
In vitro–in vivo Validation of Stimulatory Effect of Oat Ingredients on Lactobacilli



 ,
,
Abstract
:1. Introduction
2. Results
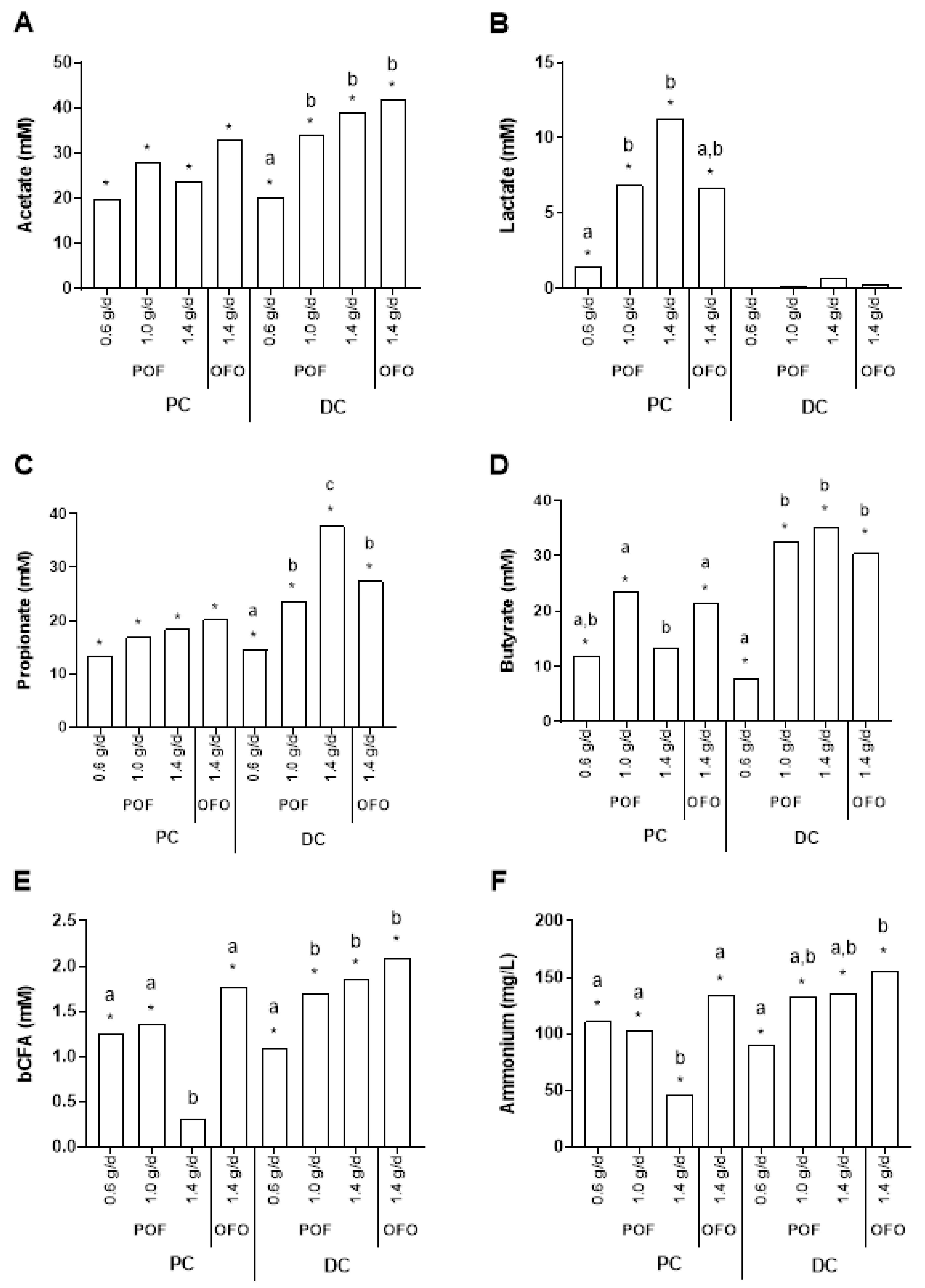
2.1. Altered Microbial Metabolic Activity in Response to Oat Treatment in vitro
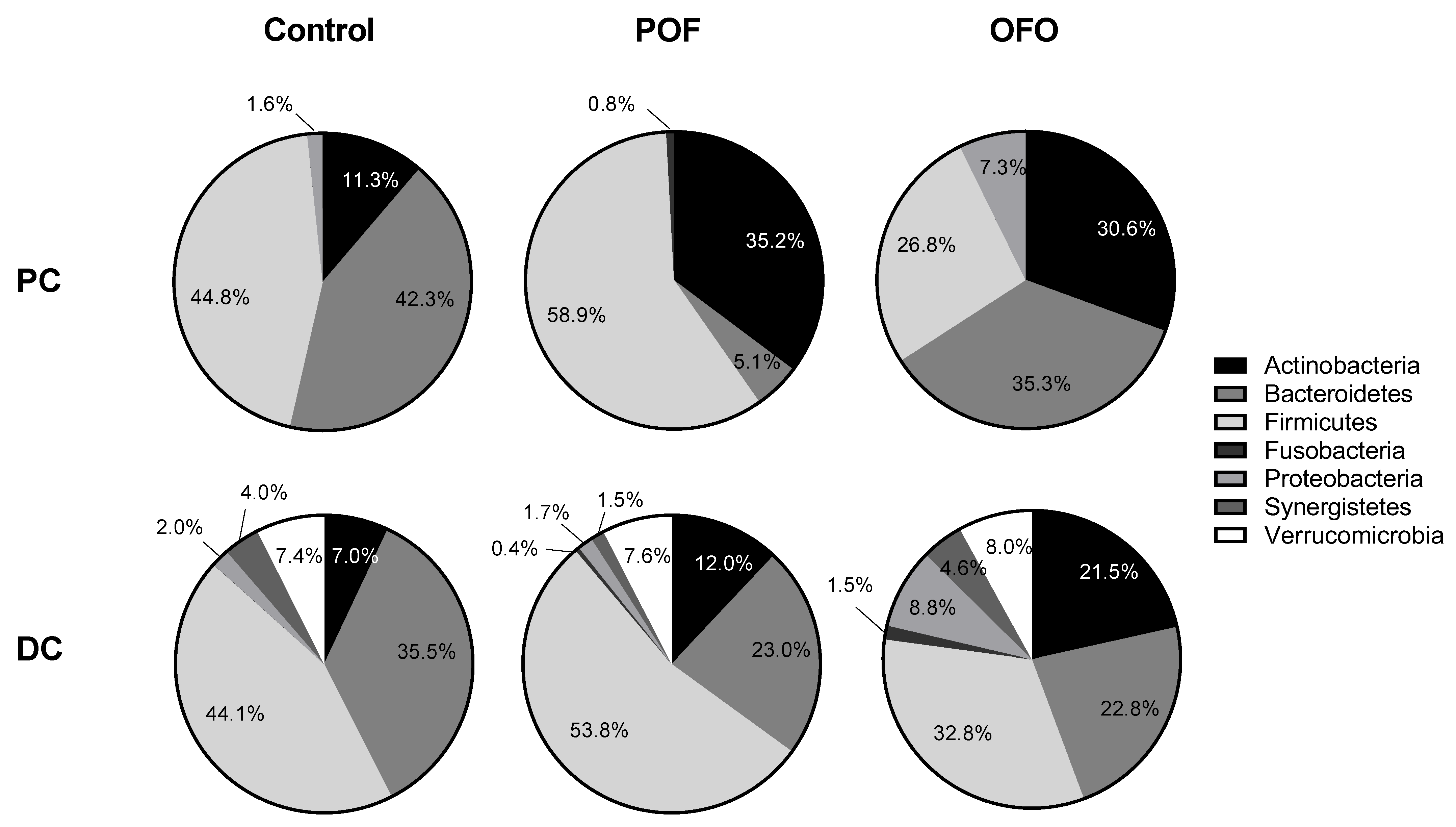
2.2. Altered Microbial Composition in Response to Oat Treatment in vitro
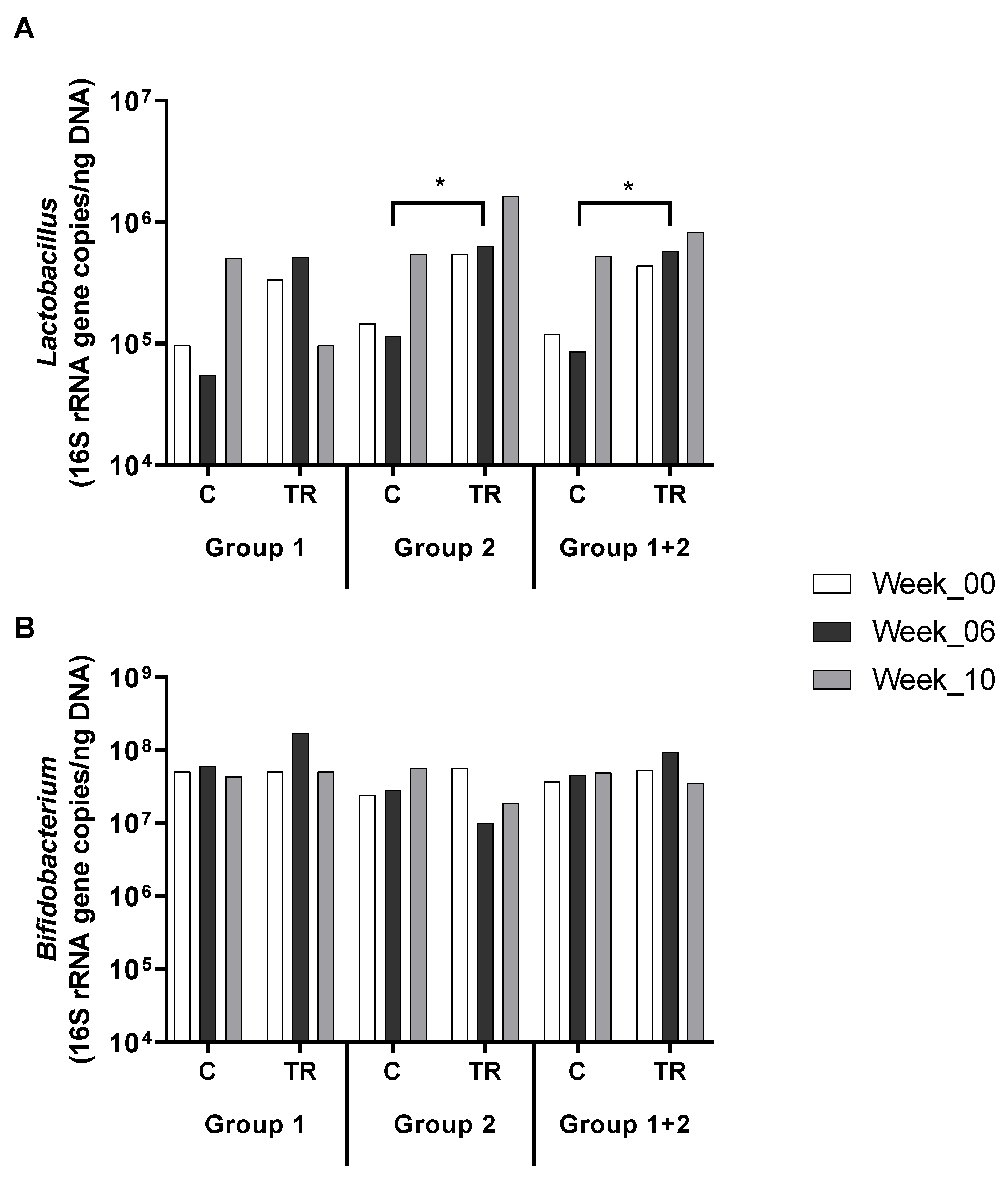
2.3. In vitro–in vivo Comparison of Microbial Response to OFO Treatment
3. Discussion
4. Materials and Methods
4.1. Chemicals and Test Product
4.2. Simulator of the Human Intestinal Microbial Ecosystem (SHIME®)
4.3. In vivo Study
4.4. Microbial Metabolic Activity
4.5. DNA Extraction
4.6. Microbial Community Analysis through qPCR
4.7. Microbial Community Analysis through 16S-Targeted Illumina Sequencing
4.8. Flow Cytometric Determination
4.9. Data and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daou, C.; Zhang, H. Oat Beta-Glucan: Its Role in Health Promotion and Prevention of Diseases. Compr. Rev. Food Sci. Food Saf. 2012, 11, 355–365. [Google Scholar] [CrossRef]
- Braaten, J.T.; Wood, P.J.; Scott, F.W.; Wolynetz, M.S.; Lowe, M.K.; Bradley-White, P.; Collins, M.W. Oat beta-glucan reduces blood cholesterol concentration in hypercholesterolemic subjects. Eur. J. Clin. Nutr. 1994, 48, 465–474. [Google Scholar] [PubMed]
- El Khoury, D.; Cuda, C.; Luhovyy, B.L.; Anderson, G.H. Beta Glucan: Health Benefits in Obesity and Metabolic Syndrome. J. Nutr. Metab. 2012, 2012, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Doehlert, D.C.; Moore, W.R. Rheological Properties of (1→3),(1→4)-β-d-Glucans from Raw, Roasted, and Steamed Oat Groats. Cereal Chem. 1998, 75, 433–438. [Google Scholar] [CrossRef]
- Grundy, M.M.L.; Quint, J.; Rieder, A.; Ballance, S.; Dreiss, C.A.; Butterworth, P.J.; Ellis, P.R. Impact of hydrothermal and mechanical processing on dissolution kinetics and rheology of oat β-glucan. Carbohydr. Polym. 2017, 166, 387–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Abbeele, P.; Kamil, A.; Fleige, L.; Chung, Y.; De Chavez, P.; Marzorati, M. Different Oat Ingredients Stimulate Specific Microbial Metabolites in the Gut Microbiome of Three Human Individuals in Vitro. ACS Omega 2018, 3, 12446–12456. [Google Scholar] [CrossRef]
- Johansson, M.L.; Nobaek, S.; Berggren, A.; Nyman, M.; Bjorck, I.; Ahrne, S.; Jeppsson, B.; Molin, G. Survival of Lactobacillus plantarum DSM 9843 (299v), and effect on the short-chain fatty acid content of faeces after ingestion of a rose-hip drink with fermented oats. Int. J. Food Microbiol. 1998, 42, 29–38. [Google Scholar] [CrossRef]
- Mårtensson, O.; Biörklund, M.; Lambo, A.M.; Dueñas-Chasco, M.; Irastorza, A.; Holst, O.; Norin, E.; Welling, G.; Öste, R.; Önning, G. Fermented, ropy, oat-based products reduce cholesterol levels and stimulate the bifidobacteria flora in humans. Nutr. Res. 2005, 25, 429–442. [Google Scholar] [CrossRef]
- Coman, M.M.; Verdenelli, M.C.; Cecchini, C.; Silvi, S.; Vasile, A.; Bahrim, G.E.; Orpianesi, C.; Cresci, A. Effect of buckwheat flour and oat bran on growth and cell viability of the probiotic strains Lactobacillus rhamnosus IMC 501(R), Lactobacillus paracasei IMC 502(R) and their combination SYNBIO(R), in synbiotic fermented milk. Int. J. Food Microbiol. 2013, 167, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Connolly, M.L.; Tzounis, X.; Tuohy, K.M.; Lovegrove, J.A. Hypocholesterolemic and Prebiotic Effects of a Whole-Grain Oat-Based Granola Breakfast Cereal in a Cardio-Metabolic “At Risk” Population. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, U.; Cummins, E. Meta-analysis of the effect of beta-glucan intake on blood cholesterol and glucose levels. Nutrition 2011, 27, 1008–1016. [Google Scholar] [CrossRef]
- Whitehead, A.; Beck, E.J.; Tosh, S.; Wolever, T.M. Cholesterol-lowering effects of oat beta-glucan: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2014, 100, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Ho, H.V.; Sievenpiper, J.L.; Zurbau, A.; Blanco Mejia, S.; Jovanovski, E.; Au-Yeung, F.; Jenkins, A.L.; Vuksan, V. The effect of oat beta-glucan on LDL-cholesterol, non-HDL-cholesterol and apoB for CVD risk reduction: A systematic review and meta-analysis of randomised-controlled trials. Br. J. Nutr. 2016, 116, 1369–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolever, T.M.; Tosh, S.M.; Gibbs, A.L.; Brand-Miller, J.; Duncan, A.M.; Hart, V.; Lamarche, B.; Thomson, B.A.; Duss, R.; Wood, P.J. Physicochemical properties of oat beta-glucan influence its ability to reduce serum LDL cholesterol in humans: A randomized clinical trial. Am. J. Clin. Nutr. 2010, 92, 723–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonsson, A.L.; Backhed, F. Role of gut microbiota in atherosclerosis. Nat. Rev. Cardiol. 2017, 14, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.L.; Tomaro-Duchesneau, C.; Martoni, C.J.; Prakash, S. Cholesterol lowering with bile salt hydrolase-active probiotic bacteria, mechanism of action, clinical evidence, and future direction for heart health applications. Expert Opin. Biol. Ther. 2013, 13, 631–642. [Google Scholar] [CrossRef]
- Joyce, S.A.; Kamil, A.; Fleige, L.; Gahan, C.G.M. The Cholesterol-Lowering Effect of Oats and Oat Beta Glucan: Modes of Action and Potential Role of Bile Acids and the Microbiome. Front. Nutr. 2019, 6. [Google Scholar] [CrossRef]
- Berggren, A.M.; Nyman, E.M.; Lundquist, I.; Bjorck, I.M. Influence of orally and rectally administered propionate on cholesterol and glucose metabolism in obese rats. Br. J. Nutr. 1996, 76, 287–294. [Google Scholar] [CrossRef]
- Lin, Y.; Vonk, R.J.; Slooff, M.J.; Kuipers, F.; Smit, M.J. Differences in propionate-induced inhibition of cholesterol and triacylglycerol synthesis between human and rat hepatocytes in primary culture. Br. J. Nutr. 1995, 74, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Connolly, M.L.; Lovegrove, J.A.; Tuohy, K.M. In Vitro evaluation of the microbiota modulation abilities of different sized whole oat grain flakes. Anaerobe 2010, 16, 483–488. [Google Scholar] [CrossRef]
- Ryan, P.M.; London, L.E.E.; Bjorndahl, T.C.; Mandal, R.; Murphy, K.; Fitzgerald, G.F.; Shanahan, F.; Ross, R.P.; Wishart, D.S.; Caplice, N.M.; et al. Microbiome and metabolome modifying effects of several cardiovascular disease interventions in apo-E−/− mice. Microbiome 2017, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Molly, K.; Vande Woestyne, M.; Verstraete, W. Development of a 5-step multi-chamber reactor as a simulation of the human intestinal microbial ecosystem. Appl. Microbiol. Biotechnol. 1993, 39, 254–258. [Google Scholar] [CrossRef]
- Minekus, M.; Smeets-Peeters, M.; Bernalier, A.; Marol-Bonnin, S.; Havenaar, R.; Marteau, P.; Alric, M.; Fonty, G.; Huis in′t Veld, J.H. A computer-controlled system to simulate conditions of the large intestine with peristaltic mixing, water absorption and absorption of fermentation products. Appl. Microbiol. Biotechnol. 1999, 53, 108–114. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Roos, S.; Eeckhaut, V.; MacKenzie, D.A.; Derde, M.; Verstraete, W.; Marzorati, M.; Possemiers, S.; Vanhoecke, B.; Van Immerseel, F.; et al. Incorporating a mucosal environment in a dynamic gut model results in a more representative colonization by lactobacilli. Microb. Biotechnol. 2012, 5, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Firrman, J.; Tanes, C.; Bittinger, K.; Thomas-Gahring, A.; Wu, G.D.; Van den Abbeele, P.; Tomasula, P.M. Establishing a mucosal gut microbial community in vitro using an artificial simulator. PLoS ONE 2018, 13, e0197692. [Google Scholar] [CrossRef] [PubMed]
- Pridmore, R.D.; Berger, B.; Desiere, F.; Vilanova, D.; Barretto, C.; Pittet, A.C.; Zwahlen, M.C.; Rouvet, M.; Altermann, E.; Barrangou, R.; et al. The genome sequence of the probiotic intestinal bacterium Lactobacillus johnsonii NCC 533. Proc. Natl. Acad. Sci. USA 2004, 101, 2512–2517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokusaeva, K.; Fitzgerald, G.F.; van Sinderen, D. Carbohydrate metabolism in Bifidobacteria. Genes Nutr. 2011, 6, 285–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raybaudi-Massilia, R.M.; Mosqueda-Melgar, J.; Soliva-Fortuny, R.; Martín-Belloso, O. Control of Pathogenic and Spoilage Microorganisms in Fresh-cut Fruits and Fruit Juices by Traditional and Alternative Natural Antimicrobials. Compr. Rev. Food Sci. Food Saf. 2009, 8, 157–180. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Louis, P.; Flint, H.J. Lactate-Utilizing Bacteria, Isolated from Human Feces, That Produce Butyrate as a Major Fermentation Product. Appl. Environ. Microbiol. 2004, 70, 5810. [Google Scholar] [CrossRef] [Green Version]
- Macfarlane, G.T.; Gibson, G.R. Carbohydrate Fermentation, Energy Transduction and Gas Metabolism in the Human Large Intestine. In Gastrointestinal Microbiology: Volume 1 Gastrointestinal Ecosystems and Fermentations; Mackie, R.I., White, B.A., Eds.; Springer US: Boston, MA, USA, 1997; pp. 269–318. [Google Scholar] [CrossRef]
- Lampe, J.W.; Navarro, S.L.; Hullar, M.A.J.; Shojaie, A. Inter-individual differences in response to dietary intervention: Integrating omics platforms towards personalised dietary recommendations. Proc. Nutr. Soc. 2013, 72, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Nagpal, R.; Kumar, R.; Hemalatha, R.; Verma, V.; Kumar, A.; Chakraborty, C.; Singh, B.; Marotta, F.; Jain, S.; et al. Cholesterol-lowering probiotics as potential biotherapeutics for metabolic diseases. Exp. Diabetes Res. 2012, 2012, 902917. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Kang, J.; Choi, S.; Park, H.; Hwang, E.; Kang, Y.; Kim, A.; Holzapfel, W.; Ji, Y. Cholesterol-lowering effect of Lactobacillus rhamnosus BFE5264 and its influence on the gut microbiome and propionate level in a murine model. PLoS ONE 2018, 13, e0203150. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, M.C.; Lajo, T.; Carrión, J.M.; Cuñé, J. Cholesterol-lowering efficacy of Lactobacillus plantarum CECT 7527, 7528 and 7529 in hypercholesterolaemic adults. Br. J. Nutr. 2013, 109, 1866–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.L.; Martoni, C.J.; Parent, M.; Prakash, S. Cholesterol-lowering efficacy of a microencapsulated bile salt hydrolase-active Lactobacillus reuteri NCIMB 30242 yoghurt formulation in hypercholesterolaemic adults. Br. J. Nutr. 2011, 107, 1505–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristek, A.; Wiese, M.; Heuer, P.; Kosik, O.; Schar, M.Y.; Soycan, G.; Alsharif, S.; Kuhnle, G.G.C.; Walton, G.; Spencer, J.P.E. Oat bran, but not its isolated bioactive beta-glucans or polyphenols, have a bifidogenic effect in an in vitro fermentation model of the gut microbiota. Br. J. Nutr. 2019, 121, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Cheung, P.C. Fermentation of beta-glucans derived from different sources by bifidobacteria: Evaluation of their bifidogenic effect. J. Agric. Food Chem. 2011, 59, 5986–5992. [Google Scholar] [CrossRef] [PubMed]
- Takata, K.; Kinoshita, M.; Okuno, T.; Moriya, M.; Kohda, T.; Honorat, J.A.; Sugimoto, T.; Kumanogoh, A.; Kayama, H.; Takeda, K.; et al. The Lactic Acid Bacterium Pediococcus acidilactici Suppresses Autoimmune Encephalomyelitis by Inducing IL-10-Producing Regulatory T Cells. PLoS ONE 2011, 6, e27644. [Google Scholar] [CrossRef]
- Schneeberger, M.; Everard, A.; Gómez-Valadés, A.G.; Matamoros, S.; Ramírez, S.; Delzenne, N.M.; Gomis, R.; Claret, M.; Cani, P.D. Akkermansia muciniphila inversely correlates with the onset of inflammation, altered adipose tissue metabolism and metabolic disorders during obesity in mice. Sci. Rep. 2015, 5, 16643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staka, A.; Bodnieks, E.; Puķītis, A. Impact of Oat-Based Products on Human Gastrointestinal Tract. Proc. Latv. Acad. Sci. 2015, 69, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, K.E.B.; Jensen, B.B.; Hansen, I. Oat Bran but Not a β-Glucan-Enriched Oat Fraction Enhances Butyrate Production in the Large Intestine of Pigs. J. Nutr. 1993, 123, 1235–1247. [Google Scholar] [CrossRef] [PubMed]
- Kedia, G.; Vazquez, J.A.; Charalampopoulos, D.; Pandiella, S.S. In Vitro fermentation of oat bran obtained by debranning with a mixed culture of human fecal bacteria. Curr. Microbiol. 2009, 58, 338–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, S.A.; Shewry, P.R.; Gibson, G.R.; McCleary, B.V.; Rastall, R.A. In Vitro fermentation of oat and barley derived beta-glucans by human faecal microbiota. FEMS Microbiol. Ecol. 2008, 64, 482–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Martin, R.J.; Tulley, R.T.; Raggio, A.M.; McCutcheon, K.L.; Shen, L.; Danna, S.C.; Tripathy, S.; Hegsted, M.; Keenan, M.J. Dietary resistant starch upregulates total GLP-1 and PYY in a sustained day-long manner through fermentation in rodents. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1160–E1166. [Google Scholar] [CrossRef] [Green Version]
- Al-Lahham, S.H.; Roelofsen, H.; Priebe, M.; Weening, D.; Dijkstra, M.; Hoek, A.; Rezaee, F.; Venema, K.; Vonk, R.J. Regulation of adipokine production in human adipose tissue by propionic acid. Eur. J. Clin. Investig. 2010, 40, 401–407. [Google Scholar] [CrossRef]
- Al-Lahham, S.; Roelofsen, H.; Rezaee, F.; Weening, D.; Hoek, A.; Vonk, R.; Venema, K. Propionic acid affects immune status and metabolism in adipose tissue from overweight subjects. Eur. J. Clin. Investig. 2012, 42, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Jan, G.; Belzacq, A.S.; Haouzi, D.; Rouault, A.; Metivier, D.; Kroemer, G.; Brenner, C. Propionibacteria induce apoptosis of colorectal carcinoma cells via short-chain fatty acids acting on mitochondria. Cell Death Differ. 2002, 9, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Shibata, K.; Sakamoto, M.; Tomita, S.; Benno, Y. Prevotella copri sp. nov. and Prevotella stercorea sp. nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2007, 57, 941–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, E.; Grootaert, C.; Verstraete, W.; Van de Wiele, T. Propionate as a health-promoting microbial metabolite in the human gut. Nutr. Rev. 2011, 69, 245–258. [Google Scholar] [CrossRef]
- Kang, E.; Crouse, A.; Chevallier, L.; Pontier, S.; Alzahrani, A.; Silue, N.; Campbell-Valois, F.-X.; Montagutelli, X.; Gruenheid, S.; Malo, D. Enterobacteria and host resistance to infection. Mamm. Genome 2018, 29. [Google Scholar] [CrossRef] [PubMed]
- Possemiers, S.; Verthe, K.; Uyttendaele, S.; Verstraete, W. PCR-DGGE-based quantification of stability of the microbial community in a simulator of the human intestinal microbial ecosystem. FEMS Microbiol. Ecol. 2004, 49, 495–507. [Google Scholar] [CrossRef]
- Alliance, I.U.N. National Adult Nutrition Survey: Summary Report on Food and Nutrient Intakes, Physical Measurements, Physical Activity Patterns and Food Choice Motives; Trinity College Dublin, Irish Universities Nutrition Alliance: Dublin, Ireland, 2011. [Google Scholar]
- De Weirdt, R.; Possemiers, S.; Vermeulen, G.; Moerdijk-Poortvliet, T.C.; Boschker, H.T.; Verstraete, W.; Van de Wiele, T. Human faecal microbiota display variable patterns of glycerol metabolism. FEMS Microbiol. Ecol. 2010, 74, 601–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duysburgh, C.; Van den Abbeele, P.; Krishnan, K.; Bayne, T.F.; Marzorati, M. A synbiotic concept containing spore-forming Bacillus strains and a prebiotic fiber blend consistently enhanced metabolic activity by modulation of the gut microbiome in vitro. Int. J. Pharm. X 2019, 1, 100021. [Google Scholar] [CrossRef]
- Brown, J.R.M.; Flemer, B.; Joyce, S.A.; Zulquernain, A.; Sheehan, D.; Shanahan, F.; O’Toole, P.W. Changes in microbiota composition, bile and fatty acid metabolism, in successful faecal microbiota transplantation for Clostridioides difficile infection. BMC Gastroenterol. 2018, 18, 131. [Google Scholar] [CrossRef] [Green Version]
- Boon, N.; Top, E.M.; Verstraete, W.; Siciliano, S.D. Bioaugmentation as a tool to protect the structure and function of an activated-sludge microbial community against a 3-chloroaniline shock load. Appl. Environ. Microbiol. 2003, 69, 1511–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furet, J.P.; Firmesse, O.; Gourmelon, M.; Bridonneau, C.; Tap, J.; Mondot, S.; Dore, J.; Corthier, G. Comparative assessment of human and farm animal faecal microbiota using real-time quantitative PCR. FEMS Microbiol. Ecol. 2009, 68, 351–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinttila, T.; Kassinen, A.; Malinen, E.; Krogius, L.; Palva, A. Development of an extensive set of 16S rDNA-targeted primers for quantification of pathogenic and indigenous bacteria in faecal samples by real-time PCR. J. Appl. Microbiol. 2004, 97, 1166–1177. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L. Assessing and improving methods used in operational taxonomic unit-based approaches for 16S rRNA gene sequence analysis. Appl. Environ. Microbiol. 2011, 77, 3219–3226. [Google Scholar] [CrossRef] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandeputte, D.; Kathagen, G.; D′Hoe, K.; Vieira-Silva, S.; Valles-Colomer, M.; Sabino, J.; Wang, J.; Tito, R.Y.; De Commer, L.; Darzi, Y.; et al. Quantitative microbiome profiling links gut community variation to microbial load. Nature 2017, 551, 507–511. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| PC | DC | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| POF 0.6 g/d | POF 1.0 g/d | POF 1.4 g/d | OFO 1.4 g/d | POF 0.6 g/d | POF 1.0 g/d | POF 1.4 g/d | OFO 1.4 g/d | ||||||||||
| Lactobacillus | Lumen | 3.5 × 108 | a | 1.4 × 109 | a | 6.2 × 109 | b | 4.3 × 108 | a | 1.5 × 108 | a | 9.1 × 108 | b,c | 5.6 × 109 | b | 2.8 × 108 | a,c |
| Mucus | 2.4 × 109 | a | 4.0 × 109 | a,b | 3.3 × 109 | b | 1.2 × 109 | a,b | 6.0 × 107 | a | 7.8 × 107 | a,b | 1.3 × 109 | b | 8.7 × 107 | a | |
| Bifidobacterium | Lumen | 4.5 × 109 | a,b | 9.0 × 109 | a | 4.5 × 109 | b | 7.2 × 109 | a | 1.4 × 109 | a | 4.2 × 109 | b | 4.3 × 109 | b | 3.6 × 109 | b |
| Mucus | 5.3 × 109 | a,b | 1.4 × 1010 | a | 9.2 × 109 | b | 1.0 × 1010 | a | 2.3 × 108 | 5.8 × 108 | 1.1 × 109 | 5.6 × 108 | |||||
| Proximal Colon | |||||
|---|---|---|---|---|---|
| POF | OFO | ||||
| C | TR | C | TR | ||
| Actinobacteria | Bifidobacteriaceae | 6.83 | 8.03 | 6.94 | 8.12 |
| Coriobacteriaceae | <LOD | 5.76 | <LOD | 5.10 | |
| Bacteroidetes | Bacteroidaceae | 7.53 | 6.01 | 7.55 | 7.85 |
| Muribaculaceae | <LOD | <LOD | <LOD | 4.37 | |
| Prevotellaceae | 5.04 | 7.65 | 4.73 | 7.12 | |
| Rikenellaceae | 4.32 | 4.20 | 4.34 | 5.41 | |
| Tannerellaceae | 5.64 | 4.67 | 5.92 | 6.58 | |
| Firmicutes | Acidaminococcaceae | 6.53 | 5.36 | 5.20 | 7.25 |
| Enterococcaceae | 3.36 | 5.34 | 3.53 | 7.53 | |
| Erysipelotrichaceae | <LOD | 4.11 | 4.64 | 4.72 | |
| Lachnospiraceae | 7.33 | 5.86 | 7.37 | 7.76 | |
| Lactobacillaceae | 3.45 | 7.78 | 3.51 | 6.32 | |
| Ruminococcaceae | 3.60 | 4.09 | 3.71 | 4.80 | |
| Veillonellaceae | 6.96 | 7.93 | 7.14 | 7.20 | |
| Fusobacteria | Fusobacteriaceae | <LOD | <LOD | 4.21 | 4.89 |
| Proteobacteria | Burkholderiaceae | 5.15 | 5.62 | 5.62 | 6.65 |
| Desulfovibrionaceae | 5.40 | 4.41 | 5.66 | 4.93 | |
| Enterobacteriaceae | 4.91 | 5.99 | 5.27 | 7.50 | |
| Pseudomonadaceae | 4.75 | 4.14 | 5.20 | 4.59 | |
| uncultured | <LOD | <LOD | 3.24 | 4.20 | |
| Synergistetes | Synergistaceae | 4.59 | 3.88 | 3.81 | 4.70 |
| Verrucomicrobia | Akkermansiaceae | <LOD | <LOD | 3.61 | 3.94 |
| Distal Colon | |||||
|---|---|---|---|---|---|
| POF | OFO | ||||
| C | TR | C | TR | ||
| Actinobacteria | Bifidobacteriaceae | 7.39 | 8.26 | 7.45 | 8.13 |
| Coriobacteriaceae | 5.61 | 5.31 | 5.66 | 4.93 | |
| Bacteroidetes | Bacteroidaceae | 8.35 | 8.20 | 8.30 | 8.04 |
| Muribaculaceae | 5.26 | 6.23 | 4.87 | 6.02 | |
| Prevotellaceae | 6.55 | 6.95 | 6.27 | 7.22 | |
| Rikenellaceae | 6.29 | 5.74 | 6.12 | 6.10 | |
| Tannerellaceae | 7.26 | 7.84 | 7.28 | 7.64 | |
| Firmicutes | Acidaminococcaceae | 6.61 | 7.22 | 6.79 | 6.94 |
| Enterococcaceae | 4.39 | 6.11 | 4.34 | 7.88 | |
| Erysipelotrichaceae | 5.64 | 4.86 | 5.65 | <LOD | |
| Lachnospiraceae | 8.23 | 8.28 | 8.16 | 8.27 | |
| Lactobacillaceae | 4.53 | 8.25 | 4.45 | 6.76 | |
| Ruminococcaceae | 6.64 | 6.12 | 6.74 | 5.90 | |
| Veillonellaceae | 7.37 | 8.04 | 7.34 | 7.36 | |
| Fusobacteria | Fusobacteriaceae | 5.62 | 6.34 | 5.70 | 7.66 |
| Proteobacteria | Burkholderiaceae | 6.48 | 6.73 | 6.52 | 6.31 |
| Desulfovibrionaceae | 6.58 | 6.79 | 6.76 | 6.94 | |
| Enterobacteriaceae | 5.42 | 6.36 | 5.45 | 7.63 | |
| Pseudomonadaceae | 6.57 | 5.52 | 6.44 | 6.48 | |
| uncultured | 5.89 | 6.51 | 6.05 | 6.33 | |
| Synergistetes | Synergistaceae | 6.93 | 7.52 | 7.16 | 7.27 |
| Verrucomicrobia | Akkermansiaceae | 7.24 | 7.67 | 7.41 | 7.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duysburgh, C.; Van den Abbeele, P.; Kamil, A.; Fleige, L.; De Chavez, P.J.; Chu, Y.; Barton, W.; O’Sullivan, O.; Cotter, P.D.; Quilter, K.; et al. In vitro–in vivo Validation of Stimulatory Effect of Oat Ingredients on Lactobacilli. Pathogens 2021, 10, 235. https://doi.org/10.3390/pathogens10020235
Duysburgh C, Van den Abbeele P, Kamil A, Fleige L, De Chavez PJ, Chu Y, Barton W, O’Sullivan O, Cotter PD, Quilter K, et al. In vitro–in vivo Validation of Stimulatory Effect of Oat Ingredients on Lactobacilli. Pathogens. 2021; 10(2):235. https://doi.org/10.3390/pathogens10020235
Chicago/Turabian StyleDuysburgh, Cindy, Pieter Van den Abbeele, Alison Kamil, Lisa Fleige, Peter John De Chavez, YiFang Chu, Wiley Barton, Orla O’Sullivan, Paul D. Cotter, Karina Quilter, and et al. 2021. "In vitro–in vivo Validation of Stimulatory Effect of Oat Ingredients on Lactobacilli" Pathogens 10, no. 2: 235. https://doi.org/10.3390/pathogens10020235
APA StyleDuysburgh, C., Van den Abbeele, P., Kamil, A., Fleige, L., De Chavez, P. J., Chu, Y., Barton, W., O’Sullivan, O., Cotter, P. D., Quilter, K., Joyce, S. A., Murphy, M., DunnGalvin, G., Dinan, T. G., & Marzorati, M. (2021). In vitro–in vivo Validation of Stimulatory Effect of Oat Ingredients on Lactobacilli. Pathogens, 10(2), 235. https://doi.org/10.3390/pathogens10020235