von Drosophila subobscura aus Marokko, mit Darstellung der Heterozygotieverhfiltnisse als Heterozygotiediagramme. Z. indukt. Abstamm.- u. VererbLehre 1965, 97, 40–45. [Google Scholar]
- Prevosti, A. Chromosomal inversion polymorphism in the southwestern range of Drosophila subobscura distribution area. Genetica 1974, 45, 111–124. [Google Scholar] [CrossRef]
- Sperlich, D. Chromosomale Strukturanalyse und Fertilitätsprüfung einer Marginalpopulation von Drosophila subobscura. Z. VererbLehre 1964, 95, 73–81. [Google Scholar]
- Pinsker, W.; Sperlich, D. Allozyme variation in natural populations of Drosophila subobscura along a north-south gradient. Genetica 1979, 50, 207–219. [Google Scholar] [CrossRef]
- Begon, M. Temporal variations in the reproductive condition of Drosophila obscura Fallén and D. subobscura Collin. Oecologia 1976, 23, 31–47. [Google Scholar] [CrossRef]
- Argemí, M.; Monclús, M.; Mestres, F.; Serra, L. Comparative analysis of a community of Drosophilids (Drosophilidae; Diptera) sampled in two periods widely separated in time. J. Zool. Syst. Evol. Res. 1999, 37, 203–210. [Google Scholar] [CrossRef]
- Lincoln, R.J.; Boxshall, G.A.; Clark, P.F. A Dictionary of Ecology, Evolution and Systematics; Cambridge University Press: Cambridge, UK, 1982. [Google Scholar]
- Laayouni, H.; García-Franco, F.; Chávez-Sandoval, B.E.; Trotta, V.; Beltran, S.; Corominas, M.; Santos, M. Thermal evolution of gene expression profiles in Drosophila subobscura. BMC Evol. Biol. 2007, 19, 42. [Google Scholar] [CrossRef] [Green Version]
- Franks, S.J.; Hoffmann, A.A. Genetics of climate change adaptation. Annu. Rev. Genet. 2012, 46, 185–208. [Google Scholar] [CrossRef] [PubMed]
- Orengo, D.J.; Puerma, E.; Papaceit, M.; Segarra, C.; Aguadé, M. Dense gene physical maps of the non-model species Drosophila subobscura. Chromosome Res. 2017, 25, 145–154. [Google Scholar] [CrossRef]
- Karageorgiou, C.; Tarrío, R.; Rodríguez-Trelles, F. The cyclically seasonal Drosophila subobscura inversion O7 originated from fragile genomic sites and relocated immunity and metabolic genes. Front. Genet. 2020, 11, 565836. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chrom. Inversion | Thermal Adapt. | Year | |||
|---|---|---|---|---|---|
| C/W | 2019 | 2020 | 2021 | 2022 | |
| Ast | C | 54 (44.3%) | 57 (37.7%) | 61 (40.7%) | 63 (41.7%) |
| A1 | C | 44 (36.1%) | 69 (45.7%) | 66 (44.0%) | 73 (48.3%) |
| A2 | W | 24 (19.6%) | 25 (16.5%) | 23 (15.3%) | 15 (9.9%) |
| TOTAL | 122 | 151 | 150 | 151 | |
| Jst | C | 68 (27.8%) | 71 (23.5%) | 65 (21.7%) | 65 (21.5%) |
| J1 | W | 172 (70.5%) | 229 (75.8%) | 232 (77.3%) | 234 (77.5%) |
| J3+4 | N | 4 (1.6%) | 2 (0.7%) | 3 (1.0%) | 3 (1.0%) |
| TOTAL | 244 | 302 | 300 | 302 | |
| Ust | C | 41 (16.8%) | 59 (19.5%) | 53 (17.7%) | 54 (17.9%) |
| U1 | N | 0 | 3 (0.9%) | 2 (0.7%) | 3 (1.0%) |
| U1+2 | W | 132 (54.1%) | 160 (52.9%) | 164 (54.7%) | 138 (45.7%) |
| U1+2+6 | N | 64 (26.2%) | 63 (20.9%) | 70 (23.3%) | 102 (33.8%) |
| U1+8+2 | W | 7 (2.9%) | 17 (5.6%) | 11 (3.7%) | 5 (1.6%) |
| TOTAL | 244 | 302 | 300 | 302 | |
| Est | C | 106 (43.4%) | 139 (46.0%) | 117 (39.0%) | 150 (49.7%) |
| E8 | N | 43 (17.6%) | 67 (22.2%) | 62 (20.7%) | 75 (24.8%) |
| E1+2 | N | 10 (4.1%) | 9 (2.9%) | 16 (5.3%) | 12 (3.9%) |
| E1+2+9 | W | 70 (28.7%) | 77 (25.5%) | 98 (32.7%) | 50 (16.5%) |
| E1+2+9+12 | W | 15 (6.1%) | 10 (3.3%) | 7 (2.3%) | 15 (4.9%) |
| TOTAL | 244 | 302 | 300 | 302 | |
| Ost | C | 83 (34.0%) | 98 (32.4%) | 98 (32.7%) | 106 (35.1%) |
| O6 | N | 6 (2.4%) | 4 (1.3%) | 1 (0.3%) | 4 (1.3%) |
| O7 | N | 4 (1.6%) | 0 | 0 | 0 |
| O3+4 | W | 88 (36.1%) | 131 (43.4%) | 126 (42.0%) | 115 (38.1%) |
| O3+4+1 | W | 35 (14.3%) | 35 (11.6%) | 36 (12.0%) | 32 (10.6%) |
| O3+4+6 | N | 2 (0.8%) | 7 (2.3%) | 6 (2.0%) | 8 (2.6%) |
| O3+4+7 | N | 1 (0.4%) | 0 | 3 (1.0%) | 1 (0.3%) |
| O3+4+8 | W | 10 (4.1%) | 11 (3.6%) | 9 (3.0%) | 17 (5.6%) |
| O3+4+17 | N | 0 | 1 (0.3%) | 0 | 0 |
| O3+4+22 | N | 15 (6.1%) | 15 (4.9%) | 21 (7.0%) | 19 (6.3%) |
| TOTAL | 244 | 302 | 300 | 302 | |
| CTI | 0.165 | 0.170 | 0.211 | 0.097 | |
| Year | ||||
|---|---|---|---|---|
| Chromosomal Karyotypes | 2019 | 2020 | 2021 | 2022 |
| Jst/Jst | 12 (9.8%) | 12 (7.9%) | 8 (5.3%) | 12 (7.9%) |
| Jst/J1 | 44 (36.1%) | 47 (31.1%) | 48 (32.0%) | 41 (27.1%) |
| Jst/J3+4 | 0 | 0 | 1 (0.7%) | 0 (%) |
| J1/J1 | 62 (50.8%) | 90 (59.6%) | 91 (60.7%) | 95 (62.9%) |
| J1/J3+4 | 4 (3.3%) | 2 (1.3%) | 2 (1.3%) | 3 (1.9%) |
| TOTAL | 122 | 151 | 150 | 151 |
| Ust/Ust | 3 (2.4%) | 11 (7.3%) | 10 (6.7%) | 6 (3.9%) |
| Ust/U1 | 0 | 0 | 1 (0.7%) | 0 |
| Ust/U1+2 | 21 (17.2%) | 19 (12.6%) | 26 (17.3%) | 22 (14.6%) |
| Ust/U1+2+6 | 13 (10.6%) | 10 (6.6%) | 6 (4.0%) | 20 (13.2%) |
| Ust/U1+8+2 | 1 (0.8%) | 8 (5.3%) | 0 | 0 |
| U1/U1 | 0 | 1 (0.7%) | 0 | 1 (0.7%) |
| U1/U1+2 | 0 | 0 | 1 (0.7%) | 0 |
| U1/U1+2+6 | 0 | 1 (0.7%) | 0 | 1 (0.7%) |
| U1+2/U1+2 | 36 (29.5%) | 50 (33.1%) | 46 (30.7%) | 43 (28.5) |
| U1+2/U1+2+6 | 34 (27.8%) | 36 (23.8%) | 36 (24.0%) | 26 (17.2%) |
| U1+2/U1+8+2 | 5 (4.1%) | 5 (3.3%) | 9 (6.5%) | 4 (2.6%) |
| U1+2+6/U1+2+6 | 8 (6.5%) | 6 (3.9%) | 14 (9.3%) | 27 (17.9%) |
| U1+2+6/U1+8+2 | 1 (0.8%) | 4 (2.6%) | 0 | 1 (0.7%) |
| U1+8+2/U1+8+2 | 0 | 0 | 1 (0.7%) | 0 |
| TOTAL | 122 | 151 | 150 | 151 |
| Est/Est | 21 (17.2%) | 42 (27.8%) | 32 (21.3%) | 39 (25.8%) |
| Est/E1+2 | 7 (5.7%) | 5 (3.3%) | 4 (2.7%) | 3 (1.9%) |
| Est/E1+2+9 | 27 (22.1%) | 23 (15.2%) | 33 (22.0%) | 24 (15.9%) |
| Est/E1+2+9+12 | 9 (7.4%) | 4 (2.6%) | 0 | 9 (5.9%) |
| Est/E8 | 21 (17.2%) | 23 (15.2%) | 16 (10.7%) | 36 (23.8%) |
| E1+2/E1+2 | 0 | 0 | 2 (1.3%) | 0 |
| E1+2/E1+2+9 | 1 (0.8%) | 1 (0.7%) | 4 (2.7%) | 2 (1.3%) |
| E1+2/E1+2+9+12 | 0 | 0 | 1 (0.7%) | 1 (0.7%) |
| E1+2/E8 | 2 (1.6%) | 3 (2.0%) | 3 (2.0%) | 6 (3.9%) |
| E1+2+9/E1+2+9 | 15 (12.3%) | 14 (9.3%) | 19 (12.7%) | 7 (4.6%) |
| E1+2+9/E1+2+9+12 | 1 (0.8%) | 0 | 2 (1.3%) | 1 (0.7%) |
| E1+2+9/E8 | 11 (9.0%) | 25 (16.5%) | 21 (14.0%) | 9 (5.9%) |
| E1+2+9+12/E1+2+9+12 | 2 (1.6%) | 1 (0.7%) | 0 | 2 (1.3%) |
| E1+2+9+12/E8 | 1 (0.8%) | 4 (2.6%) | 4 (2.7%) | 0 |
| E8/E8 | 4 (3.3%) | 6 (3.9%) | 9 (6%) | 12 (7.9%) |
| TOTAL | 122 | 151 | 150 | 151 |
| Ost/Ost | 12 (9.8%) | 17 (11.2%) | 16 (10.7%) | 36 (23.8%) |
| Ost/O6 | 1 (0.8%) | 3 (2%) | 1 (0.7%) | 2 (1.3%) |
| Ost/O7 | 4 (3.3%) | 0 | 0 | 0 |
| Ost/O3+4 | 30 (24.6%) | 43 (28.5%) | 41 (27.3%) | 20 (13.2%) |
| Ost/O3+4+1 | 14 (11.5%) | 11 (7.3%) | 13 (8.7%) | 6 (3.9%) |
| Ost/O3+4+6 | 0 | 1 (0.7%) | 3 (2.0%) | 0 |
| Ost/O3+4+7 | 1 (0.8%) | 0 | 0 | 1 (0.7%) |
| Ost/O3+4+8 | 4 (3.3%) | 1 (0.7%) | 2 (1.3%) | 3 (2.0%) |
| Ost/O3+4+22 | 5 (4.1%) | 5 (3.3%) | 6 (4.0%) | 2 (1.3%) |
| O6/O3+4 | 3 (2.4%) | 1 (0.7%) | 0 | 0 |
| O6/O3+4+8 | 0 | 0 | 0 | 2 (1.3%) |
| O6/O3+4+22 | 2 (1.6%) | 0 | 0 | 0 |
| O3+4/O3+4 | 18 (14.7%) | 28 (18.5%) | 31 (20.7%) | 31 (20.5%) |
| O3+4/O3+4+1 | 9 (7.4%) | 15 (9.9%) | 12 (8.0%) | 13 (8.6%) |
| O3+4/O3+4+6 | 1 (0.8%) | 3 (2.0%) | 3 (2.0%) | 3 (2.0%) |
| O3+4/O3+4+8 | 4 (3.3%) | 9 (5.9%) | 0 | 3 (2.0%) |
| O3+4/O3+4+17 | 0 | 1 (0.7%) | 0 | 0 |
| O3+4/O3+4+22 | 5 (4.1%) | 3 (2.0%) | 8 (5.3%) | 14 (9.3%) |
| O3+4+1/O3+4+1 | 4 (3.3%) | 1 (0.7%) | 5 (3.3%) | 5 (3.3%) |
| O3+4+1/O3+4+6 | 0 | 3 (2.0%) | 0 | 0 |
| O3+4+1/O3+4+8 | 1 (0.8%) | 0 | 1 (0.7%) | 1 (0.7%) |
| O3+4+1/O3+4+22 | 3 (2.4%) | 4 (2.6%) | 0 | 2 (1.3%) |
| O3+4+6/O3+4+6 | 0 | 0 | 0 | 2 (1.3%) |
| O3+4+6/O3+4+8 | 1 (0.8%) | 0 | 0 | 0 |
| O3+4+6/O3+4+22 | 0 | 0 | 0 | 1 (0.7%) |
| O3+4+7/O3+4+7 | 0 | 0 | 1 (0.7%) | 0 |
| O3+4+7/O3+4+8 | 0 | 0 | 1 (0.7%) | 0 |
| O3+4+8/O3+4+8 | 0 | 0 | 2 (1.3%) | 4 (2.6%) |
| O3+4+8/O3+4+22 | 0 | 1 (0.7%) | 1 (0.7%) | 0 |
| O3+4+22/O3+4+22 | 0 | 1 (0.7%) | 3 (2.0%) | 0 |
| TOTAL | 122 | 151 | 150 | 151 |
| IFR | 79.84 ± 0.90 | 82.28 ± 0.77 | 82.93 ± 0.75 | 84.45 ± 0.75 |
| Climatic Variables | Component Coefficients | ||
|---|---|---|---|
| PC1 | PC2 | PC3 | |
| Tmean | 0.856 | −0.205 | −0.022 |
| Tmin | 0.829 | 0.274 | 0.108 |
| Tmax | 0.781 | −0.405 | −0.034 |
| Rainfall | 0.520 | 0.705 | −0.083 |
| Humidity | −0.117 | 0.872 | 0.012 |
| Chrom. | Nominal p | Adjusted p | ||||
|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | |
| Ast | 0.4770 | 0.6510 | 0.7280 | 0.6559 | 0.7723 | 0.8712 |
| A1 | 0.0369 | 0.0336 | 0.5584 | 0.2706 | 0.1379 | 0.8388 |
| A2 | 0.7240 | 0.2220 | 0.1850 | 0.8424 | 0.4440 | 0.6783 |
| Jst | 0.7460 | 0.9680 | 0.6100 | 0.8424 | 0.9690 | 0.8388 |
| J1 | 0.0031 | 0.0376 | 0.4429 | 0.0678 | 0.1379 | 0.8388 |
| J3+4 | 0.7658 | 0.6590 | 0.5542 | 0.8424 | 0.7723 | 0.8388 |
| Ust | 0.2470 | 0.2440 | 0.3170 | 0.4528 | 0.4473 | 0.8388 |
| U1+2 | 0.0924 | 0.7948 | 0.1344 | 0.3104 | 0.8743 | 0.5914 |
| U1+2+6 | 0.3961 | 0.0197 | 0.0229 | 0.6224 | 0.1083 | 0.2519 |
| U1+8+2 | 0.5431 | 0.6232 | 0.0065 | 0.7028 | 0.7723 | 0.1430 |
| Est | 0.4403 | 0.0029 | 0.7555 | 0.6458 | 0.0399 | 0.8712 |
| E8 | 0.0599 | 0.0120 | 0.7410 | 0.3065 | 0.0880 | 0.8712 |
| E1+2 | 0.2100 | 0.6230 | 0.5210 | 0.4528 | 0.7723 | 0.8388 |
| E1+2+9 | 0.0697 | 0.0036 | 0.0466 | 0.3065 | 0.0399 | 0.3420 |
| E1+2+9+12 | 0.1230 | 0.3820 | 0.3240 | 0.3382 | 0.6465 | 0.8388 |
| Ost | 0.2460 | 0.1990 | 0.9530 | 0.4528 | 0.4440 | 0.9566 |
| O6 | 0.0988 | 0.4382 | 0.9566 | 0.3104 | 0.6886 | 0.9566 |
| O3+4 | 0.0150 | 0.2150 | 0.0752 | 0.1650 | 0.4440 | 0.4136 |
| O3+4+1 | 0.9410 | 0.6670 | 0.7920 | 0.9410 | 0.7723 | 0.8712 |
| O3+4+6 | 0.1760 | 0.1080 | 0.5910 | 0.4302 | 0.3394 | 0.8388 |
| O3+4+8 | 0.9030 | 0.1320 | 0.4710 | 0.9410 | 0.3630 | 0.8388 |
| O3+4+22 | 0.2810 | 0.9690 | 0.5660 | 0.4755 | 0.9690 | 0.8388 |
| Chrom. | Ratio Increasing 1 Unit | Ratio Decreasing 1 Unit | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Tmean | Tmin | Tmax | Hm | Rfl | Tmean | Tmin | Tmax | Hm | Rf | |
| Ast | 0.97 | 0.95 | 0.98 | 0.98 | 0.96 | 1.03 | 1.05 | 1.02 | 1.02 | 1.04 |
| A1 | 0.91 | 0.86 | 0.94 | 0.90 | 0.84 | 1.10 | 1.17 | 1.06 | 1.11 | 1.19 |
| A2 | 1.00 | 1.09 | 0.97 | 1.13 | 1.11 | 1.00 | 0.92 | 1.03 | 0.88 | 0.90 |
| Jst | 1.02 | 1.02 | 1.02 | 1.00 | 1.01 | 0.98 | 0.98 | 0.98 | 1.00 | 0.99 |
| J1 | 0.93 | 0.90 | 0.95 | 0.95 | 0.90 | 1.08 | 1.11 | 1.06 | 1.05 | 1.11 |
| J3+4 | 1.05 | 1.09 | 1.02 | 1.11 | 1.16 | 0.95 | 0.92 | 0.98 | 0.90 | 0.86 |
| Ust | 0.94 | 0.92 | 0.96 | 0.94 | 0.90 | 1.06 | 1.09 | 1.04 | 1.06 | 1.12 |
| U1+2 | 0.94 | 0.95 | 0.94 | 1.02 | 0.97 | 1.07 | 1.05 | 1.06 | 0.98 | 1.03 |
| U1+2+6 | 0.99 | 0.91 | 1.02 | 0.89 | 0.90 | 1.01 | 1.10 | 0.98 | 1.12 | 1.12 |
| U1+8+2 | 0.89 | 0.99 | 0.87 | 1.11 | 0.98 | 1.13 | 1.01 | 1.15 | 0.90 | 1.02 |
| Est | 1.00 | 0.94 | 1.03 | 0.89 | 0.89 | 1.00 | 1.07 | 0.98 | 1.12 | 1.12 |
| E8 | 0.92 | 0.86 | 0.96 | 0.88 | 0.83 | 1.08 | 1.17 | 1.04 | 1.13 | 1.20 |
| E1+2 | 0.85 | 0.87 | 0.85 | 1.09 | 0.97 | 1.18 | 1.15 | 1.18 | 0.92 | 1.04 |
| E1+2+9 | 0.88 | 0.98 | 0.85 | 1.19 | 1.07 | 1.14 | 1.02 | 1.17 | 0.84 | 0.93 |
| E1+2+9+12 | 1.28 | 1.17 | 1.29 | 0.86 | 1.05 | 0.78 | 0.85 | 0.77 | 1.16 | 0.95 |
| Ost | 0.96 | 0.93 | 0.98 | 0.95 | 0.92 | 1.04 | 1.07 | 1.02 | 1.05 | 1.08 |
| O6 | 1.82 | 1.59 | 1.83 | 0.74 | 1.17 | 0.55 | 0.63 | 0.55 | 1.35 | 0.86 |
| O3+4 | 0.91 | 0.9 | 0.93 | 0.96 | 0.89 | 1.10 | 1.11 | 1.08 | 1.04 | 1.12 |
| O3+4+1 | 0.99 | 1.01 | 0.98 | 1.03 | 1.02 | 1.01 | 0.99 | 1.02 | 0.97 | 0.98 |
| O3+4+6 | 0.77 | 0.66 | 0.86 | 0.75 | 0.62 | 1.30 | 1.52 | 1.17 | 1.33 | 1.62 |
| O3+4+8 | 1.07 | 0.95 | 1.12 | 0.82 | 0.87 | 0.94 | 1.06 | 0.90 | 1.22 | 1.15 |
| O3+4+22 | 0.89 | 0.89 | 0.9 | 1.02 | 0.94 | 1.12 | 1.12 | 1.11 | 0.98 | 1.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zivanovic, G.; Arenas, C.; Mestres, F. The Adaptive Value of Chromosomal Inversions and Climatic Change—Studies on the Natural Populations of Drosophila subobscura from the Balkans. Insects 2023, 14, 596. https://doi.org/10.3390/insects14070596
Zivanovic G, Arenas C, Mestres F. The Adaptive Value of Chromosomal Inversions and Climatic Change—Studies on the Natural Populations of Drosophila subobscura from the Balkans. Insects. 2023; 14(7):596. https://doi.org/10.3390/insects14070596
Chicago/Turabian StyleZivanovic, Goran, Concepció Arenas, and Francesc Mestres. 2023. "The Adaptive Value of Chromosomal Inversions and Climatic Change—Studies on the Natural Populations of Drosophila subobscura from the Balkans" Insects 14, no. 7: 596. https://doi.org/10.3390/insects14070596
APA StyleZivanovic, G., Arenas, C., & Mestres, F. (2023). The Adaptive Value of Chromosomal Inversions and Climatic Change—Studies on the Natural Populations of Drosophila subobscura from the Balkans. Insects, 14(7), 596. https://doi.org/10.3390/insects14070596