
Recognition Pattern of the Fasciola hepatica Excretome/Secretome during the Course of an Experimental Infection in Sheep by 2D Immunoproteomics

 , ,
, , 
Abstract
:1. Introduction
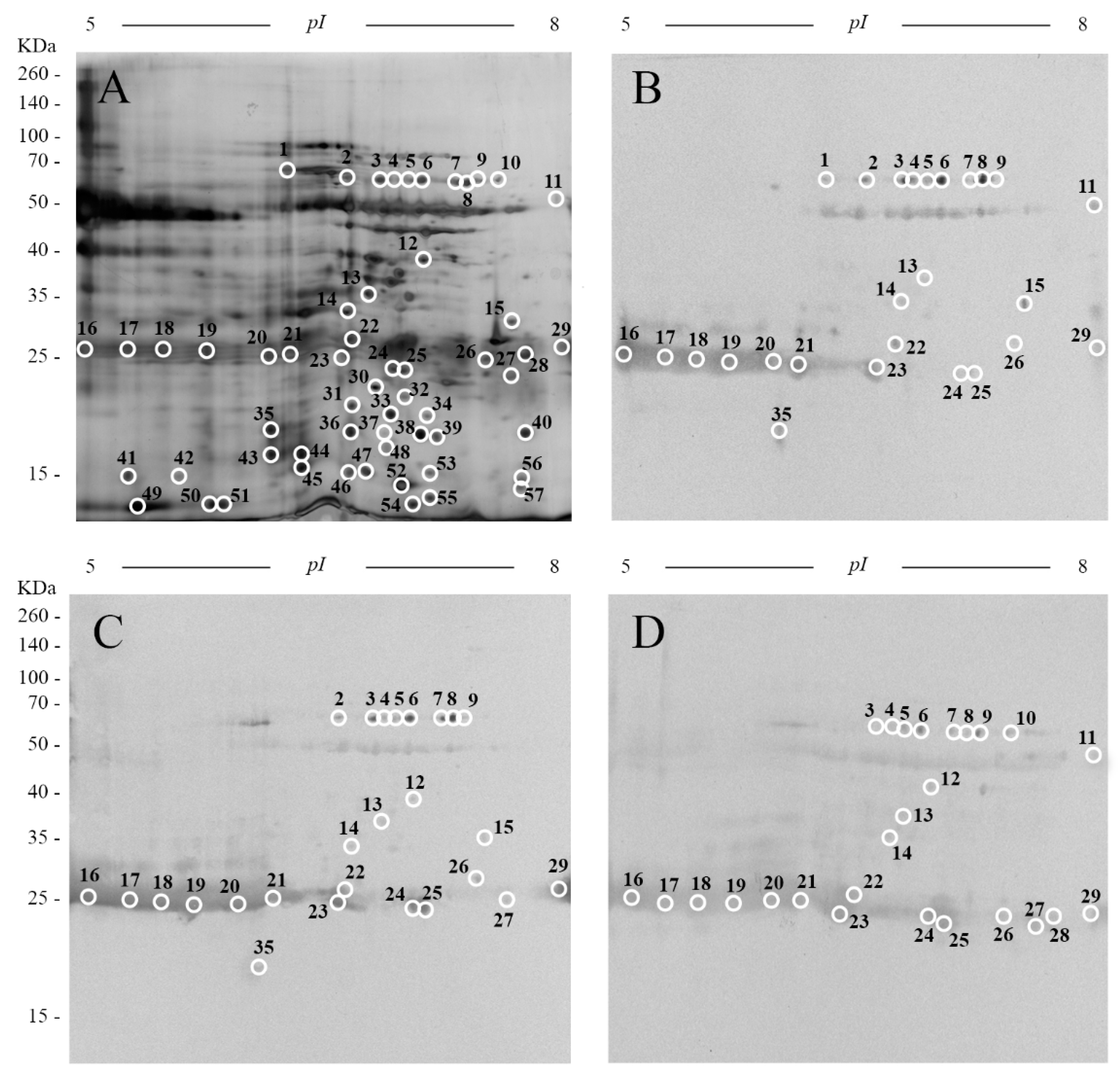
2. Results and Discussion
3. Materials and Methods
3.1. Collection of a F. hepatica ES Extract
3.2. Obtention of Sheep Serum Samples at Different Infection Times
3.3. Protein Separation by 2D-PAGE
3.4. Immunoblot Assays
3.5. Image Acquisition and Spot Selection
3.6. Spot Identification by Mass Spectrometry
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siles-Lucas, M.; Becerro-Recio, D.; Serrat, J.; González-Miguel, J. Fascioliasis and fasciolopsiasis: Current knowledge and future trends. Res. Vet. Sci. 2021, 134, 27–35. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Bargues, M.D.; Esteban, J.G. Human fasciolosis. In Fasciolosis; Dalton, J.P., Ed.; CAB International Publishing: Wallingford, UK, 1999; pp. 411–434. ISBN 0-85199-260-9. [Google Scholar]
- Moazeni, M.; Ahmadi, A. Controversial aspects of the life cycle of Fasciola hepatica. Exp. Parasitol. 2016, 169, 81–89. [Google Scholar] [CrossRef]
- González-Miguel, J.; Becerro-Recio, D.; Siles-Lucas, M. Insights into Fasciola hepatica Juveniles: Crossing the Fasciolosis Rubicon. Trends Parasitol. 2021, 37, 35–47. [Google Scholar] [CrossRef]
- Harrington, D.; Lamberton, P.H.L.; McGregor, A. Human liver flukes. Lancet Gastroenterol. Hepatol. 2017, 2, 680–689. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Valero, M.A.; Bargues, M.D. Fascioliasis. Adv. Exp. Med. Biol. 2019, 1154, 71–103. [Google Scholar] [CrossRef] [PubMed]
- Behm, C.A.; Sangster, N.C. Pathology, pathophysiology and clinical aspects. In Fasciolosis; Dalton, J.P., Ed.; CAB International Publishing: Wallingford, UK, 1999; pp. 185–224. ISBN 0-85199-260-9. [Google Scholar]
- Madhumitha, R.; Gohel, S.; Vishwanathan, L.; Gopalakrishnan, R. Liver Lesions, Fever and Eosinophilia Caused by Fasciola hepatica in a 15-year-old Girl. Indian J. Pediatr. 2015, 82, 967–968. [Google Scholar] [CrossRef] [PubMed]
- Arslan, F.; Batirel, A.; Samasti, M.; Tabak, F.; Mert, A.; Özer, S. Fascioliasis: 3 cases with three different clinical presentations. Turk. J. Gastroenterol. 2012, 23, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Alvarez Rojas, C.A.; Jex, A.R.; Gasser, R.B.; Scheerlinck, J.P. Techniques for the diagnosis of Fasciola infections in animals: Room for improvement. Adv. Parasitol. 2014, 85, 65–107. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, K.; Zhang, H.; Sabir, A.J.; Abbas, R.Z.; Ijaz, M.; Durrani, A.Z.; Saleem, M.H.; Ur Rehman, M.; Iqbal, M.K.; Wang, Y.; et al. A review on epidemiology, global prevalence and economical losses of fasciolosis in ruminants. Microb. Pathog. 2017, 109, 253–262. [Google Scholar] [CrossRef]
- Keiser, J.; Utzinger, J. Emerging foodborne trematodiasis. Emerg. Infect. Dis. 2005, 11, 1507–1514. [Google Scholar] [CrossRef]
- Fürst, T.; Keiser, J.; Utzinger, J. Global burden of human food-borne trematodiasis: A systematic review and meta-analysis. Lancet Infect. Dis. 2012, 12, 210–221. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Bargues, M.D.; Valero, M.A. Diagnosis of human fascioliasis by stool and blood techniques: Update for the present global scenario. Parasitology 2014, 141, 1918–1946. [Google Scholar] [CrossRef] [PubMed]
- Radfar, M.H.; Nourollahi-Fard, S.R.; Mohammadyari, N. Bovine fasciolosis: Prevalence, relationship between faecal egg count and worm burden and its economic impact due to liver condemnation at Rudsar abattoir, Northern Iran. J. Parasit. Dis. 2015, 39, 522–525. [Google Scholar] [CrossRef] [Green Version]
- Cwiklinski, K.; O’Neill, S.M.; Donnelly, S.; Dalton, J.P. A prospective view of animal and human Fasciolosis. Parasite Immunol. 2016, 38, 558–568. [Google Scholar] [CrossRef] [Green Version]
- Sarkari, B.; Khabisi, S.A. Immunodiagnosis of Human Fascioliasis: An Update of Concepts and Performances of the Serological Assays. J. Clin. Diagn. Res. 2017, 11, OE05–OE10. [Google Scholar] [CrossRef] [PubMed]
- Webb, C.M.; Cabada, M.M. Recent developments in the epidemiology, diagnosis, and treatment of Fasciola infection. Curr. Opin. Infect. Dis. 2018, 31, 409–414. [Google Scholar] [CrossRef]
- Keiser, J.; Engels, D.; Büscher, G.; Utzinger, J. Triclabendazole for the treatment of fascioliasis and paragonimiasis. Expert. Opin. Investig. Drugs. 2005, 14, 1513–1526. [Google Scholar] [CrossRef]
- Brennan, G.P.; Fairweather, I.; Trudgett, A.; Hoey, E.; McCoy; McConville, M.; Meaney, M.; Robinson, M.; McFerran, N.; Ryan, L.; et al. Understanding triclabendazole resistance. Exp. Mol. Pathol. 2007, 82, 104–109. [Google Scholar] [CrossRef]
- Kamaludeen, J.; Graham-Brown, J.; Stephens, N.; Miller, J.; Howell, A.; Beesley, N.J.; Hodgkinson, J.; Learmount, J.; Williams, D. Lack of efficacy of triclabendazole against Fasciola hepatica is present on sheep farms in three regions of England, and Wales. Vet. Rec. 2019, 184, 502. [Google Scholar] [CrossRef] [Green Version]
- Power, C.; Danaher, M.; Sayers, R.; O’Brien, B.; Clancy, C.; Furey, A.; Jordan, K. Investigation of the migration of triclabendazole residues to milk products manufactured from bovine milk, and stability therein, following lactating cow treatment. J. Dairy Sci. 2013, 96, 6223–6232. [Google Scholar] [CrossRef] [Green Version]
- Dalton, J.P.; Robinson, M.W.; Mulcahy, G.; O’Neill, S.M.; Donnelly, S. Immunomodulatory molecules of Fasciola hepatica: Candidates for both vaccine and immunotherapeutic development. Vet. Parasitol. 2013, 195, 272–285. [Google Scholar] [CrossRef]
- Meemon, K.; Sobhon, P. Juvenile-specific cathepsin proteases in Fasciola spp.: Their characteristics and vaccine efficacies. Parasitol. Res. 2015, 114, 2807–2813. [Google Scholar] [CrossRef]
- McManus, D.P. Recent Progress in the Development of Liver Fluke and Blood Fluke Vaccines. Vaccines 2020, 8, 553. [Google Scholar] [CrossRef]
- Dominguez, M.F.; González-Miguel, J.; Carmona, C.; Dalton, J.P.; Cwiklinski, K.; Tort, J.; Siles-Lucas, M. Low allelic diversity in vaccine candidates genes from different locations sustain hope for Fasciola hepatica immunization. Vet. Parasitol. 2018, 258, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Harnett, W. Secretory products of helminth parasites as immunomodulators. Mol. Biochem. Parasitol. 2014, 195, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.; Shiels, J.; Taggart, C.C.; Dalton, J.P.; Weldon, S. Fasciola hepatica-Derived Molecules as Regulators of the Host Immune Response. Front. Immunol. 2020, 11, 2182. [Google Scholar] [CrossRef]
- Cwiklinski, K.; Dalton, J.P. Advances in Fasciola hepatica research using ’omics’ technologies. Int. J. Parasitol. 2018, 48, 321–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corral-Ruiz, G.M.; Sánchez-Torres, L.E. Fasciola hepatica-derived molecules as potential immunomodulators. Acta Trop. 2020, 210, 105548. [Google Scholar] [CrossRef] [PubMed]
- Cwiklinski, K.; Dalton, J.P.; Dufresne, P.J.; La Course, J.; Williams, D.J.; Hodgkinson, J.; Paterson, S. The Fasciola hepatica genome: Gene duplication and polymorphism reveals adaptation to the host environment and the capacity for rapid evolution. Genome Biol. 2015, 16, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.W.; Tort, J.F.; Lowther, J.; Donnelly, S.M.; Wong, E.; Xu, W.; Stack, C.M.; Padula, M.; Herbert, B.; Dalton, J.P. Proteomics and phylogenetic analysis of the cathepsin L protease family of the helminth pathogen Fasciola hepatica: Expansion of a repertoire of virulence-associated factors. Mol. Cell. Proteomics. 2008, 7, 1111–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cwiklinski, K.; Donnelly, S.; Drysdale, O.; Jewhurst, H.; Smith, D.; De Marco Verissimo, C.; Pritsch, I.C.; O’Neill, S.; Dalton, J.P.; Robinson, M.W. The cathepsin-like cysteine peptidases of trematodes of the genus Fasciola. Adv. Parasitol. 2019, 104, 113–164. [Google Scholar] [CrossRef] [PubMed]
- González-Miguel, J.; Valero, M.A.; Reguera-Gomez, M.; Mas-Bargues, C.; Bargues, M.D.; Simón, F.; Mas-Coma, S. Numerous Fasciola plasminogen-binding proteins may underlie blood-brain barrier leakage and explain neurological disorder complexity and heterogeneity in the acute and chronic phases of human fascioliasis. Parasitology 2019, 146, 284–298. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, B.; Hecht, H.J.; Flohé, L. Peroxiredoxins. Biol. Chem. 2002, 383, 347–364. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef]
- Donnelly, S.; O’Neill, S.M.; Sekiya, M.; Mulcahy, G.; Dalton, J.P. Thioredoxin peroxidase secreted by Fasciola hepatica induces the alternative activation of macrophages. Infect. Immun. 2005, 73, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Chemale, G.; Morphew, R.; Moxon, J.V.; Morassuti, A.L.; Lacourse, E.J.; Barrett, J.; Johnston, D.A.; Brophy, P.M. Proteomic analysis of glutathione transferases from the liver fluke parasite, Fasciola hepatica. Proteomics 2006, 6, 6263–6273. [Google Scholar] [CrossRef] [PubMed]
- Spithill, T.W.; Smooker, P.M.; Copeman, D.B. Fasciola gigantica: Epidemiology, control, immunology and molecular biology. In Fasciolosis; Dalton, J.P., Ed.; CAB International Publishing: Wallingford, UK, 1999; pp. 465–525. ISBN 0-85199-260-9. [Google Scholar]
- Toh, S.Q.; Gobert, G.N.; Malagón Martínez, D.; Jones, M.K. Haem uptake is essential for egg production in the haematophagous blood fluke of humans, Schistosoma mansoni. FEBS J. 2015, 282, 3632–3646. [Google Scholar] [CrossRef] [PubMed]
- Costa-Paiva, E.M.; Coates, C.J. Recent Insights into the Diversity and Evolution of Invertebrate Hemerythrins and Extracellular Globins. Subcell. Biochem. 2020, 94, 251–273. [Google Scholar] [CrossRef]
- Dewilde, S.; Ioanitescu, A.I.; Kiger, L.; Gilany, K.; Marden, M.C.; Van Doorslaer, S.; Vercruysse, J.; Pesce, A.; Nardini, M.; Bolognesi, M.; et al. The hemoglobins of the trematodes Fasciola hepatica and Paramphistomum epiclitum: A molecular biological, physico-chemical, kinetic, and vaccination study. Protein Sci. 2008, 17, 1653–1662. [Google Scholar] [CrossRef] [Green Version]
- Cabán-Hernández, K.; Gaudier, J.F.; Espino, A.M. Characterization and differential expression of a ferritin protein from Fasciola hepatica. Mol. Biochem. Parasitol. 2012, 182, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Jefferies, J.R.; Campbell, A.M.; van Rossum, A.J.; Barrett, J.; Brophy, P.M. Proteomic analysis of Fasciola hepatica excretory-secretory products. Proteomics 2001, 1, 1128–1132. [Google Scholar] [CrossRef]
- Morphew, R.M.; Wright, H.A.; LaCourse, E.J.; Woods, D.J.; Brophy, P.M. Comparative proteomics of excretory-secretory proteins released by the liver fluke Fasciola hepatica in sheep host bile and during in vitro culture ex host. Mol. Cell. Proteomics 2007, 6, 963–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, T.R.; Ainsworth, S.; Armstrong, S.; Hodgkinson, J.; Williams, D. Differences in the antibody response to adult Fasciola hepatica excretory/secretory products in experimentally and naturally infected cattle and sheep. Vet. Parasitol. 2021, 289, 109321. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Spot Number | Accession Code | Protein Definition | Sequence Coverage (%) | Molecular Function | Biological Process | Recognition Time (Weeks PI) |
|---|---|---|---|---|---|---|
| 10 | A8E0R8 | Thioredoxin glutathione reductase | 22.68 | oxidoreductase | cell redox homeostasis | 12 |
| A0A4E0R242 | Lysosomal Pro-X carboxypeptidase | 10.03 | serine-type peptidase | - | 12 | |
| 14 | A7UNB2 | Cathepsin B | 10.5 | cysteine-type endopeptidase | regulation of catalytic activity | 4–8–12 |
| 16 | Q24940 | Cathepsin L | 55.95 | cysteine-type endopeptidase | proteolysis | 4–8–12 |
| A0A4E0RY97 | Fimbrin, putative | 4.23 | actin filament binding | actin filament bundle assembly | 4–8–12 | |
| 19 | Q24940 | Cathepsin L | 56.75 | cysteine-type endopeptidase | proteolysis | 4–8–12 |
| 23 | Q24940 | Cathepsin L | 46.43 | cysteine-type endopeptidase | proteolysis | 4–8–12 |
| P91883 | Peroxiredoxin | 16.05 | oxidoreductase | cell redox homeostasis | 4–8–12 | |
| 24 | Q24940 | Cathepsin L | 12.5 | cysteine-type endopeptidase | proteolysis | 4–8–12 |
| P91883 | Peroxiredoxin | 11.52 | oxidoreductase | cell redox homeostasis | 4–8–12 | |
| A4V9Q5 | Calmodulin | 19.46 | calcium ion binding | calcium-mediated signaling | 4–8–12 | |
| 25 | E3UTT4 | Glutathione S-transferase | 36.19 | transferase | - | 4–8–12 |
| P91883 | Peroxiredoxin | 29.89 | oxidoreductase | cell redox homeostasis | 4–8–12 | |
| 26 | E3UTT4 | Glutathione S-transferase | 20.9 | transferase | - | 4–8–12 |
| P91883 | Peroxiredoxin | 22.63 | oxidoreductase | cell redox homeostasis | 4–8–12 | |
| A0A2H1BSW4 | Proteasome subunit alpha type | 27.66 | threonine-type endopeptidase | ubiquitin-dependent protein catabolic process | 4–8–12 | |
| 27 | P91883 | Peroxiredoxin | 39.92 | oxidoreductase | cell redox homeostasis | 8–12 |
| 28 | Q24940 | Cathepsin L | 12.5 | cysteine-type endopeptidase | proteolysis | 12 |
| P91883 | Peroxiredoxin | 16.46 | oxidoreductase | cell redox homeostasis | 12 | |
| 33 | A0A4E0R9N4 | C2H2-type domain-containing protein | 7.96 | nucleic acid binding | regulation of transcription | No recognition |
| 35 | A0A4E0RV44 | Epididymal secretory protein E1 | 10.82 | - | intracellular cholesterol transport | 4–8 |
| 44 | A0A4E0RV44 | Epididymal secretory protein E1 | 7.89 | - | intracellular cholesterol transport | No recognition |
| A0A4E0RD35 | Ferritin | 21.05 | ferric iron binding | cellular iron ion homeostasis | No recognition | |
| 47 | Q9XY94 | Superoxide dismutase [Cu-Zn] | 33.77 | oxidoreductase | removal of superoxide radicals | No recognition |
| 55 | A0A2H1CJ88 | Globin-3 | 9.59 | heme binding | oxygen transport | No recognition |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becerro-Recio, D.; González-Miguel, J.; Ucero, A.; Sotillo, J.; Martínez-Moreno, Á.; Pérez-Arévalo, J.; Cwiklinski, K.; Dalton, J.P.; Siles-Lucas, M. Recognition Pattern of the Fasciola hepatica Excretome/Secretome during the Course of an Experimental Infection in Sheep by 2D Immunoproteomics. Pathogens 2021, 10, 725. https://doi.org/10.3390/pathogens10060725
Becerro-Recio D, González-Miguel J, Ucero A, Sotillo J, Martínez-Moreno Á, Pérez-Arévalo J, Cwiklinski K, Dalton JP, Siles-Lucas M. Recognition Pattern of the Fasciola hepatica Excretome/Secretome during the Course of an Experimental Infection in Sheep by 2D Immunoproteomics. Pathogens. 2021; 10(6):725. https://doi.org/10.3390/pathogens10060725
Chicago/Turabian StyleBecerro-Recio, David, Javier González-Miguel, Alberto Ucero, Javier Sotillo, Álvaro Martínez-Moreno, José Pérez-Arévalo, Krystyna Cwiklinski, John P. Dalton, and Mar Siles-Lucas. 2021. "Recognition Pattern of the Fasciola hepatica Excretome/Secretome during the Course of an Experimental Infection in Sheep by 2D Immunoproteomics" Pathogens 10, no. 6: 725. https://doi.org/10.3390/pathogens10060725
APA StyleBecerro-Recio, D., González-Miguel, J., Ucero, A., Sotillo, J., Martínez-Moreno, Á., Pérez-Arévalo, J., Cwiklinski, K., Dalton, J. P., & Siles-Lucas, M. (2021). Recognition Pattern of the Fasciola hepatica Excretome/Secretome during the Course of an Experimental Infection in Sheep by 2D Immunoproteomics. Pathogens, 10(6), 725. https://doi.org/10.3390/pathogens10060725