
Successful Eradication of Porcine Epidemic Diarrhea in an Enzootically Infected Farm: A Two-Year Follow-Up Study


Abstract
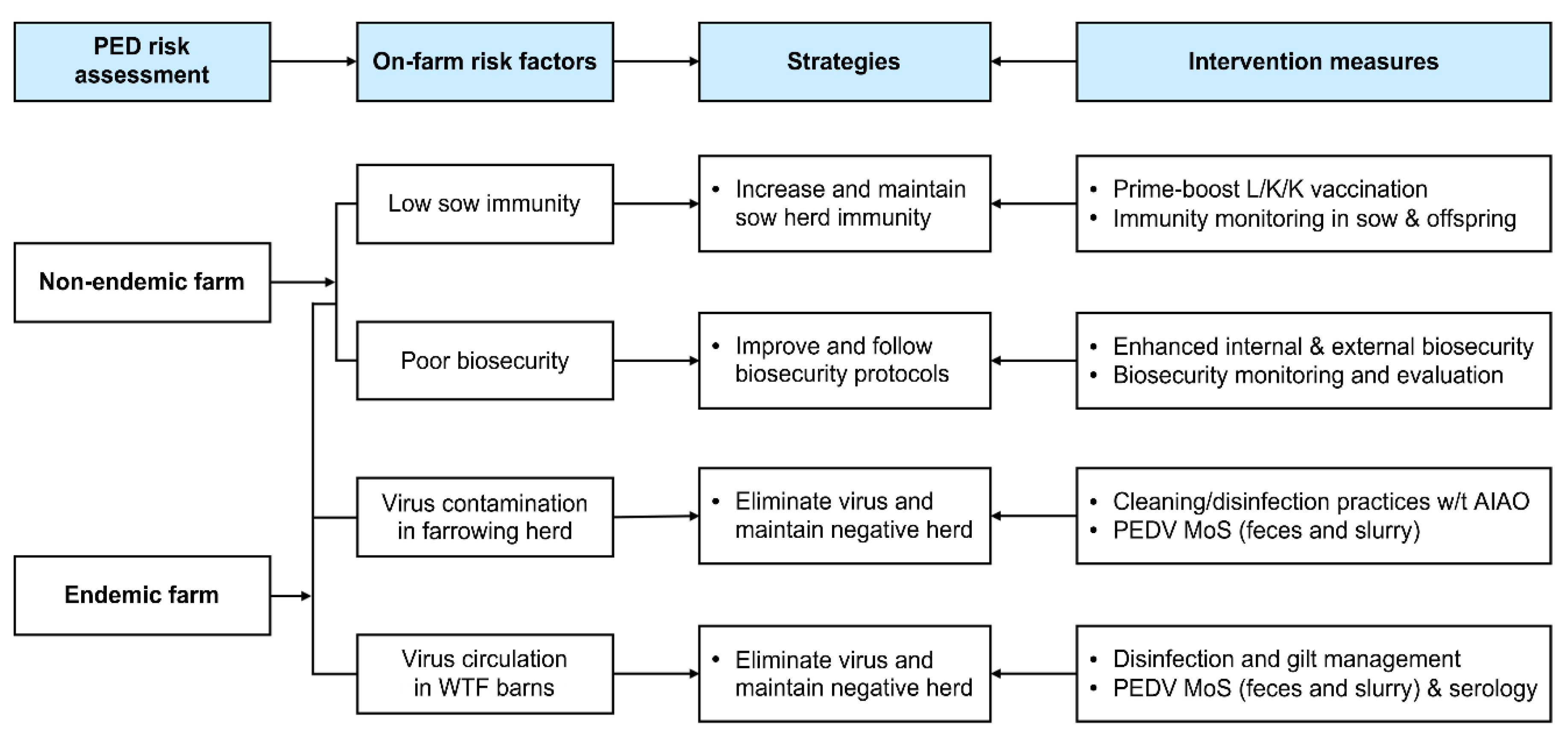
:1. Introduction
2. Results
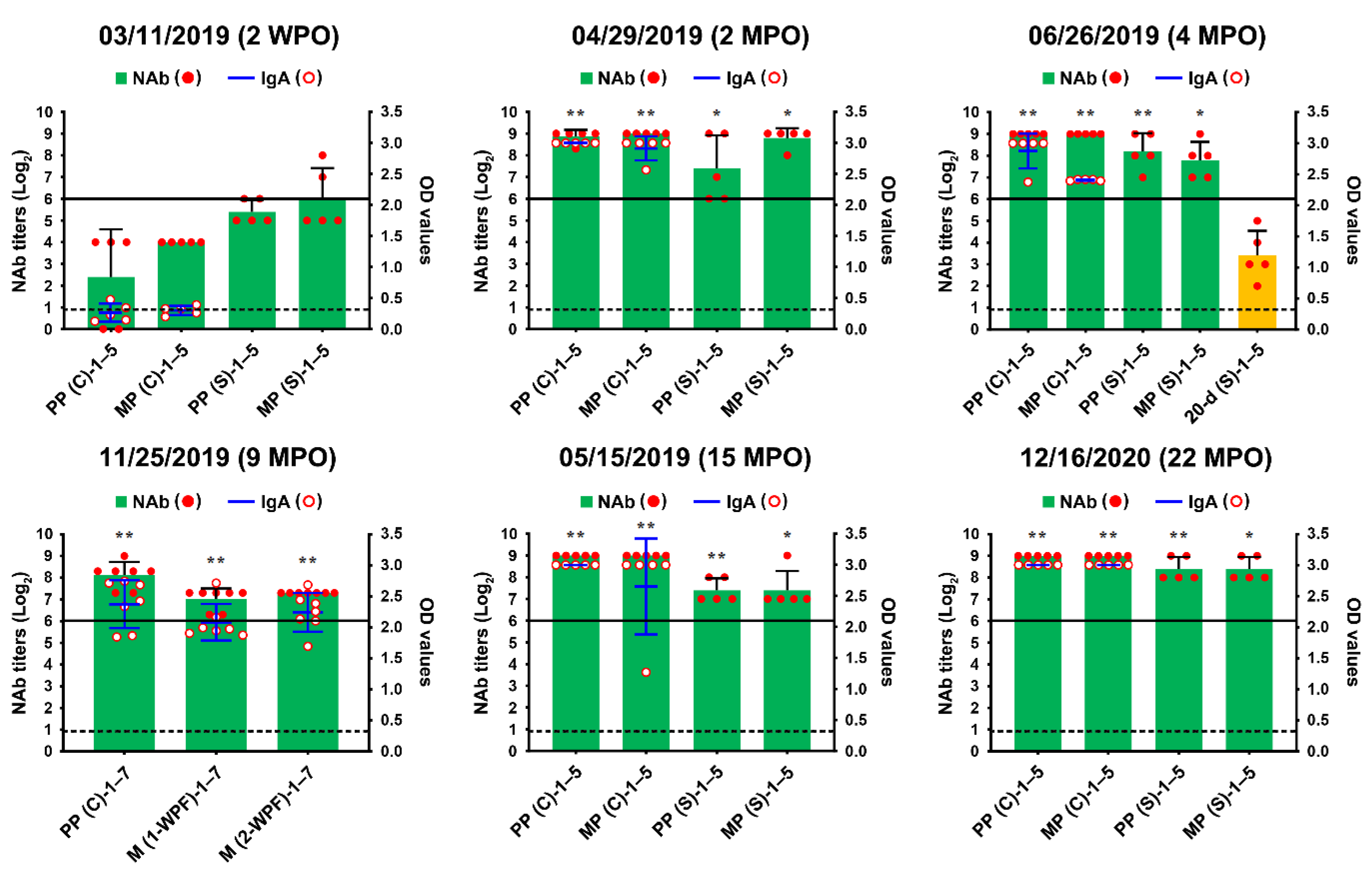
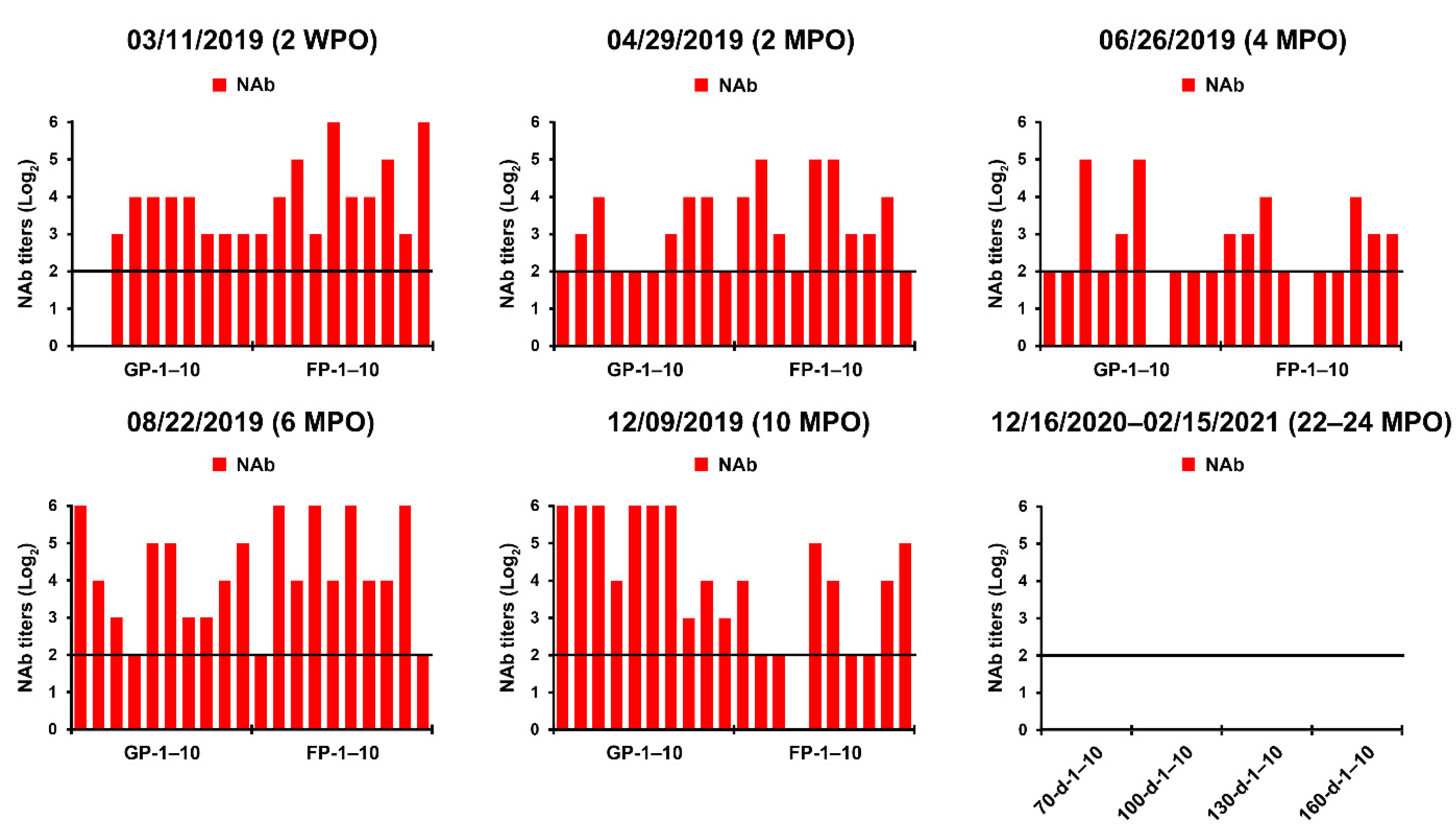
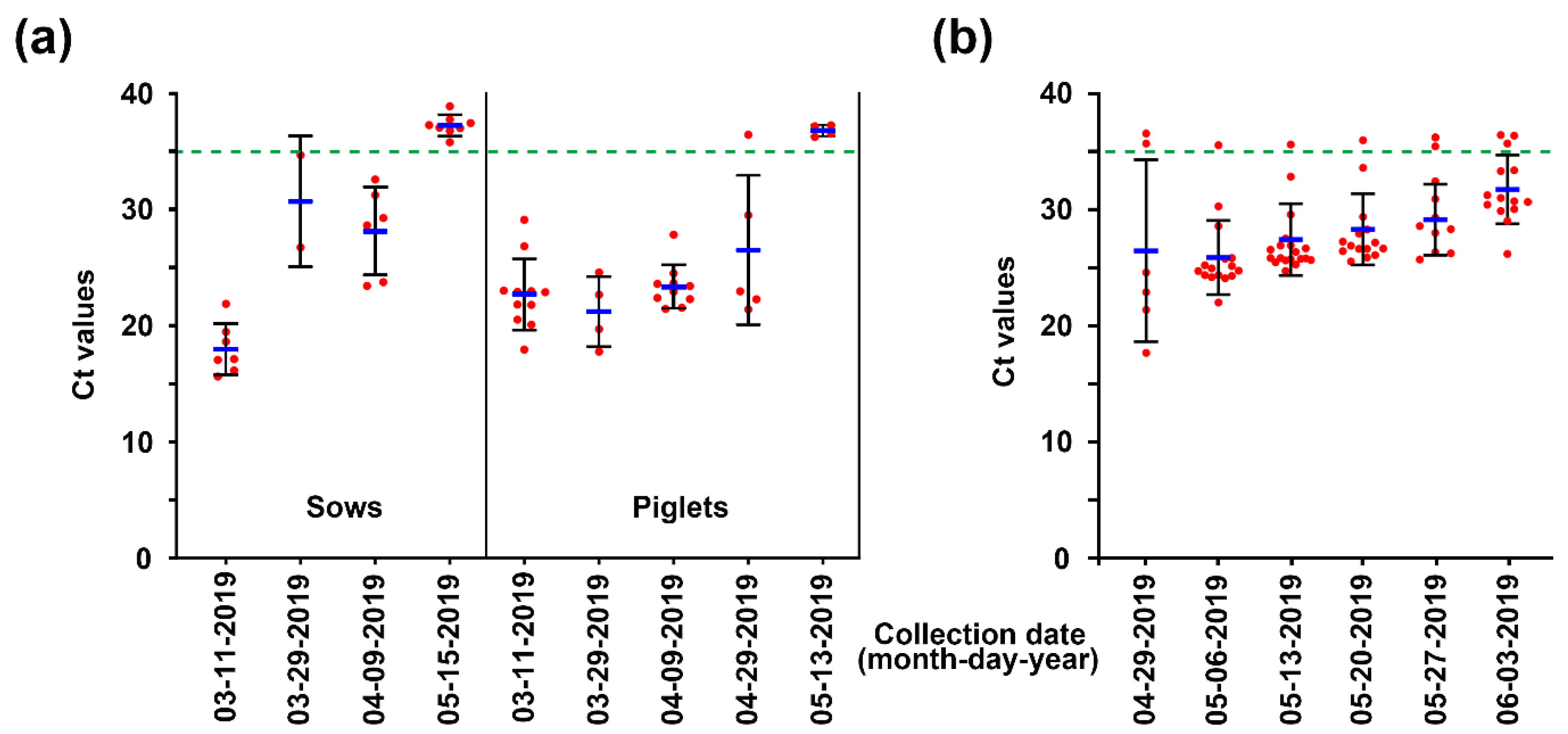
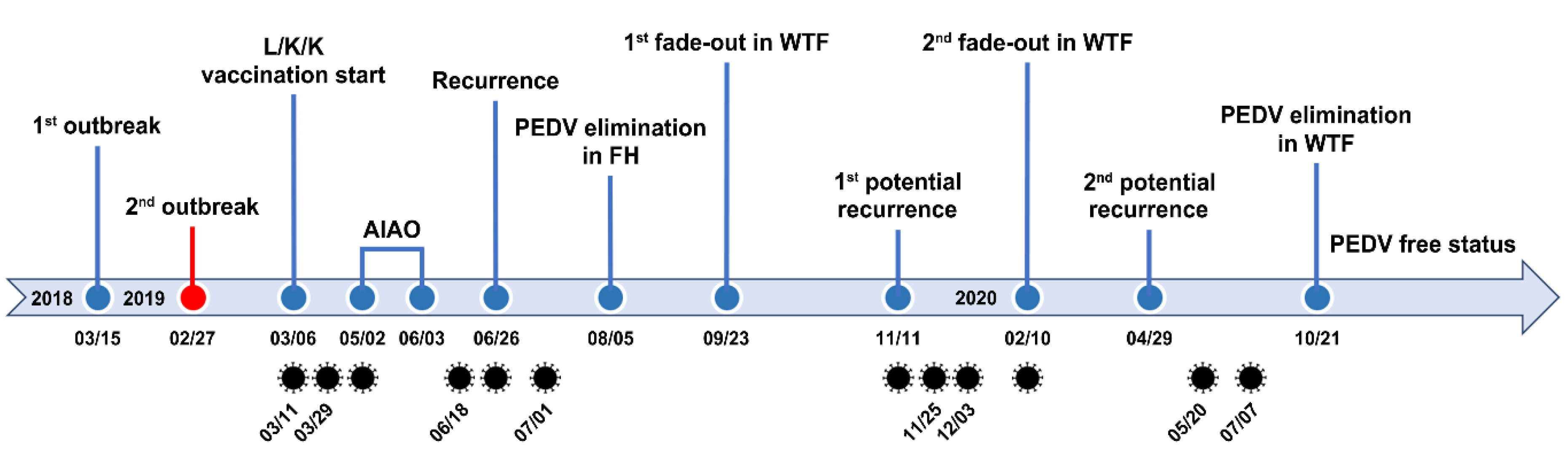
2.1. PEDV Monitoring and Surveillance Investigations
2.2. Application of Two-Track Strategies for PED Eradication
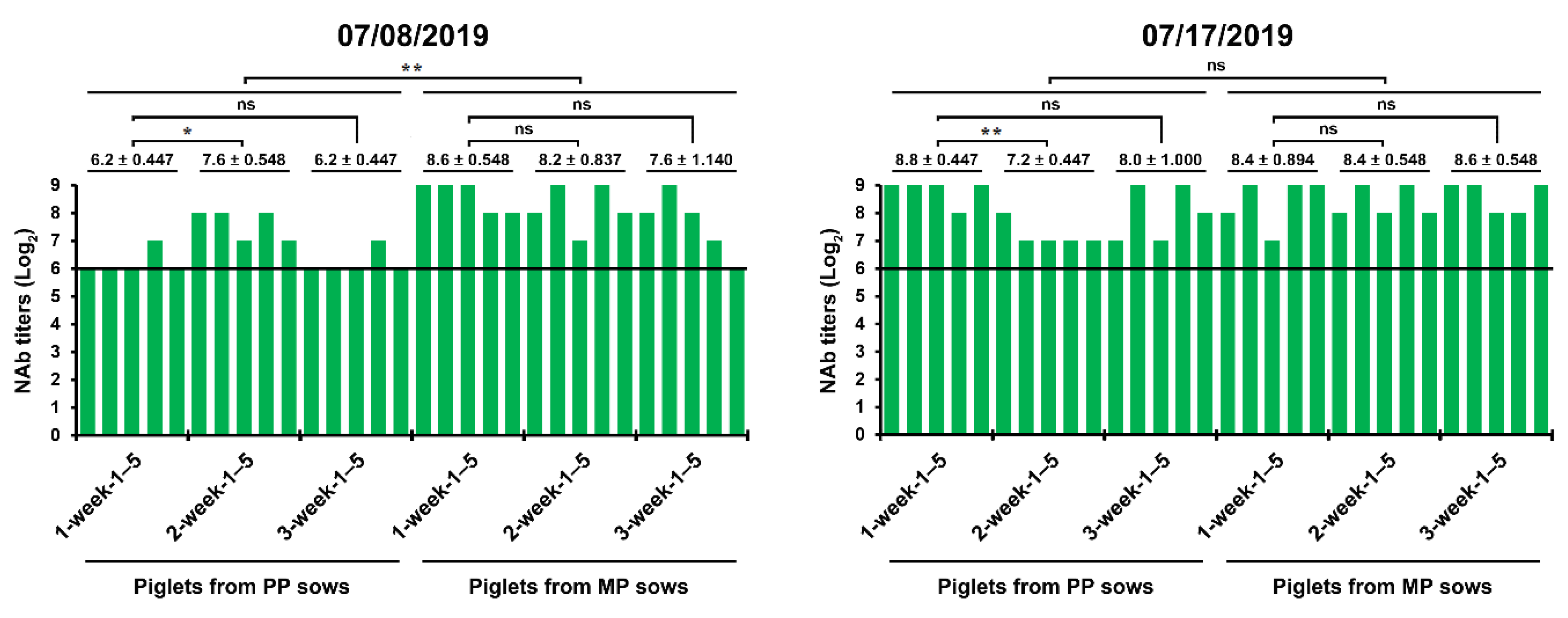
2.3. Recurrent Infection in the Farrowing House on 18 June 2019
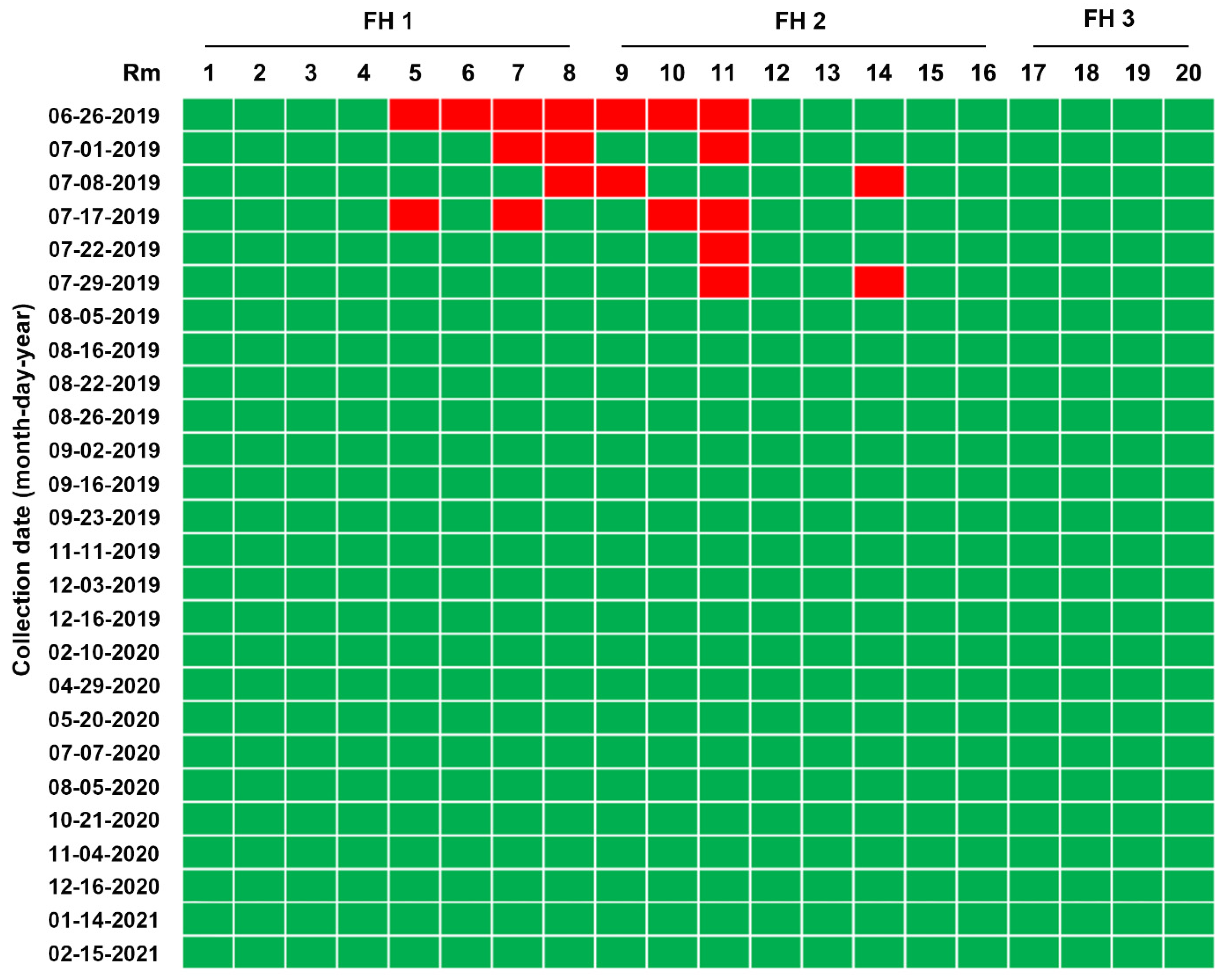
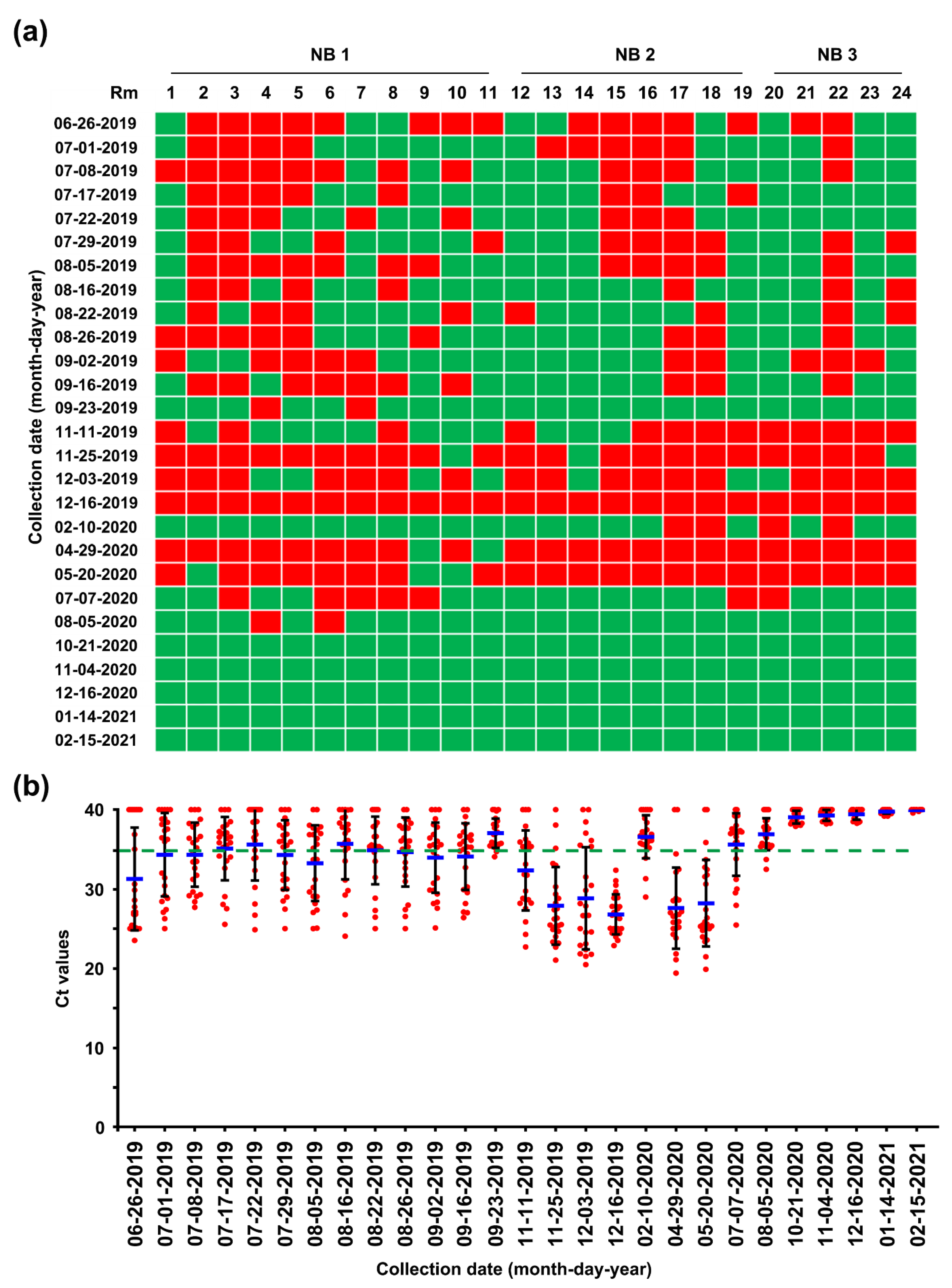
2.4. First Potential Recurrence in the Nursery Barn in November 2019
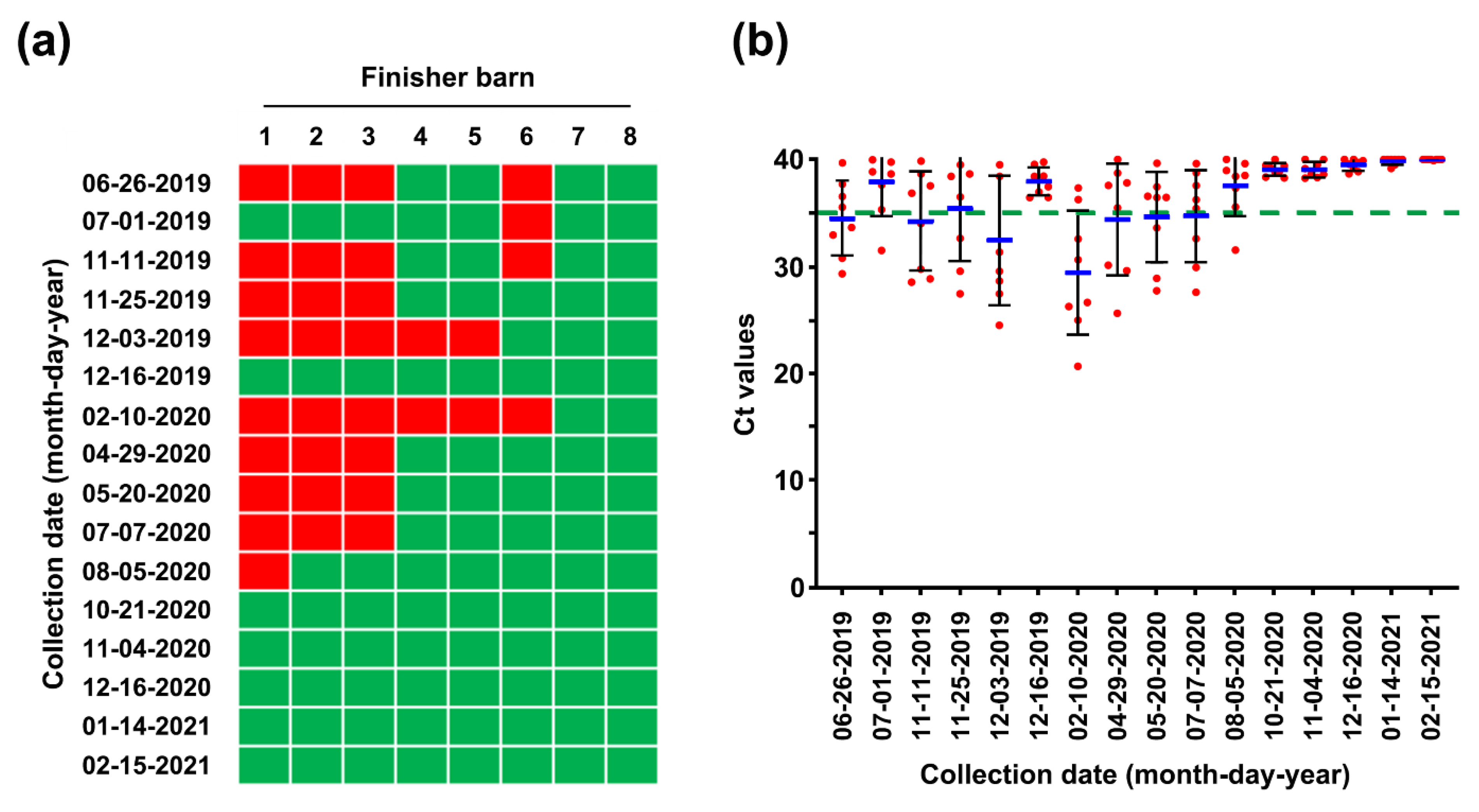
2.5. Second Potential Recurrence in the Nursery Barn in April 2020
2.6. Eradication of PEDV from This Farm
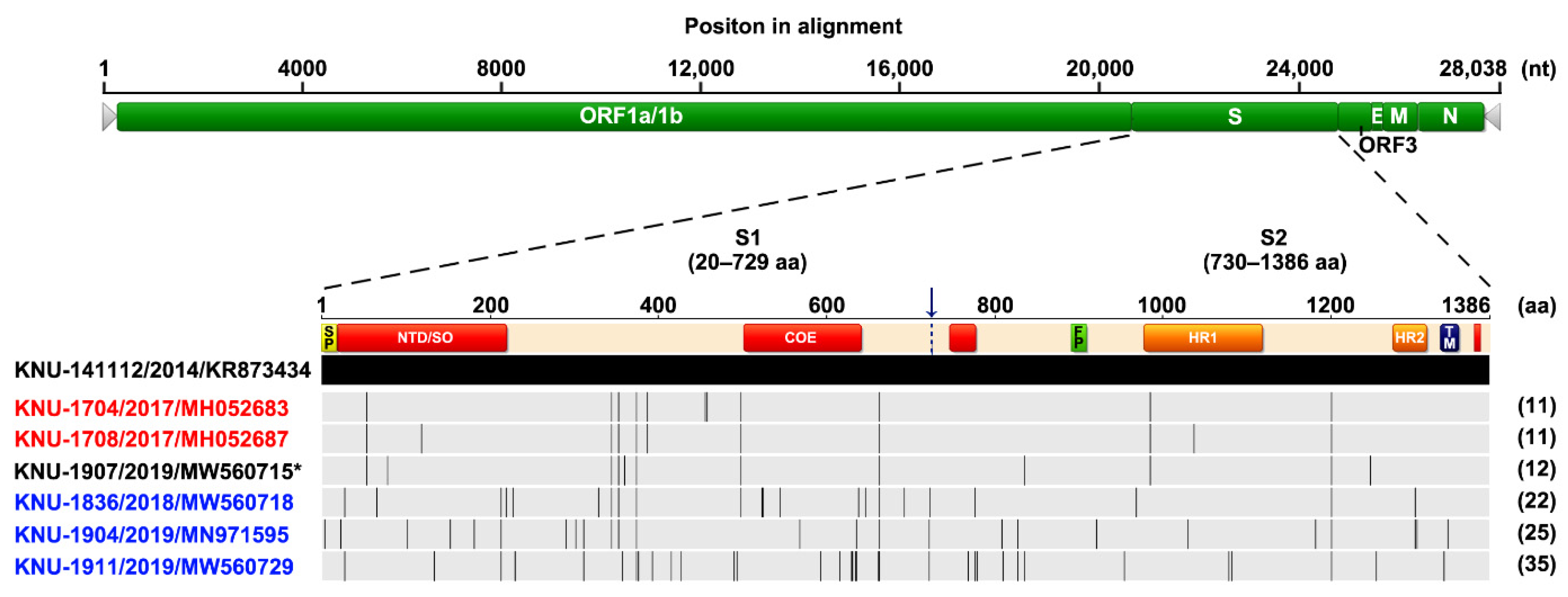
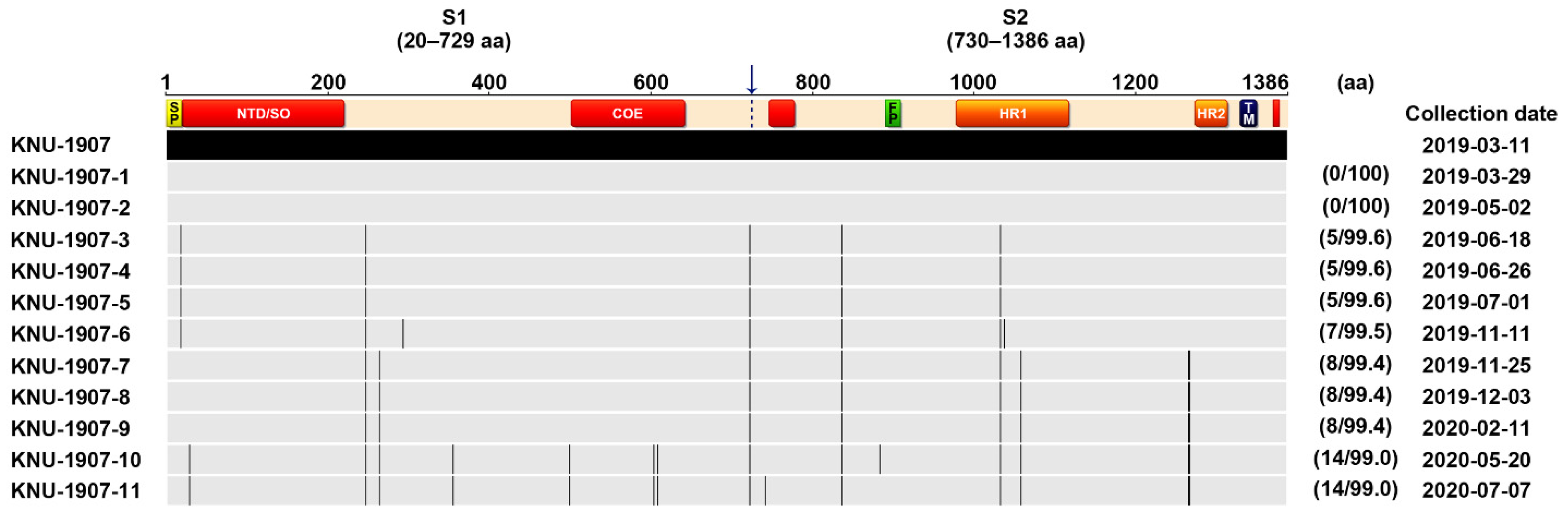
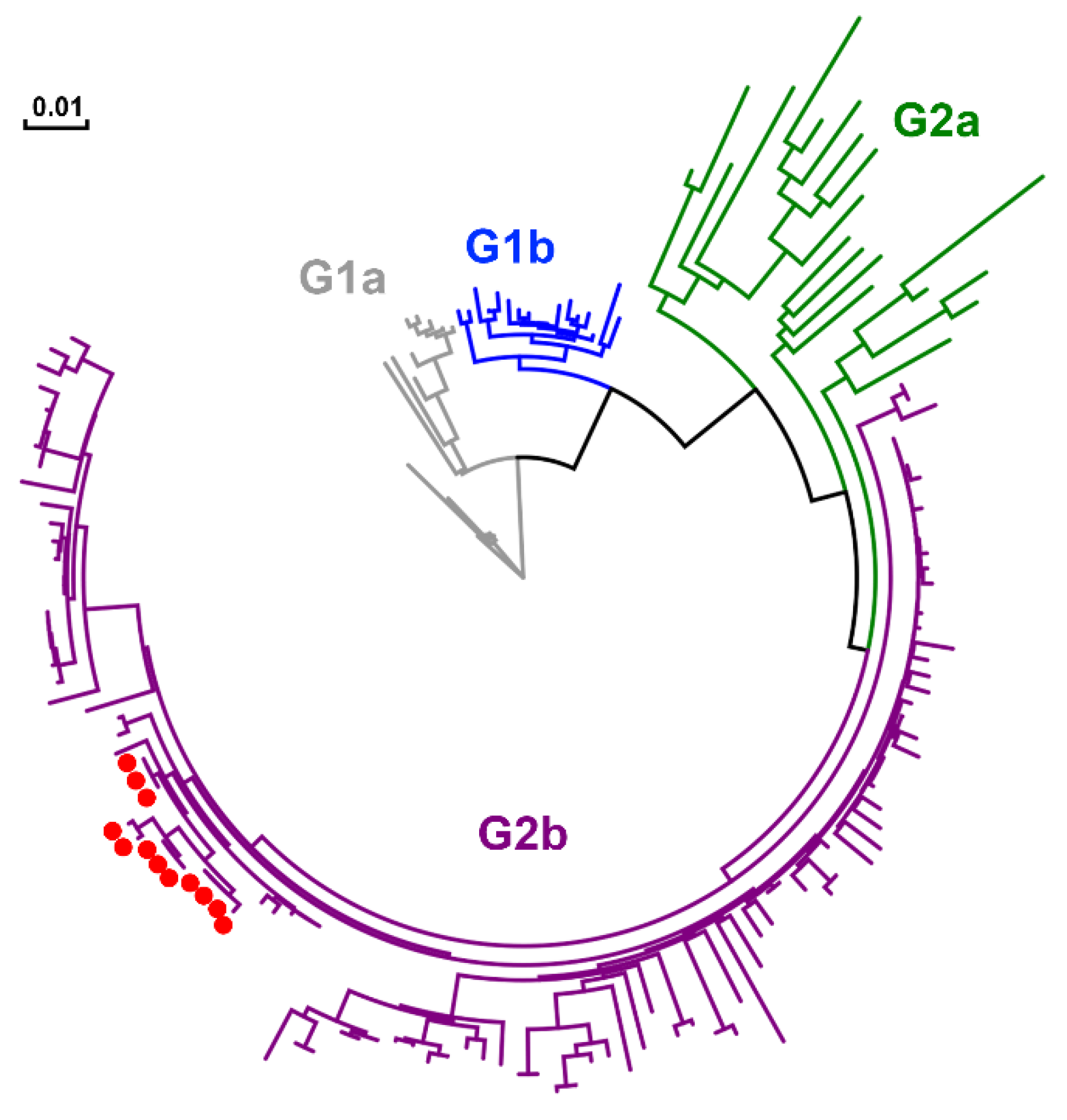
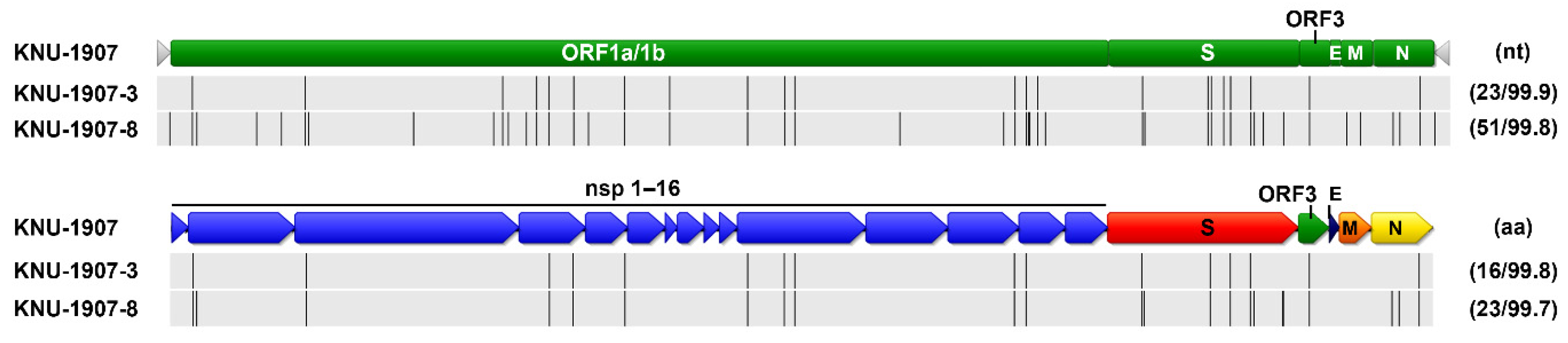
2.7. Genetic Characteristics and Evolution of PEDV in the Farm
3. Discussion
4. Materials and Methods
4.1. Farm Information
4.2. Sample Collection
4.3. Quantitative Real-Time RT-PCR (rRT-PCR)
4.4. Nucleotide Sequence Analysis
4.5. Virus Neutralization
4.6. Enzyme-Linked Immunosorbent Assay
4.7. Biosecurity Monitoring
4.8. Risk Assessment with the Pentagon Profile System
4.9. PED Vaccination Program
4.10. All-in-all-out (AIAO) Management
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, C. Porcine epidemic diarrhea virus: An emerging and re-emerging epizootic swine virus. J. Virol. 2015, 12, 193. [Google Scholar] [CrossRef] [Green Version]
- Lee, C. Porcine epidemic diarrhoea virus. In Porcine Viruses: From Pathogenesis to Strategies for Control; Zakaryan, H., Ed.; Caister Academic Press: Norfolk, UK, 2019; pp. 107–134. [Google Scholar]
- Gorbalenya, A.E.; Enjuanes, L.; Ziebuhr, J.; Snijder, E.J. Nidovirales: Evolving the largest RNA virus genome. Virus. Res. 2006, 117, 17–37. [Google Scholar] [CrossRef]
- Kocherhans, R.; Bridgen, A.; Ackermann, M.; Tobler, K. Completion of the porcine epidemic diarrhoea coronavirus (PEDV) genome sequence. Virus Genes 2001, 23, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Kweon, C.H.; Kwon, B.J.; Jung, T.S.; Kee, Y.J.; Hur, D.H.; Hwang, E.K.; An, S.H. Isolation of porcine epidemic diarrhea virus (PEDV) in Korea. Korean J. Vet. Res. 1993, 33, 249–254. [Google Scholar]
- Lee, S.; Ko, D.H.; Kwak, W.K.; Lim, C.H.; Moon, S.U.; Lee, D.S.; Lee, C. Reemergence of porcine epidemic diarrhea virus on Jeju Island. Korean. J. Vet. Res. 2014, 54, 185–188. [Google Scholar]
- Lee, S.; Lee, C. Outbreak-related porcine epidemic diarrhea virus strains similar to US strains, South Korea, 2013. Emerg. Infect. Dis. 2014, 20, 1223–1226. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.; Won, H.; Lee, D.U.; Noh, Y.H.; Lee, S.C.; Choi, H.W.; Yoon, I.J.; Lee, Y.J.; Yoo, H.S.; Lee, C. Assessment of the safety and efficacy of an attenuated live vaccine based on highly virulent genotype 2b porcine epidemic diarrhea virus in nursing piglets. Vet. Microbiol. 2019, 231, 120–128. [Google Scholar] [CrossRef]
- Lee, S.; Lee, C. Complete genome sequence of a novel S-insertion variant of porcine epidemic diarrhea virus from South Korea. Arch. Virol. 2017, 162, 2919–2922. [Google Scholar] [CrossRef]
- Lee, S.; Lee, C. Genomic and antigenic characterization of porcine epidemic diarrhoea virus strains isolated from South Korea, 2017. Transbound. Emerg. Dis. 2018, 65, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, D.U.; Noh, Y.H.; Lee, S.C.; Choi, H.W.; Yang, H.S.; Seol, J.H.; Mun, S.H.; Kang, W.M.; Yoo, H.; et al. Molecular characteristics and pathogenic assessment of porcine epidemic diarrhea virus isolates from the 2018 endemic outbreaks on Jeju Island, South Korea. Transbound. Emerg. Dis. 2019, 66, 1894–1909. [Google Scholar] [CrossRef] [PubMed]
- Park, C.K.; Lee, C. Clinical examination and control measures in a commercial pig farm persistently infected with porcine epidemic diarrhea (PED) virus. J. Vet. Clin. 2009, 26, 463–466. [Google Scholar]
- Lee, D.K.; Park, C.K.; Kim, S.H.; Lee, C. Heterogeneity in spike protein genes of porcine epidemic diarrhea viruses isolated in Korea. Virus Res. 2010, 149, 175–182. [Google Scholar] [CrossRef]
- Li, C.; Li, W.; Lucio de Esesarte, E.; Guo, H.; van den Elzen, P.; Aarts, E.; van den Born, E.; Rottier, P.J.M.; Bosch, B.J. Cell attachment domains of the porcine epidemic diarrhea virus spike protein are key targets of neutralizing antibodies. J. Virol. 2017, 12, e00273-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okda, F.A.; Lawson, S.; Singrey, A.; Nelson, J.; Hain, K.S.; Joshi, L.R.; Christopher-Hennings, J.; Nelson, E.A.; Diel, D.G. The S2 glycoprotein subunit of porcine epidemic diarrhea virus contains immunodominant neutralizing epitopes. Virology 2017, 509, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Chattha, K.S.; Roth, J.A.; Saif, L.J. Strategies for design and application of enteric viral vaccines. Annu. Rev. Anim. Biosci. 2015, 3, 375–395. [Google Scholar] [CrossRef]
- Langel, S.N.; Paim, F.C.; Lager, K.M.; Vlasova, A.N.; Saif, L.J. Lactogenic immunity and vaccines for porcine epidemic diarrhea virus (PEDV): Historical and current concepts. Virus Res. 2016, 226, 93–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjustrom-Kraft, J.; Woodard, K.; Giménez-Lirola, L.; Rotolo, M.; Wang, C.; Sun, Y.; Lasley, P.; Zhang, J.; Baum, D.; Gauger, P.; et al. Porcine epidemic diarrhea virus (PEDV) detection and antibody response in commercial growing pigs. BMC Vet. Res. 2016, 12, 99. [Google Scholar] [CrossRef] [Green Version]
- Crawford, K.; Lager, K.; Miller, L.; Opriessnig, T.; Gerber, P.; Hesse, R. Evaluation of porcine epidemic diarrhea virus transmission and the immune response in growing pigs. Vet. Res. 2015, 46, 49. [Google Scholar] [CrossRef] [Green Version]
- Environmental Stability of PEDV (Porcine Epidemic Diarrhea Virus). Available online: https://porkcheckoff.org/wp-content/uploads/2021/02/13-215-GOYAL-UMN.pdf (accessed on 17 April 2021).
- Tun, H.M.; Cai, Z.; Khafipour, E. Monitoring survivability and infectivity of porcine epidemic diarrhea virus (PEDv) in the infected on-farm earthen manure storages (EMS). Front. Microbiol. 2016, 7, 265. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Lin, C.M.; Annamalai, T.; Gao, X.; Lu, Z.; Esseili, M.A.; Jung, K.; El-Tholoth, M.; Saif, L.J.; Wang, Q. Determination of the infectious titer and virulence of an original US porcine epidemic diarrhea virus PC22A strain. Vet. Res. 2015, 46, 109. [Google Scholar] [CrossRef] [Green Version]
- Andersen, P.H.; Nielsen, M.; Lund, O. rediction of residues in discontinuous B-cell epitopes using protein 3D structures. Protein. Sci. 2006, 15, 2558–2567. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, I.J.; Pyo, H.M.; Tark, D.S.; Song, J.Y.; Hyun, B.H. Multiplex real-time RT-PCR for the simultaneous detection and quantification of transmissible gastroenteritis virus and porcine epidemic diarrhea virus. J. Virol. Methods. 2007, 146, 172–177. [Google Scholar] [CrossRef]
- Lee, S.; Son, K.Y.; Noh, Y.H.; Lee, S.C.; Choi, H.W.; Yoon, I.J.; Lee, C. Genetic characteristics, pathogenicity, and immunogenicity associated with cell adaptation of a virulent genotype 2b porcine epidemic diarrhea virus. Vet. Microbiol. 2017, 207, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Sagong, M.; Lee, C. Porcine reproductive and respiratory syndrome virus nucleocapsid protein modulates interferon-β production by inhibiting IRF3 activation in immortalized porcine alveolar macrophages. Arch. Virol. 2011, 156, 2187–2195. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, Y.; Lee, C. Isolation and characterization of a Korean porcine epidemic diarrhea virus strain KNU-141112. Virus Res. 2015, 208, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.N.; Lee, C. Complete genome sequence of a novel porcine parainfluenza virus 5 isolate in Korea. Arch. Virol. 2013, 158, 1765–1772. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The ClustalX windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.; Lee, K.W.; Choi, H.W.; Lee, C. Immunogenicity and protective efficacy of recombinant S1 domain of the porcine epidemic diarrhea virus spike protein. Arch. Virol. 2014, 159, 2977–2987. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.F.; Gong, Q.; Huang, Y.W.; Wang, C.; Holtkamp, D.; Opriessnig, T. Detection of antibodies against porcine epidemic diarrhea virus in serum and colostrum by indirect ELISA. Vet. J. 2014, 202, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.F.; Opriessnig, T. Detection of immunoglobulin (Ig) A antibodies against porcine epidemic diarrhea virus (PEDV) in fecal and serum samples. MethodsX 2015, 2, 368–373. [Google Scholar] [CrossRef]
- Jang, G.; Lee, S.; Lee, C. Assessing the risk of recurrence of porcine epidemic diarrhea virus in affected farms on Jeju Island, South Korea. J. Vet. Sci. 2021, 22, e48. [Google Scholar] [CrossRef]












 ) represents the date of virus detection.
) represents the date of virus detection.
) represents the date of virus detection.
) represents the date of virus detection.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Detection Date | Source | Strain Name | Sequence |
|---|---|---|---|
| 03-11-2019 | Feces | KNU-1907 | Full genome |
| 03-29-2019 | Feces | KNU-1907-1 | Spike |
| 05-02-2019 | Slurry | KNU-1907-2 | Spike |
| 06-18-2019 | Feces | KNU-1907-3 | Full genome |
| 06-26-2019 | Slurry | KNU-1907-4 | Spike |
| 07-01-2019 | Feces | KNU-1907-5 | Spike |
| 11-11-2019 | Slurry | KNU-1907-6 | Spike |
| 11-25-2019 | Slurry | KNU-1907-7 | Spike |
| 12-03-2019 | Feces | KNU-1907-8 | Full genome |
| 02-11-2020 | Slurry | KNU-1907-9 | Spike |
| 05-20-2020 | Slurry | KNU-1907-10 | Spike |
| 07-07-2020 | Slurry | KNU-1907-11 | Spike |
| Substitution | ||
|---|---|---|
| Mean (±S.E. 1) | 95% HPD 2 | |
| Spike (nt) | 1.683 × 10−4 (±0.02 10−4) | 2.082 × 10−5 (low) 3.216 × 10−4 (high) |
| Spike (aa) | 2.239 × 10−3 (±0.02 10−3) | 4.328 × 10−4 (low) 4.215 × 10−3 (high) |
| Full genome | 4.921 × 10−7 (±0.03 10−7) | 8.845 × 10−8 (low) 8.412 × 10−7 (high) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, G.; Park, J.; Lee, C. Successful Eradication of Porcine Epidemic Diarrhea in an Enzootically Infected Farm: A Two-Year Follow-Up Study. Pathogens 2021, 10, 830. https://doi.org/10.3390/pathogens10070830
Jang G, Park J, Lee C. Successful Eradication of Porcine Epidemic Diarrhea in an Enzootically Infected Farm: A Two-Year Follow-Up Study. Pathogens. 2021; 10(7):830. https://doi.org/10.3390/pathogens10070830
Chicago/Turabian StyleJang, Guehwan, Jonghyun Park, and Changhee Lee. 2021. "Successful Eradication of Porcine Epidemic Diarrhea in an Enzootically Infected Farm: A Two-Year Follow-Up Study" Pathogens 10, no. 7: 830. https://doi.org/10.3390/pathogens10070830
APA StyleJang, G., Park, J., & Lee, C. (2021). Successful Eradication of Porcine Epidemic Diarrhea in an Enzootically Infected Farm: A Two-Year Follow-Up Study. Pathogens, 10(7), 830. https://doi.org/10.3390/pathogens10070830