
Antibiogram Profile of Acinetobacterbaumannii Recovered from Selected Freshwater Resources in the Eastern Cape Province, South Africa



Abstract
:1. Introduction
2. Results
2.1. Identification and Antibiotic Resistance Profile of Acinetobacter Species
2.2. Phenotypic Antibiotic Resistance (PAR) Profile of A. baumannii
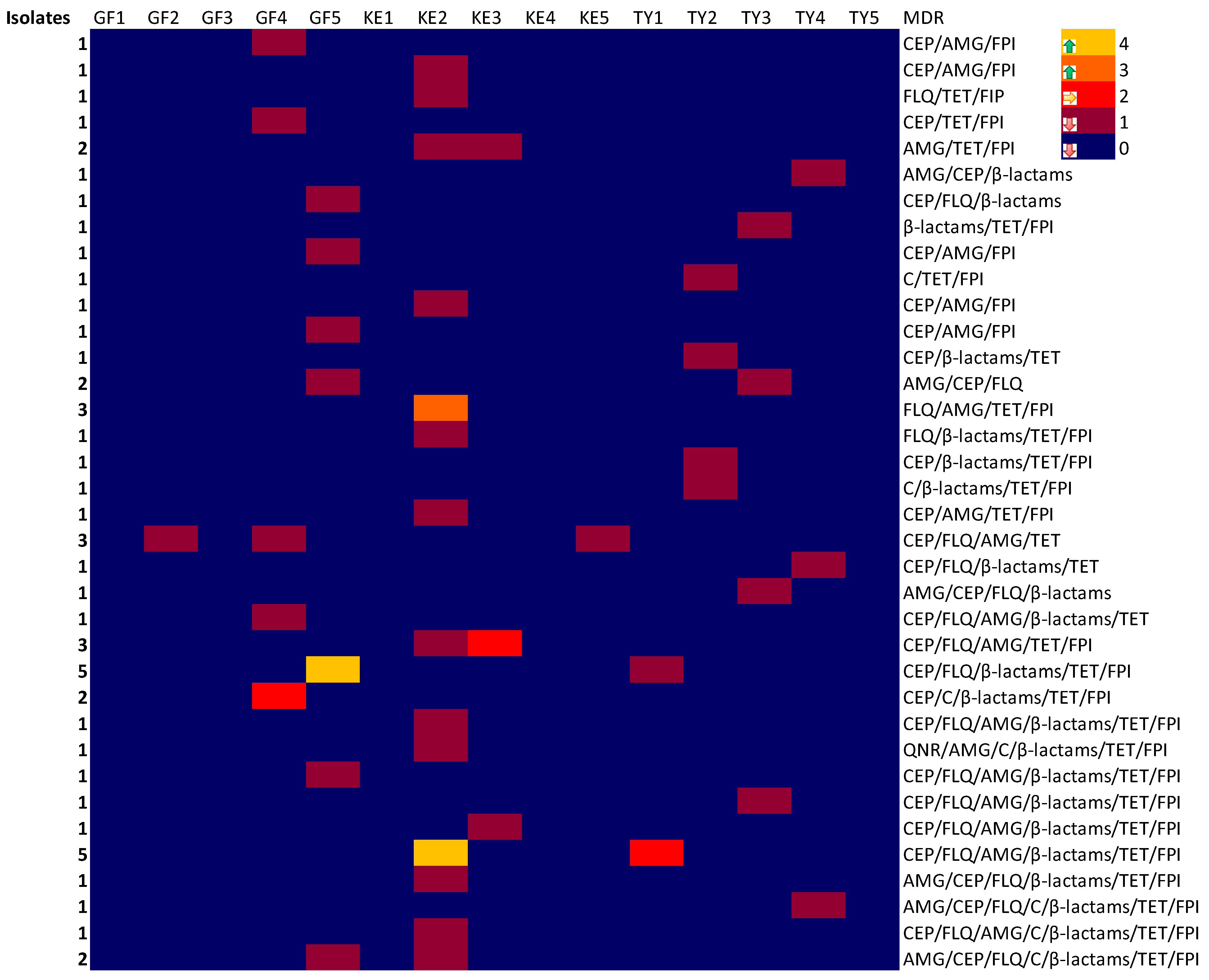
2.3. Multiple Drug Resistance (MDR) Phenotypes
2.4. Profile of ARGs among A. baumannii
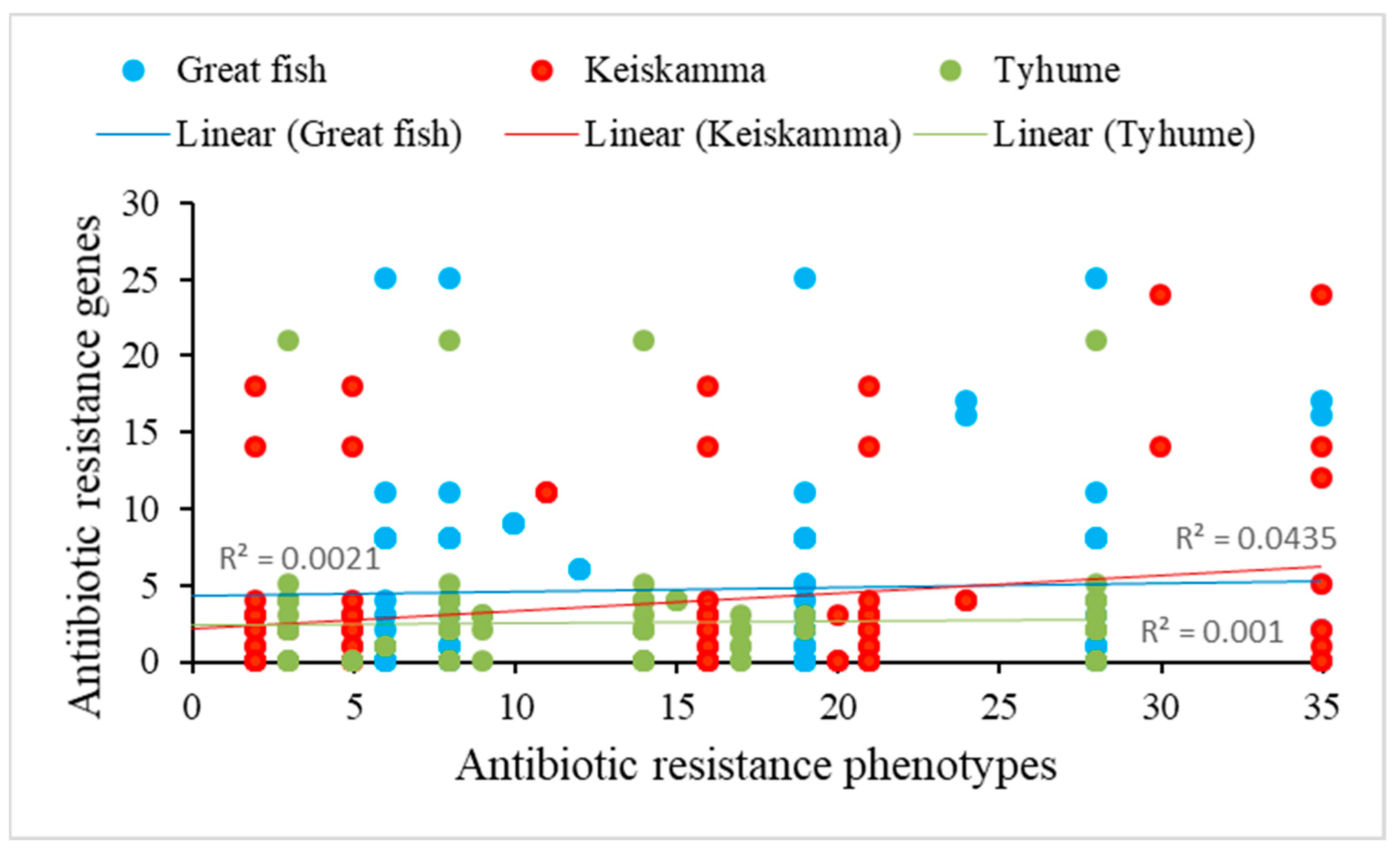
2.5. Correlation between PAR and ARGs
3. Discussion
4. Materials and Methods
4.1. Description of Sampling Areas
4.2. Isolation and Molecular Confirmation of A. baumannii
4.3. Antibiotics Susceptibility Testing (AST)
4.4. Screening of A. baumannii for Antibiotic Resistance Genes (ARGs)
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maravić, A.; Skočibušić, M.; Fredotović, Z.; Šamanić, I.; Cvjetan, S.; Knezović, M.; Puizina, J. Urban riverine environment is a source of multidrug-resistant and ESBL-producing clinically important Acinetobacter spp. Environ. Sci. Pollut. Res. 2016, 23, 3525–3535. [Google Scholar] [CrossRef]
- Doughari, H.J.; Ndakidemi, P.A.; Human, I.S.; Benade, S. The ecology, biology and pathogenesis of Acinetobacter spp.: An overview. Microbes Environ. 2011, 26, 101–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basri, R.; Zueter, A.R.; Mohamed, Z.; Alam, M.K.; Norsa’adah, B.; Hasan, S.A.; Hasan, H.; Ahmad, F. Burden of bacterial meningitis: A retrospective review on laboratory parameters and factors associated with death in meningitis, Kelantan Malaysia. Nagoya J. Med. Sci. 2015, 77, 59. [Google Scholar] [PubMed]
- Chiang, T.; Pastagia, M.; Huang, D.B. Bacteremia Caused by Acinetobacter baumannii: Epidemiologic Features, Antimicrobial Susceptibility, and Outcomes. Adv. Infect. Dis. 2014, 4, 66–71. [Google Scholar] [CrossRef] [Green Version]
- Gundeslioglu, O.O.; Gokmen, T.G.; Horoz, O.O.; Aksaray, N.; Koksal, F.; Yaman, A.; Yıldızdas, R.D.; Alhan, E.; Kocabas, E.; Alabaz, D. Molecular epidemiology and antibiotic susceptibility pattern of Acinetobacter baumannii isolated from children in a Turkish University hospital. Turk. J. Pediatrics 2014, 56, 360–367. [Google Scholar]
- Guerrero, D.M.; Perez, F.; Conger, N.G.; Solomkin, J.S.; Adams, M.D.; Rather, P.N.; Bonomo, R.A. Acinetobacter baumannii-Associated Skin and Soft Tissue Infections: Recognizing a Broadening Spectrum of Disease. Surg. Infect. 2010, 11, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Darvishi, M. Virulence factors profile and antimicrobial resistance of Acinetobacter baumannii strains isolated from various infections recovered from immunosuppressive patients. Biomed. Pharm. J. 2016, 9, 1057–1062. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDPC). Annual Epidemiological Report Food-and Waterborne Diseases and Zoonoses; European Centre for Disease Prevention and Control (ECDPC): Stockholm, Sweden, 2014. [Google Scholar]
- World Health Organization (WHO). Global Report on Surveillance of Antimicrobial Resistance; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- Kanafani, A.Z.; Kanj, S.S. Ministry of Health, Kingdome of Saudi Arabia. 2014. Available online: http://www.uptodate.com/contents/acinetobacterinfection-treatment-and-prevention (accessed on 6 April 2020).
- Gallagher, L.A.; Ramage, E.; Weiss, E.J.; Radey, M.; Hayden, H.S.; Held, K.G.; Huse, H.K.; Zurawski, D.V.; Brittnacher, M.J.; Manoil, C. Resources for genetic and genomic analysis of emerging pathogen Acinetobacter baumannii. J. Bacteriol. 2015, 197, 2027–2035. [Google Scholar] [CrossRef] [Green Version]
- Abdi, S.N.; Ghotaslou, R.; Ganbarov, K.; Mobed, A.; Tanomand, A.; Yousefi, M.; Asgharzadeh, M.; Kafil, H.S. Acinetobacter baumannii Efflux Pumps and Antibiotic Resistance Infection and Drug Resistance. Infect. Drug Resist. 2020, 13, 423–434. [Google Scholar] [CrossRef] [Green Version]
- Rumbo, C.; Gato, E.; López, M.; de Alegría, C.R.; Fernández-Cuenca, F.; Martínez-Martínez, L.; Vila, J.; Pachón, J.; Cisneros, J.M.; Rodríguez-Baño, J.; et al. Contribution of efflux pumps, porins, and β-lactamases to multidrug resistance in clinical isolates of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2013, 57, 5247–5257. [Google Scholar] [CrossRef] [Green Version]
- Gheorghe, I.; Novais, A.; Grosso, F.; Rodrigues, C.; Chifiriuc, M.C.; Lazar, V.; Peixe, L. Snapshot on carbapenemase-producing Pseudomonas aeruginosa and Acinetobacter baumannii in Bucharest hospitals reveals unusual clones and novel genetic surroundings for blaOXA-23. J. Antimicrob. Chemother. 2015, 70, 1016–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez-López, R.; Solano-Gálvez, S.G.; Vignon-Whaley, J.J.J.; Vaamonde, J.A.A.; Alonzo, L.A.P.; Reséndiz, A.R.; Álvarez, M.M.; López, E.N.V.; Franyuti-Kelly, G.; Álvarez-Hernández, D.A.; et al. Acinetobacter baumannii Resistance: A Real Challenge for Clinicians. Antibiotics 2020, 9, 205. [Google Scholar] [CrossRef] [PubMed]
- Ardehal, S.H.; Azimi, T.; Fallah, F.; Owrang, M.; Aghamohammadi, N.; Azimi, L. Role of efflux pumps in reduced susceptibility to tigecycline in Acinetobacter baumannii. New Microbes New Infect. 2019, 30, 1–6. [Google Scholar] [CrossRef]
- Perez, F.; Hujer, A.M.; Hujer, K.M.; Decker, B.K.; Rather, P.N.; Bonomo, R.A. Global challenge of multidrug-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2007, 51, 3471–3484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falagas, M.E.; Vardakas, K.Z.; Kapaskelis, A.; Triarides, N.A.; Roussos, N.S. Tetracyclines for multidrug-resistant Acinetobacter baumannii infections. Int. J. Antimicrob. Agents 2015, 45, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Hrenovic, J.; Durn, G.; Goic-Barisic, I.; Kovacic, A. Occurrence of an Environmental Acinetobacter baumannii Strain Similar to a Clinical Isolate in Paleosol from Croatia. Appl. Environ. Microbiol. 2014, 80, 2860–2866. [Google Scholar] [CrossRef] [Green Version]
- Goswami, R.; Mukherjee, S.; Rana, V.S.; Saha, D.R.; Raman, R.; Padhy, P.K.; Mazumder, S. Isolation and Characterization of Arsenic-Resistant Bacteria from Contaminated Water-Bodies in West Bengal, India. Geomicrobiol. J. 2015, 32, 17–26. [Google Scholar] [CrossRef]
- Kittinger, C.; Kirschner, A.; Lipp, M.; Baumert, R.; Mascher, F.; Farnleitner, A.H.; Zarfel, G.E. Antibiotic Resistance of Acinetobacter spp. isolates from the River Danube: Susceptibility Stays High. Int. J. Environ. Res. Public Health 2018, 15, 52. [Google Scholar] [CrossRef] [Green Version]
- Ashbolt, N.J. Microbial contamination of drinking water and human health from community water systems. Curr. Environ. Health Rpt. 2015, 2, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Bitton, G. Microbiology of Drinking Water Production and Distribution, 1st ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; p. 312. [Google Scholar]
- Fernando, D.M.; Tun, H.M.; Poole, J.; Patidar, R.; Li, R.; Mi, R.; Amarawansha, G.E.A.; Fernando, W.G.D.; Khafipour, E.; Farenhorst, A.; et al. Detection of antibiotic resistance genes in source and drinking water samples from a First Nations community in Canada. Appl. Environ. Microbiol. 2016, 82, 4767–4775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.C.; Chou, M.Y.; Shih, Y.J.; Huang, T.Y.; Yang, P.Y.; Chiu, Y.C.; Chen, J.S.; Hsu, B.M. Distribution and genotyping of aquatic Acinetobacter baumannii strains isolated from the Puzi River and its tributaries near areas of livestock farming. Water 2018, 10, 1374. [Google Scholar] [CrossRef] [Green Version]
- Cikman, A.; Gulhan, B.; Aydin, M.; Ceylan, M.R.; Parlak, M.; Karakecili, F.; Karagoz, A. In vitro Activity of Colistin in Combination with Tigecycline against Carbapenem-Resistant Acinetobacter baumannii Strains Isolated from Patients with Ventilator-Associated Pneumonia. Int. J. Med. Sci. 2015, 12, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Anh, N.T.; Thieu Nga, T.V.; Tuan, H.M.; Tuan, N.S.; Dao, Y.; Chau, N.V.V.; Baker, S.; Duong, H.H.T. Molecular epidemiology and antimicrobial resistance phenotypes of Acinetobacter baumannii isolated from patients in three hospitals in southern Vietnam. J. Med. Microbiol. 2017, 66, 46–53. [Google Scholar]
- Ekwealor, P.A.; Ugwu, M.C.; Ezeobi, I.; Amalukwe, G.; Ugwu, B.C.; Okezie, U.; Stanley, C.; Esimone, C. Antimicrobial Evaluation of Bacterial Isolates from Urine Specimen of Patients with Complaints of Urinary Tract Infections in Awka, Nigeria. Int. J. Microbiol. 2016, 2016, 9740273. [Google Scholar] [CrossRef] [Green Version]
- Al-Bahry, S.; Mahmoud, I.; Elshafie, A.; Al-Harthy, A.; Al-Ghafri, S.; Al-Amri, I.; Alkindi, A. Bacterial flora and antibiotic resistance from eggs of green turtles Chelonia myans: An indication of polluted effluents. Mar. Pollut. Bull. 2009, 58, 720–725. [Google Scholar] [CrossRef]
- Leungtongkam, U.; Thummeepak, R.; Tasanapak, K.; Sitthisak, S. Acquisition and transfer of antibiotic resistance genes in association with conjugative plasmid or class 1 integrons of Acinetobacter baumannii. PLoS ONE 2018, 13, e0208468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taitt, C.R.; Leski, T.A.; Stockelman, M.G.; Craft, D.W.; Zurawski, D.V.; Kirkup, B.C.; Vora, G.J. Antimicrobial resistance determinants in Acinetobacter baumannii isolates taken from Military Treatment Facilities. Antimicrob. Agents Chemother. 2013, 58, 767–781. [Google Scholar] [CrossRef] [Green Version]
- Hujer, K.M.; Hujer, A.M.; Hulten, E.A.; Bajaksouzian, S.; Adams, J.M.; Donskey, C.J.; Ecker, D.J.; Massire, C.; Eshoo, M.W.; Sampath, R.; et al. Analysis of antibiotic resistance genes in multidrug-resistant Acinetobacter sp. isolates from military and civilian patients treated at the Walter Reed Army Medical Center. Antimicrob. Agents Chemother. 2006, 50, 4114–4123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.F.; Lan, C.Y. Antimicrobial resistance in Acinetobacter baumannii: From bench to bedside. World J. Clin. Cases 2014, 2, 787–814. [Google Scholar] [CrossRef] [PubMed]
- Potron, A.; Poirel, L.; Nordmann, P. Emerging broad-spectrum resistance in Pseudomonas aeruginosa and Acinetobacter baumannii: Mechanisms and epidemiology. Int. J. Antimicrob. Agents 2015, 45, 568–585. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Corvec, S.; Rapoport, M.; Mugnier, P.; Petroni, A.; Pasteran, F.; Faccone, D.; Galas, M.; Drugeon, H.; Cattoir, V.; et al. Identification of the novel narrow-spectrum beta-lactamase SCO1 in Acinetobacter spp. from Argentina. Antimicrob. Agents Chemother. 2007, 51, 2179–2184. [Google Scholar] [CrossRef] [Green Version]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [Green Version]
- Hamidian, M.; Ambrose, S.J.; Hall, R.M. A large conjugative Acinetobacter baumannii plasmid carrying the sul2 sulphonamide and strAB streptomycin resistance genes. Plasmid 2016, 87, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Jamal, S.; Al Atrouni, A.; Rafei, R.; Dabboussi, F.; Hamze, M.; Osman, M. Molecular mechanisms of antimicrobial resistance in Acinetobacter baumannii, with a special focus on its epidemiology in Lebanon. J. Glob. Antimicrob. Resist. 2018, 15, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Knight, D.B.; Rudin, S.D.; Bonomo, R.A.; Rather, P.N. Acinetobacter nosocomialis: Defining the Role of Efflux Pumps in Resistance to Antimicrobial Therapy, Surface Motility, and Biofilm Formation. Front. Microbiol. 2018, 9, 1902. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.C. Tetracycline resistance determinants: Mechanisms of action, regulation of expression, genetic mobility, and distribution. FEMS Microbiol. Rev. 1996, 19, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Guardabassi, L.; Dijkshoorn, L.; Collard, J.M.; Olsen, J.E.; Dalsgaard, A. Distribution and in-vitro transfer of tetracycline resistance determinants in clinical and aquatic Acinetobacter strains. J. Med. Microbiol. 2000, 49, 929–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribera, A.; Ruiz, J.; Vila, J. Presence of the tetM determinant in a clinical isolate of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2003, 47, 2310–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattoir, V. Quinolones de l’antibiogramme aux phénotypes de résistance Quinolones: From antibiogram to resistance phenotypes. Revue Franco-Phone Lab. 2012, 445, 79–87. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L. Emergence of plasmid-mediated resistance to quinolones in Enterobacteriaceae. J. Antimicrob. Chemother. 2005, 56, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Bonnin, R.A.; Nordmann, P. Genetic basis of antibiotic resistance in pathogenic Acinetobacter species. IUBMB Life 2011, 63, 1061–1067. [Google Scholar] [CrossRef] [PubMed]
- Towner, K.J. Antibiotic resistance in Acinetobacter spp. In Acinetobacter Molecular Biology; Gerischer, U., Ed.; Caister Academic Press: Norfolk, UK, 2008; pp. 331–343. [Google Scholar]
- Nemec, A.; Dolzani, L.; Brisse, S.; van den Broek, P.; Dijkshoorn, L. Diversity of aminoglycoside-resistance genes and their association with class 1 integrons among strains of pan-European Acinetobacter baumannii clones. J. Med. Microbiol. 2004, 53, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- American Public Health Association (APHA)/American Water Works Association (AWWA)/Water Environment Federation (WEF). Standard Methods for the Examination of Water and Wastewater; APHA/AWWA/WEF: Washington, DC, USA, 2005. [Google Scholar]
- Chen, T.L.; Lee, Y.T.; Kuo, S.C.; Yang, S.P.; Fung, C.P.; Lee, S.D. Rapid identification of Acinetobacter baumannii, Acinetobacter nosocomialis and Acinetobacter pittii with a multiplex PCR assay. J. Med. Microbiol. 2014, 63, 1154–1159. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Disk Susceptibility Tests, 28th ed.; CLSI Supplement M100: Wayne, PA, USA, 2018. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Agent | Percentage of Isolates Sensitive to Antibiotics | |||
|---|---|---|---|---|
| Susceptible (S) | Resistance (R) | Intermediate Resistance (I) | Non-Susceptible | |
| R + I | ||||
| PTZ | 299 (72.9%) | 46 (11.2%) | 65 (15.8%) | 111 (27%) |
| CAZ | 289 (70.5%) | 49 (12%) | 72 (17.6%) | 121 (29.6%) |
| CTX | 69 (16.8%) | 77 (18.8%) | 264 (64.4%) | 341 (83.2%) |
| CPM | 363 (88.5%) | 36 (8.8%) | 11 (2.7%) | 47 (11.5%) |
| IMI | 394 (96.1%) | 11 (2.7%) | 5 (1.2%) | 16 (3.9%) |
| MEM | 381 (92.9%) | 17 (4.15%) | 12 (2.9%) | 29 (7.05%) |
| AK | 400 (97.6%) | 10 (2.4%) | 0 (0%) | 10 (2.4%) |
| GM | 369 (90.0%) | 36 (8.8%) | 5 (1.2%) | 41 (10%) |
| TET | 307 (74.9%) | 69 (16.8%) | 34 (8.3%) | 103 (25.1%) |
| CIP | 312 (76.1%) | 45 (11%) | 53 (12.9%) | 98 (23.9%) |
| SXT | 304 (74.1%) | 84 (20.5%) | 22 (5.4%) | 106 (25.9%) |
| Antibiotic Class | Antibiotic Agent | Resistance Genes | No. of Positive Isolates Per Sampling Site | Total (%) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GF1 | GF2 | GF3 | GF4 | GF5 | KE1 | KE2 | KE3 | KE4 | KE5 | TY1 | TY2 | TY3 | TY4 | TY5 | ||||
| apHAI | 0 | 0 | 0 | 5 | 4 | 0 | 0 | 3 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 13 (25.5) | ||
| apHA2 | 0 | 0 | 0 | 3 | 3 | 0 | 11 | 2 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 21 (41.2) | ||
| β-lactamases (n = 101) | PTZ CAZ CTX CPM IMI MEM | blaTEM | 1 | 3 | 0 | 9 | 14 | 0 | 12 | 5 | 0 | 0 | 2 | 1 | 3 | 7 | 8 | 65 (64.4) |
| blaSHV | 0 | 0 | 0 | 5 | 3 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 12 (11.9) | ||
| blaCTX-M-1 | 0 | 0 | 0 | 3 | 5 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 12 (11.9) | ||
| blaCTX-M-2 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 2 (2.0) | ||
| blaCTX-M-9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| blaCTX-M-8/-25 | 1 | 0 | 1 | 0 | 6 | 0 | 2 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 2 | 12 (11.9) | ||
| blaPER | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 (2.9) | ||
| blaVEB | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 2 | 5 (4.9) | ||
| blaGES | 0 | 2 | 0 | 1 | 5 | 0 | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 13 (12.9) | ||
| blaOXA-23 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| blaOXA-40 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| blaOXA-48 | 0 | 0 | 1 | 2 | 0 | 0 | 3 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 9 (8.9) | ||
| blaOXA-51 | 0 | 0 | 2 | 3 | 6 | 0 | 11 | 3 | 0 | 0 | 2 | 1 | 0 | 0 | 1 | 29 (28.7) | ||
| blaOXA-58 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| blaVIM | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| blaIMP | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 6 (5.9) | ||
| blaKPC | 0 | 0 | 0 | 1 | 1 | 0 | 2 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 8 (7.9) | ||
| Fluoroquilonone (n = 44) | CIP | qnrA | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| qnrB | 0 | 0 | 0 | 0 | 2 | 0 | 1 | 2 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 7 (14.6) | ||
| qnrC | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| qnrD | 0 | 0 | 0 | 1 | 4 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 8 (16.7) | ||
| qnrS | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| Sulfonamides (n = 83) | SXT | sul1 | 1 | 1 | 1 | 6 | 7 | 2 | 8 | 3 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 32 (37.1) |
| sul2 | 0 | 1 | 0 | 7 | 9 | 0 | 20 | 3 | 0 | 0 | 1 | 0 | 0 | 0 | 2 | 43 (49.4) | ||
| Tetracyclines (n = 70) | TET | tet(A) | 0 | 0 | 0 | 1 | 1 | 0 | 2 | 3 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 8 (11.4) |
| tet(B) | 0 | 0 | 0 | 4 | 1 | 0 | 12 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 19 (27.1) | ||
| tet(C) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 2 (2.9) | ||
| tet(L) | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 3 (4.3) | ||
| tet(M) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 (2.9) | ||
| tet(O) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| Antimicrobial Group | Antimicrobial Agent | Disc Content |
|---|---|---|
| β-lactam/β-lactamase-inhibitor combinations | Piperacillin-tazobactam (PTZ) | 100/10 μg |
| Cephems (CEP) | Ceftazidime (CAZ) | 30 μg |
| Cefepime (CPM) | 30 μg | |
| Cefotaxime (CTX) | 30 μg | |
| Carbapenems (C) | Imipenem (IMI) | 10 μg |
| Meropenem (MEM) | 10 μg | |
| Aminoglycosides (AMG) | Amikacin (AK) | 30 μg |
| Gentamicin (GM) | 10 μg | |
| Tetracyclines (TET) | Tetracycline (TET) | 30 μg |
| Fluoroquinolones (FLQ) | Ciprofloxacin (CIP) | 5 μg |
| Folate pathway inhibitors (FPI) | Trimethoprim/sulfamethoxazole (SXT) | 1.25/23.75 μg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adewoyin, M.A.; Ebomah, K.E.; Okoh, A.I. Antibiogram Profile of Acinetobacterbaumannii Recovered from Selected Freshwater Resources in the Eastern Cape Province, South Africa. Pathogens 2021, 10, 1110. https://doi.org/10.3390/pathogens10091110
Adewoyin MA, Ebomah KE, Okoh AI. Antibiogram Profile of Acinetobacterbaumannii Recovered from Selected Freshwater Resources in the Eastern Cape Province, South Africa. Pathogens. 2021; 10(9):1110. https://doi.org/10.3390/pathogens10091110
Chicago/Turabian StyleAdewoyin, Mary Ayobami, Kingsley Ehi Ebomah, and Anthony Ifeanyi Okoh. 2021. "Antibiogram Profile of Acinetobacterbaumannii Recovered from Selected Freshwater Resources in the Eastern Cape Province, South Africa" Pathogens 10, no. 9: 1110. https://doi.org/10.3390/pathogens10091110
APA StyleAdewoyin, M. A., Ebomah, K. E., & Okoh, A. I. (2021). Antibiogram Profile of Acinetobacterbaumannii Recovered from Selected Freshwater Resources in the Eastern Cape Province, South Africa. Pathogens, 10(9), 1110. https://doi.org/10.3390/pathogens10091110