
The Carboxy Terminal Region on Spike Protein of Porcine Epidemic Diarrhea Virus (PEDV) Is Important for Evaluating Neutralizing Activity



Abstract
:1. Introduction
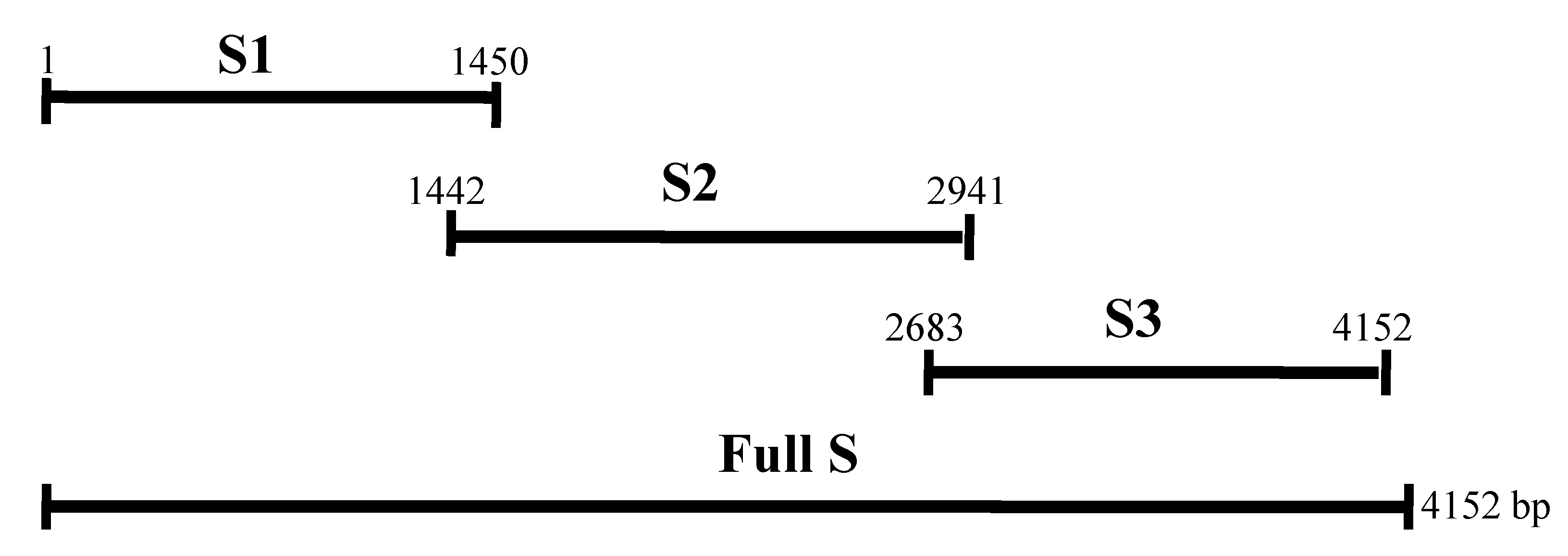
2. Results
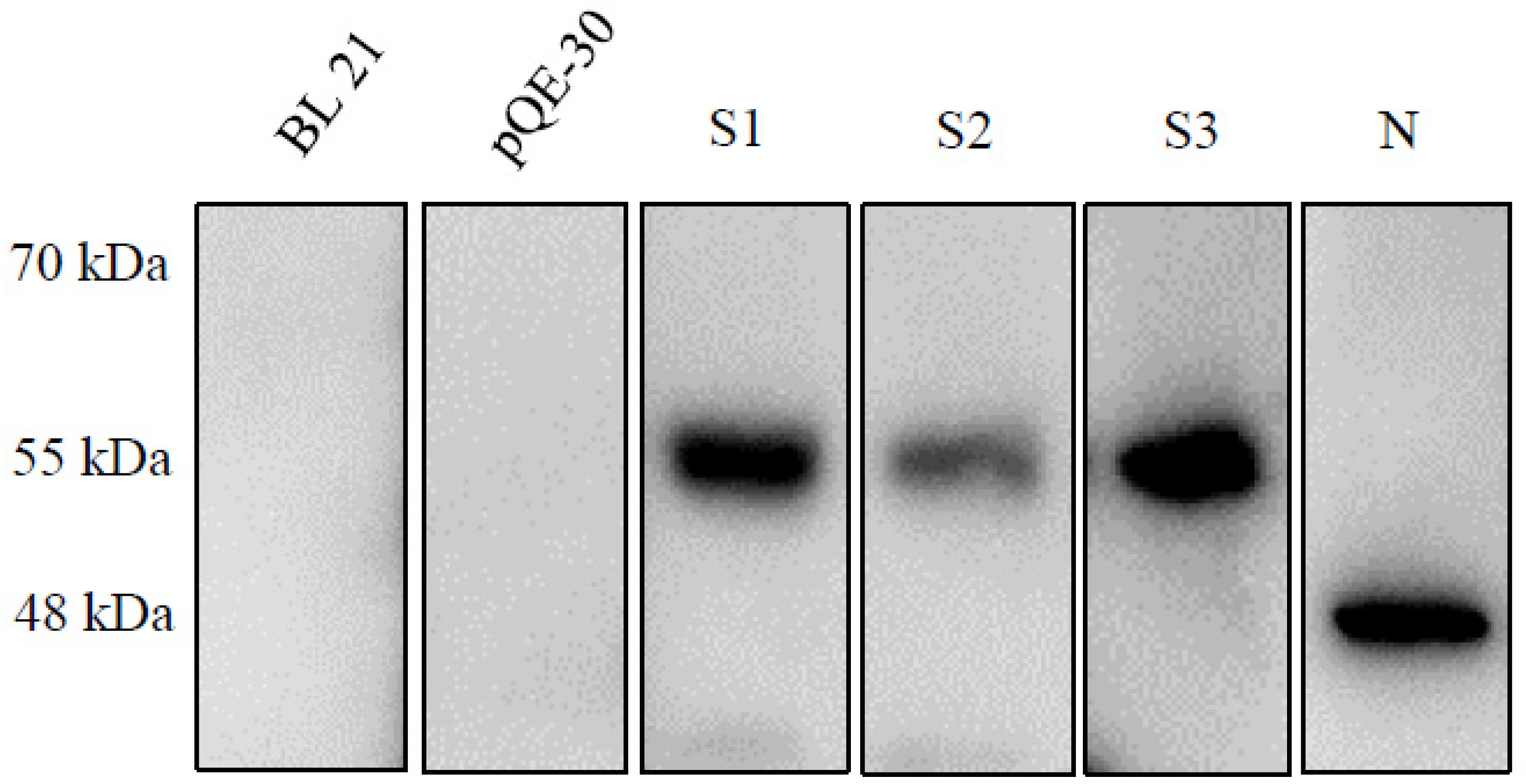
2.1. Expression of the Recombinant Spike and N Proteins
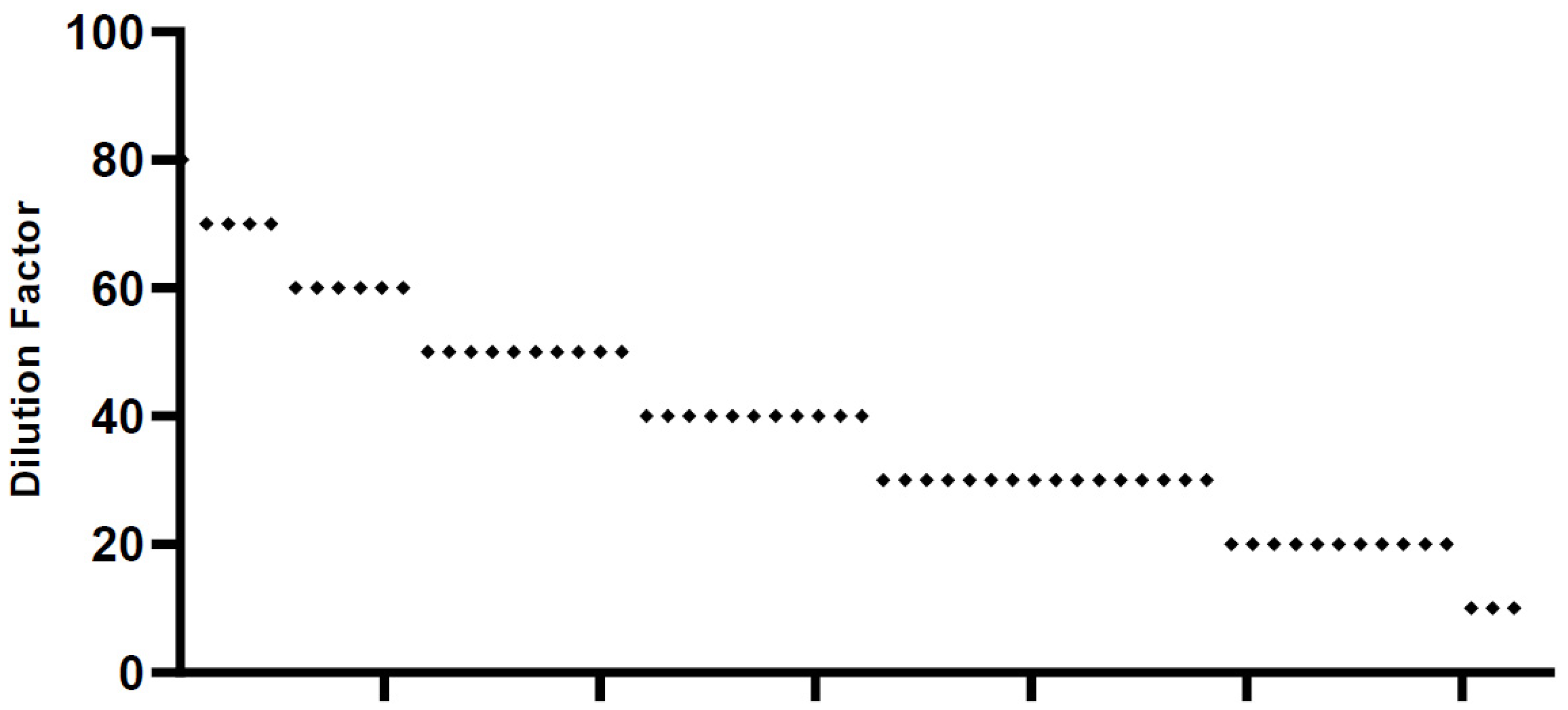
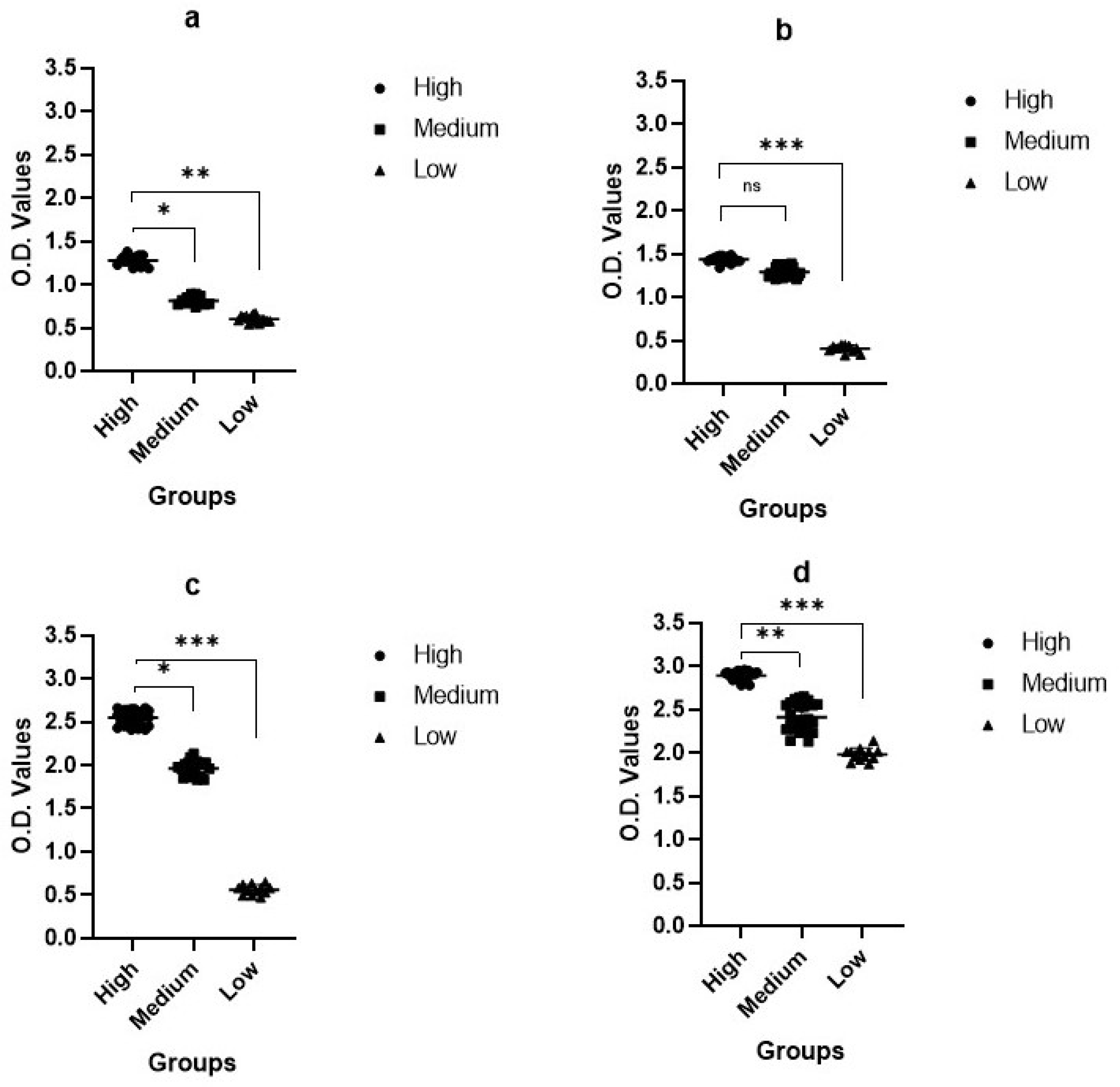
2.2. Serum Neutralization Test (SNT)
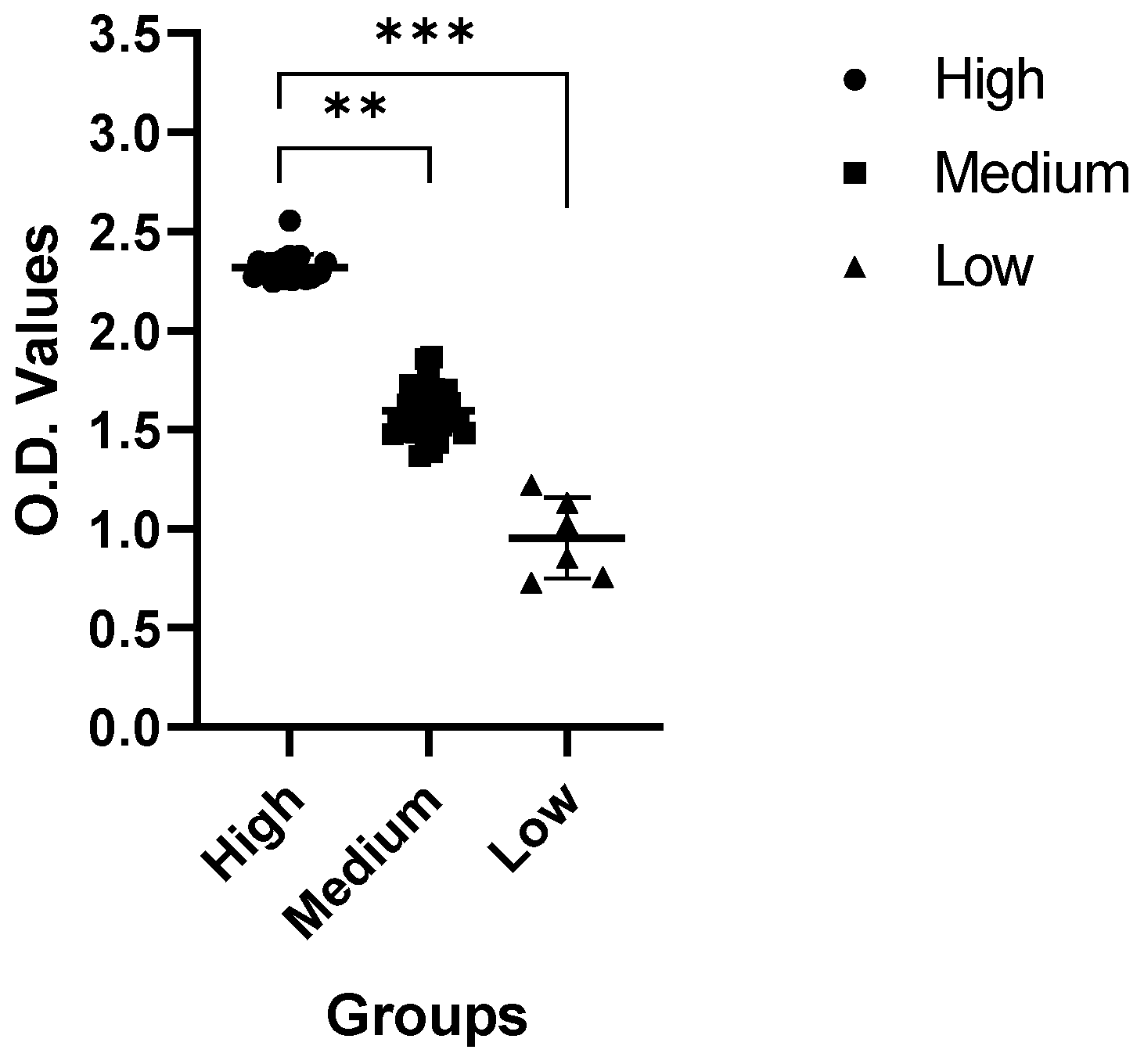
2.3. Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. Protein ELISA (pELISA)
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Viruses
4.3. RT-PCR for Amplification of Genes
4.4. Prokaryotic Expression
4.5. SDS-PAGE and Immunoblot Analysis
4.6. Serum Neutralization Test (SNT)
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Protein ELISA (pELISA)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Debouck, P.; Pensaert, M. Experimental infection of pigs with a new porcine enteric coronavirus, CV 777. Am. J. Vet. Res. 1980, 41, 219–223. [Google Scholar] [PubMed]
- Wang, Q.; Vkasiva, A.; Kenney, S.; Saif, L. Emerging and re-emerging coronaviruses in pigs. Curr. Opin. Microbiol. 2019, 34, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Okada, K.; Ohshima, K. An outbreak of swine diarrhea of a new-type associated with coronavirus-like particles in Japan. Nihon Juigaku Zasshi 1983, 45, 829–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C. Porcine epidemic diarrhea virus: An emerging and re-emerging epizootic swine virus. Virol. J. 2015, 12, 193. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.; Fang, L.; Jing, H.; Zeng, S.; Wang, D.; Liu, L.; Zhang, H.; Luo, R.; Chen, H.; Xiao, S. Porcine Epidemic Diarrhea Virus Nucleocapsid Protein Antagonizes Beta Interferon Production by Sequestering the Interaction between IRF3 and TBK1. J. Virol. 2014, 88, 8936–8945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.E.; Cruz, D.J.; Shin, H.J. Clathrin- and serine proteases-dependent uptake of porcine epidemic diarrhea virus into Vero cells. Virus Res. 2014, 191, 21–29. [Google Scholar] [CrossRef]
- Oh, J.S.; Lee, K.W.; Choi, H.W.; Lee, C.H. Immunogenicity and protective efficacy of recombinant S1 domain of the porcine epidemic diarrhea virus spike protein. Arch. Virol. 2014, 159, 2977–2987. [Google Scholar] [CrossRef]
- Li, C.H.; Li, W.T.; Esesarte, L.D.; Guo, H.B.; Elzen, P.V.D.; Aarts, E.; Born, E.V.D.; Rottier, P.J.M.; Bosch, B.J. Cell Attachment Domains of the Porcine Epidemic Diarrhea Virus Spike Protein Are Key Targets of Neutralizing Antibodies. J. Virol. 2017, 91, e00273-17. [Google Scholar] [CrossRef] [Green Version]
- Pijpers, A.; van Nieuwstadt, A.P.; Terpstra, C.; Verheijden, J.H. Porcine epidemic diarrhoea virus as a cause of persistent diarrhoea in a herd of breeding and finishing pigs. Vet. Rec. 1993, 132, 129–131. [Google Scholar] [CrossRef]
- Hofmann, M.; Wyler, R. Propagation of the virus of porcine epidemic diarrhea in cell culture. J. Clin. Microbiol. 1988, 26, 2235–2239. [Google Scholar] [CrossRef] [Green Version]
- Callebaut, P.; Debouck, P.; Pensaert, M. Enzyme-linked immunosorbent assay for the detection of the coronavirus-like agent and its antibodies in pigs with porcine epidemic diarrhea. Vet. Microbiol. 1982, 7, 295–306. [Google Scholar] [CrossRef]
- Carvajal, A.; Lanza, I.; Diego, R.; Rubio, P.; Carmenes, P. Evaluation of a blocking ELISA using monoclonal antibodies for the detection of porcine epidemic diarrhea virus and its antibodies. J. Vet. Diagn. Investig. 1995, 7, 60–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knuchel, M.; Ackermann, M.; Muller, H.K.; Kihm, U. An ELISA for detection of antibodies against porcine epidemic diarrhoea virus (PEDV) based on the specific solubility of the viral surface glycoprotein. Vet. Microbiol. 1992, 32, 117–134. [Google Scholar] [CrossRef]
- Kweon, C.H.; Lee, J.G.; Han, M.G.; Kang, Y.B. Rapid diagnosis of porcine epidemic diarrhea virus infection by polymerase chain reaction. J. Vet. Med. Sci. 1997, 59, 231–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Nieuwstadt, A.P.; Zetstra, T. Use of two enzyme-linked immunosorbent assays to monitor antibody responses in swine with experimentally induced infection with porcine epidemic diarrhea virus. Am. J. Vet. Res. 1991, 52, 1044–1050. [Google Scholar] [PubMed]
- Sueyoshi, M.; Tsuda, T.; Yamazaki, K.; Yoshida, K.; Nakazawa, M.; Sato, K.; Minami, T.; Iwashita, K.; Watanabe, M.; Suzuki, Y.; et al. An immunohistochemical investigation of porcine epidemic diarrhoea. J. Comp. Pathol. 1995, 113, 59–67. [Google Scholar] [CrossRef]
- Song, D.-S.; Kang, B.-K.; Lee, S.-S.; Yang, J.-S.; Moon, H.-J.; Oh, J.-S.; Ha, G.-W.; Jang, Y.-S.; Park, B.-K. An internal control in the quantitative RT-PCR assay for quantitation of porcine epidemic diarrhea virus shedding in pigs. J. Virol. Methods 2006, 133, 27–33. [Google Scholar] [CrossRef]
- To, T.L.; Ward, L.A.; Yuan, L.; Saif, L.J. Serum and intestinal isotype antibody responses and correlates of protective immunity to human rotavirus in a gnotobiotic pig model of disease. J. Gen. Virol. 1998, 79 Pt 11, 2661–2672. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Zhou, H.; Gao, L.; Li, B.; He, K.; Fa, H. Development and application of an indirect ELISA for the detection of antibodies to porcine epidemic diarrhea virus based on a recombinant spike protein. BMC Vet. Res. 2018, 14, 243. [Google Scholar] [CrossRef]
- Lirola, L.G.G.; Zhang, J.; Avil, J.A.C.; Chen, Q.; Magtoto, R.; Poonsuk, K.; Baum, D.H.; Piñeyro, P.; Zimmerman, J. Reactivity of Porcine Epidemic Diarrhea Virus Structural Proteins to Antibodies against Porcine Enteric Coronaviruses: Diagnostic Implications. J. Clin. Microbiol. 2017, 55, 1426–1436. [Google Scholar] [CrossRef] [Green Version]
- Egberink, H.F.; Ederveen, J.; Callebaut, P.; Horzinek, M.C. Characterization of the structural proteins of porcine epizootic diarrhea virus, strain CV777. Am. J. Vet. Res. 1988, 49, 1320–1324. [Google Scholar]
- Duarte, M.; Laude, H. Sequence of the spike protein of the porcine epidemic diarrhoea virus. J. Gen. Virol. 1994, 75 Pt 5, 1195–1200. [Google Scholar] [CrossRef]
- Bosch, B.J.; van der Zee, R.; de Haan, C.A.; Rottier, P.J. The coronavirus spike protein is a class I virus fusion protein: Structural and functional characterization of the fusion core complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.J.; Han, J.H.; Kwon, H.M. Partial sequence of the spike glycoprotein gene of transmissible gastroenteritis viruses isolated in Korea. Vet. Microbiol. 2003, 94, 195–206. [Google Scholar] [CrossRef]
- Cruz, D.J.; Kim, C.J.; Shin, H.J. Phage-displayed peptides having antigenic similarities with porcine epidemic diarrhea virus (PEDV) neutralizing epitopes. Virology 2006, 354, 28–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.H.; Bae, J.L.; Kang, T.J.; Kim, J.; Chung, G.H.; Lim, C.W.; Laude, H.; Yang, M.S.; Jang, Y.S. Identification of the epitope region capable of inducing neutralizing antibodies against the porcine epidemic diarrhea virus. Mol. Cells 2002, 14, 295–299. [Google Scholar] [PubMed]
- Kang, T.J.; Han, S.C.; Yang, M.S.; Jang, Y.S. Expression of synthetic neutralizing epitope of porcine epidemic diarrhea virus fused with synthetic B subunit of Escherichia coli heat-labile enterotoxin in tobacco plants. Protein Expr. Purif. 2006, 46, 16–22. [Google Scholar] [CrossRef]
- Godeke, G.J.; de Haan, C.A.; Rossen, J.W.; Vennema, H.; Rottier, P.J. Assembly of spikes into coronavirus particles is mediated by the carboxy-terminal domain of the spike protein. J. Virol. 2000, 74, 1566–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, R.; Montalto-Morrison, C.; Masters, P.S. Genetic analysis of determinants for spike glycoprotein assembly into murine coronavirus virions: Distinct roles for charge-rich and cysteine-rich regions of the endodomain. J. Virol. 2004, 78, 9904–9917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okda, F.A.; Lawson, S.; Singrey, A.; Nelson, J.; Hain, K.S.; Joshi, L.R.; Christopher-Hennings, J.; Nelson, E.A.; Diel, D.G. The S2 glycoprotein subunit of porcine epidemic diarrhea virus contains immunodominant neutralizing epitopes. Virology 2017, 509, 185–194. [Google Scholar] [CrossRef]
- Buratti, E.; Tisminetzky, S.G.; D’Agaro, P.; Baralle, F.E. A neutralizing monoclonal antibody previously mapped exclusively on human immunodeficiency virus type 1 gp41 recognizes an epitope in p17 sharing the core sequence IEEE. J. Virol. 1997, 71, 2457–2462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boucher, C.A.; Krone, W.J.; Goudsmit, J.; Meloen, R.H.; Naylor, P.H.; Goldstein, A.L.; Sun, D.K.; Sarin, P.S. Immune response and epitope mapping of a candidate HIV-1 p17 vaccine HGP30. J. Clin. Lab. Anal. 1990, 4, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Cruz, D.J.; Kim, C.J.; Shin, H. The GPRLQPY motif located at the carboxy-terminal of the spike protein induces antibodies that neutralize Porcine epidemic diarrhea virus. Virus Res. 2008, 132, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins (Gene Size) | Primer | Sequence |
|---|---|---|
| Spike S1 (1450 bp) | PED S1-S | 5′ GGA TCC ATG AGG TCT TTA ATT TAC TTC 3′ |
| PED S1-AS | 5′ GGT ACC ATG GGG TAAA AAC CAT CGT CAA GGT CAA AAG 3′ | |
| Spike S2 (1499 bp) | PED S2-S | 5′ GGA TCC TTA CCC CAT CTC TTC TAG AAA CCT 3′ |
| PED S2-AS | 5′ GGT ACC TAC AAA CAT ATG TAG CAC AAT CAA CAA CAC AC 3′ | |
| Spike S3 (1469 bp) | PED S3-S | 5′ GGA TCC AAG ACT TGC TTT TAA ATA 3′ |
| PED S3-AS | 5′ GGT ACC TTA GTG ATG GTG ATG GTG ATG CTG CAC GTG AAC CT3′ | |
| Nucleocapsid (1326 bp) | PED N-S | 5′ GGA TTC GCA TCT GTC AGC TTT CAG 3′ |
| PED N-AS | 5′ GGT ACC TTA ATT TCC TGT ATC GAA 3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, K.-J.; Kim, D.-H.; Hong, E.-J.; Shin, H.-J. The Carboxy Terminal Region on Spike Protein of Porcine Epidemic Diarrhea Virus (PEDV) Is Important for Evaluating Neutralizing Activity. Pathogens 2021, 10, 683. https://doi.org/10.3390/pathogens10060683
Kang K-J, Kim D-H, Hong E-J, Shin H-J. The Carboxy Terminal Region on Spike Protein of Porcine Epidemic Diarrhea Virus (PEDV) Is Important for Evaluating Neutralizing Activity. Pathogens. 2021; 10(6):683. https://doi.org/10.3390/pathogens10060683
Chicago/Turabian StyleKang, Ki-Jong, Dong-Hwan Kim, Eui-Ju Hong, and Hyun-Jin Shin. 2021. "The Carboxy Terminal Region on Spike Protein of Porcine Epidemic Diarrhea Virus (PEDV) Is Important for Evaluating Neutralizing Activity" Pathogens 10, no. 6: 683. https://doi.org/10.3390/pathogens10060683
APA StyleKang, K. -J., Kim, D. -H., Hong, E. -J., & Shin, H. -J. (2021). The Carboxy Terminal Region on Spike Protein of Porcine Epidemic Diarrhea Virus (PEDV) Is Important for Evaluating Neutralizing Activity. Pathogens, 10(6), 683. https://doi.org/10.3390/pathogens10060683