
Plant- and Bacteria-Derived Compounds with Anti-Philasterides dicentrarchi Activity

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Analysis of the Toxicity to P. dicentrarchi of Natural Compounds
2.2. Effect of Pseudomonas H6 Surfactant on P. dicentrarchi
3. Discussion
4. Materials and Methods
4.1. Ciliate Culture
4.2. Culture of the Epithelioma Papulosum Cyprini (EPC) Cell Line
4.3. Determination of Anticiliate Activity
4.4. Assessment of the Toxicity of Compounds on EPC Cell Line
4.5. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iglesias, R.; Paramá, A.; Álvarez, M.F.; Leiro, J.; Fernández, J.; Sanmartín, M.L. Philasterides dicentrarchi (Ciliophora, Scuticociliatida) as the Causative Agent of Scuticociliatosis in Farmed Turbot, Scophthalmus maximus in Galicia (NW Spain). Dis. Aquat. Organ. 2001, 46, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez-Pellitero, P.; Palenzuela, O.; Padrós, F.; Sitjà-Bobadilla, A.; Riaza, A.; Silva, R.; Arán, J. Histiophagous Scuticocliatids (Ciliophora) Parasitizing Turbot Scophthalmus maximus: Morphology, in vitro Culture and Virulence. Folia Parasitol. 2004, 51, 177–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, M.F.; Costa, A.R.; Barandela, T.; Saraiva, A.; Rodrigues, P.N. Scuticociliate Infection and Pathology in Cultured Turbot Scophthalmus maximus from the North of Portugal. Dis. Aquat. Organ. 2007, 74, 249–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyková, I.; Figueras, A. Histopathological Changes in Turbot Scophthalmus maximus due to a Histiophagous Ciliate. Dis. Aquat. Organ. 1994, 18, 5–9. [Google Scholar] [CrossRef]
- Sterud, E.; Hansen, M.K.; Mo, T.A. Systemic Infection with Uronema-like Ciliates in Farmed Turbot, Scophthalmus maximus (L.). J. Fish Dis. 2000, 23, 33–37. [Google Scholar] [CrossRef]
- Paramá, A.; Iglesias, R.; Álvarez, M.F.; Leiro, J.; Aja, C.; Sanmartín, M.L. Philasterides dicentrarchi (Ciliophora, Scuticociliatida): Experimental Infection and Possible Routes of Entry in Farmed Turbot (Scophthalmus maximus). Aquaculture 2003, 217, 73–80. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Palenzuela, O.; Álvarez-Pellitero, P. Immune Response of Turbot, Psetta Maxima (L.) (Pisces: Teleostei), to Formalin-Killed Scuticociliates (Ciliophora) and Adjuvanted Formulations. Fish Shellfish Immunol. 2008, 24, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sanmartín, M.L.; Paramá, A.; Castro, R.; Cabaleiro, S.; Leiro, J.; Lamas, J. Vaccination of Turbot against the Protozoan Parasite Philasterides dicentrarchi. Effects on Antibody Production and Protection. J. Fish Dis. 2008, 31, 135–140. [Google Scholar] [CrossRef]
- Lamas, J.; Sanmartín, M.L.; Paramá, A.I.; Castro, R.; Cabaleiro, S.; Ruiz de Ocenda, M.V.; Barja, J.L.; Leiro, J. Optimization of an Inactivated Vaccine against a Scuticociliate Parasite of Turbot: Effect of Antigen, Formalin and Adjuvant Concentration on Antibody Response and Protection against the Pathogen. Aquaculture 2008, 278, 22–26. [Google Scholar] [CrossRef]
- Piazzon, C.; Lamas, J.; Castro, R.; Budiño, B.; Cabaleiro, S.; Sanmartín, M.; Leiro, J. Antigenic and Cross-Protection Studies on Two Turbot Scuticociliate Isolates. Fish Shellfish Immunol. 2008, 25, 417–424. [Google Scholar] [CrossRef]
- Song, J.Y.; Sasaki, K.; Okada, T.; Sakashita, M.; Kawakami, H.; Matsuoka, S.; Nakayama, K.; Kang, H.S.; Jung, S.J.; Oh, M.J.; et al. Antigenic Differences of the Scuticociliate Miamiensis Avidus from Japan. J. Fish Dis. 2009, 32, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, R.; Paramá, A.; Álvarez, M.F.; Leiro, J.; Sanmartín, M.L. Antiprotozoals Effective in vitro against the Scuticociliate Fish Pathogen Philasterides dicentrarchi. Dis. Aquat. Organ. 2002, 49, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Paramá, A.; Iglesias, R.; Álvarez, F.; Leiro, J.M.; Quintela, J.M.; Peinador, C.; González, L.; Riguera, R.; Sanmartín, M.L. In vitro Efficacy of New Antiprotozoals against Philasterides dicentrarchi (Ciliophora, Scuticociliatida). Dis. Aquat. Organ. 2004, 62, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.-N.; Harikrishnan, R.; Moon, Y.-G.; Kim, M.-C.; Kim, J.-S.; Balasundaram, C.; Heo, M.-S. Effectiveness of Chemotherapeutants against Scuticociliate Philasterides dicentrarchi, a Parasite of Olive Flounder. Vet. Parasitol. 2010, 168, 19–24. [Google Scholar] [CrossRef]
- Quintela, J.M.; Peinador, C.; González, L.; Iglesias, R.; Paramá, A.; Álvarez, F.; Sanmartín, M.L.; Riguera, R. Piperazine N-Substituted Naphthyridines, Pyridothienopyrimidines and Pyridothienotriazines: New Antiprotozoals Active against Philasterides dicentrarchi. Eur. J. Med. Chem. 2003, 38, 265–275. [Google Scholar] [CrossRef]
- Bulfon, C.; Volpatti, D.; Galeotti, M. Current Research on the Use of Plant-Derived Products in Farmed Fish. Aquac. Res. 2015, 46, 513–551. [Google Scholar] [CrossRef]
- Leiro, J.; Arranz, J.A.; Paramá, A.; Álvarez, M.F.; Sanmartín, M.L. In vitro Effects of the Polyphenols Resveratrol, Mangiferin and Epigallocatechin-3-Gallate on the Scuticociliate Fish Pathogen Philasterides dicentrarchi. Dis. Aquat. Organ. 2004, 59, 171–174. [Google Scholar] [CrossRef] [Green Version]
- Budiño, B.; Pata, M.P.; Leiro, J.; Lamas, J. Differences in the in vitro Susceptibility to Resveratrol and Other Chemical Compounds among Several Philasterides dicentrarchi Isolates from Turbot. Parasitol Res. 2012, 110, 1573–1578. [Google Scholar] [CrossRef]
- Mallo, N.; DeFelipe, A.P.; Folgueira, I.; Sueiro, R.A.; Lamas, J.; Leiro, J.M. Combined Antiparasitic and Anti-Inflammatory Effects of the Natural Polyphenol Curcumin on Turbot Scuticociliatosis. J. Fish Dis. 2017, 40, 205–217. [Google Scholar] [CrossRef]
- Mallo, N.; Lamas, J.; Leiro, J.M. Alternative Oxidase Inhibitors as Antiparasitic Agents against Scuticociliatosis. Parasitology 2014, 141, 1311–1321. [Google Scholar] [CrossRef]
- Morais, P.; Piazzon, C.; Lamas, J.; Mallo, N.; Leiro, J.M. Effect of Resveratrol on Oxygen Consumption by Philasterides dicentrarchi, a Scuticociliate Parasite of Turbot. Protist 2013, 164, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Lamas, J.; Leiro, J.M. Scuticociliatosis. In Climate Change and Infectious Fish Diseases; Woo, P.T.K., Leong, J.A., Buchmann, K., Eds.; CABI: Wallingford, UK, 2020; pp. 319–348. [Google Scholar]
- Friedman, M. Anticarcinogenic, Cardioprotective, and Other Health Benefits of Tomato Compounds Lycopene, α -Tomatine, and Tomatidine in Pure Form and in Fresh and Processed Tomatoes. J. Agric. Food Chem. 2013, 61, 9534–9550. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Tomato Glycoalkaloids: Role in the Plant and in the Diet. J. Agric. Food Chem. 2002, 50, 5751–5780. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Kanetake, S.; Wu, Y.H.; Tam, C.; Cheng, L.W.; Land, K.M.; Friedman, M.J. Antiprotozoal Effects of the Tomato Tetrasaccharide Glycoalkaloid Tomatine and the Aglycone Tomatidine on Mucosal Trichomonads. J. Agric. Food Chem. 2016, 64, 8806–8810. [Google Scholar] [CrossRef]
- Tedesco, P.; Beraldo, P.; Massimo, M.; Fioravanti, M.L.; Volpatti, D.; Dirks, R.; Galuppi, R. Comparative Therapeutic Effects of Natural Compounds against Saprolegnia spp. (Oomycota) and Amyloodinium Ocellatum (Dinophyceae). Front. Vet. Sci. 2020, 7, 83. [Google Scholar] [CrossRef]
- Friedman, M.; McQuistan, T.; Hendricks, J.D.; Pereira, C.; Bailey, G.S. Protective Effect of Dietary Tomatine against Dibenzo[a,l]pyrene (DBP)-Induced Liver and Stomach Tumors in Rainbow Trout. Mol. Nutr. Food Res. 2007, 51, 1485–1491. [Google Scholar] [CrossRef]
- Dawid, C.; Weber, D.; Musiol, E.; Janas, V.; Baur, S.; Lang, R.; Fromme, T. Comparative Assessment of Purified Saponins as Permeabilization agents during Respirometry. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148251. [Google Scholar] [CrossRef]
- Padhye, S.; Dandawate, P.; Yusufi, M.; Ahmad, A.; Sarkar, F.H. Perspectives on Medicinal Properties of Plumbagin and Its Analogs. Med. Res. Rev. 2012, 32, 1131–1158. [Google Scholar] [CrossRef]
- Tu, X.; Duan, C.; Wu, S.; Fu, S.; Ye, J. Identification of Plumbagin as an Effective Chemotherapeutic Agent for Treatment of Gyrodactylus Infections. Aquaculture 2021, 535, 736372. [Google Scholar] [CrossRef]
- Rozmer, Z.; Perjési, P. Naturally Occurring Chalcones and Their Biological Activities. Phytochem. Rev. 2016, 15, 87–120. [Google Scholar] [CrossRef]
- Sufian, A.S.; Ramasamy, K.; Ahmat, N.; Zakaria, Z.A.; Yusof, M.I.M. Isolation and Identification of Antibacterial and Cytotoxic Compounds from the Leaves of Muntingia Calabura L. J. Ethnopharmacol. 2013, 146, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Passalacqua, T.G.; Torres, F.A.; Nogueira, C.T.; de Almeida, L.; Del Cistia, M.L.; dos Santos, M.B.; Dutra, L.A.; Bolzani, V.S.; Regasini, L.O.; Graminha, M.A.; et al. The 2′,4′-Dihydroxychalcone Could Be Explored to Develop New Inhibitors against the Glycerol-3-Phosphate Dehydrogenase from Leishmania Species. Bioorg. Med. Chem. Lett. 2015, 25, 3564–3568. [Google Scholar] [CrossRef] [PubMed]
- Lamas, J.; Morais, P.; Arranz, J.A.; Sanmartín, M.L.; Orallo, F.; Leiro, J. Resveratrol Promotes an Inhibitory Effect on the Turbot Scuticociliate Parasite Philasterides dicentrarchi by Mechanisms Related to Cellular Detoxification. Vet. Parasitol. 2009, 161, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Al-Jubury, A.; Lu, C.; Kania, P.W.; von Gersdorff Jørgensen, L.; Liu, Y.; de Bruijn, I.; Raaijmakers, J.; Buchmann, K. Impact of Pseudomonas H6 Surfactant on All External Life Cycle Stages of the Fish Parasitic Ciliate Ichthyophthirius multifiliis. J. Fish Dis. 2018, 41, 1147–1152. [Google Scholar] [CrossRef] [Green Version]
- Jensen, H.M.; Karami, A.M.; Mathiessen, H.; Al-Jubury, A.; Kania, P.W.; Buchmann, K. Gill Amoebae from Freshwater Rainbow Trout (Oncorhynchus mykiss): In vitro Evaluation of Antiparasitic Compounds against Vannella sp. J. Fish Dis. 2020, 43, 665–672. [Google Scholar] [CrossRef]
- Liu, Y.; Rzeszutek, E.; van der Voort, M.; Wu, C.H.; Thoen, E.; Skaar, I.; Bulone, V.; Dorrestein, P.C.; Raaijmakers, J.M.; de Bruijn, I. Diversity of Aquatic Pseudomonas Species and Their Activity against the Fish Pathogenic Oomycete Saprolegnia. PLoS ONE 2015, 10, e0136241. [Google Scholar] [CrossRef]
- Iglesias, R.; Paramá, A.; Álvarez, M.F.; Leiro, J.; Aja, C.; Sanmartín, M.L. In vitro Growth Requeriments for the Fish Pathogen Philasterides dicentrarchi (Ciliophora, Scuticociliatida). Vet. Parasitol. 2003, 111, 19–30. [Google Scholar] [CrossRef]
- Denizot, F.; Lang, R. Rapid Colorimetric Assay for Cell Growth and Survival. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]





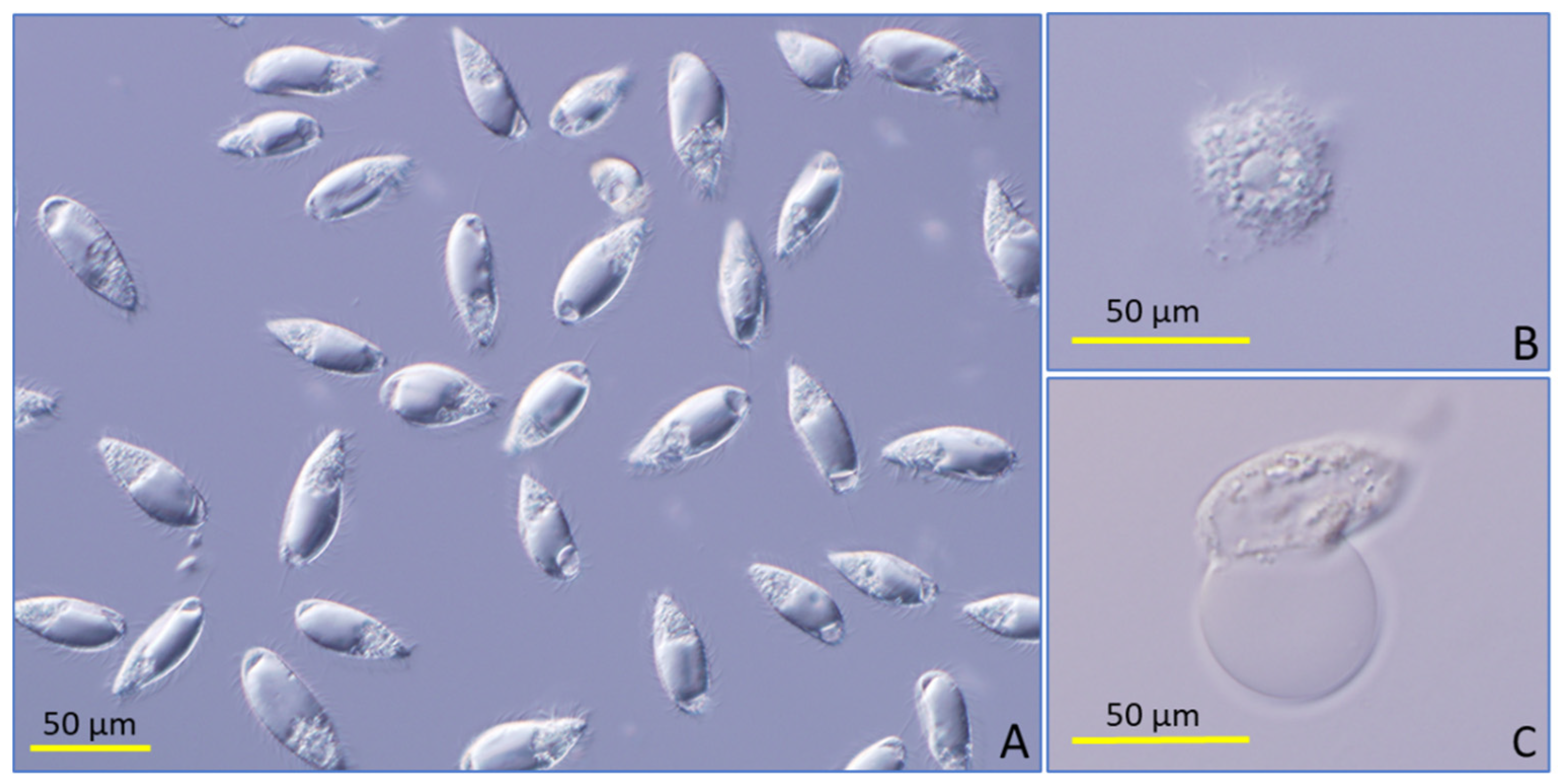{kind=link}
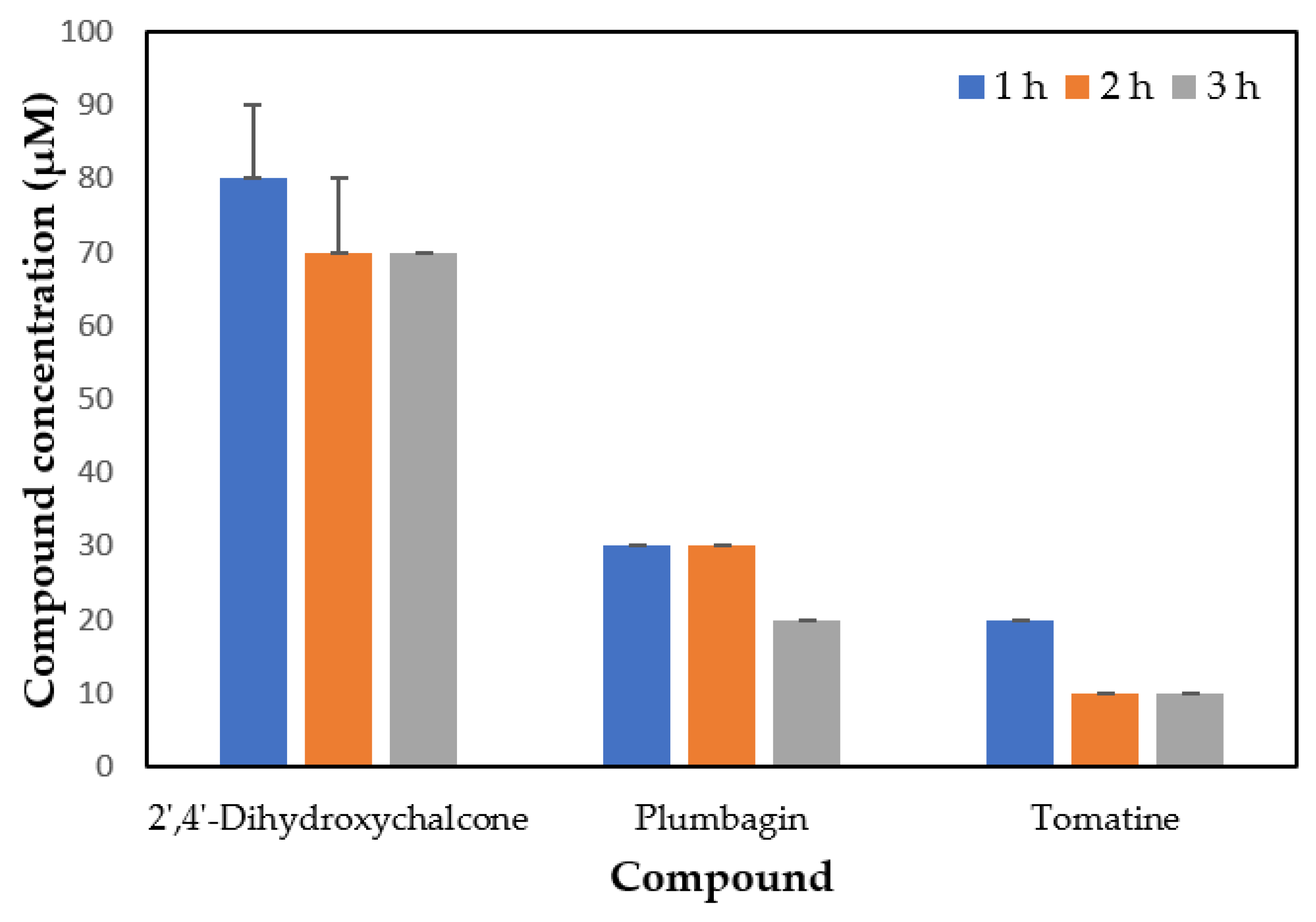{kind=link}
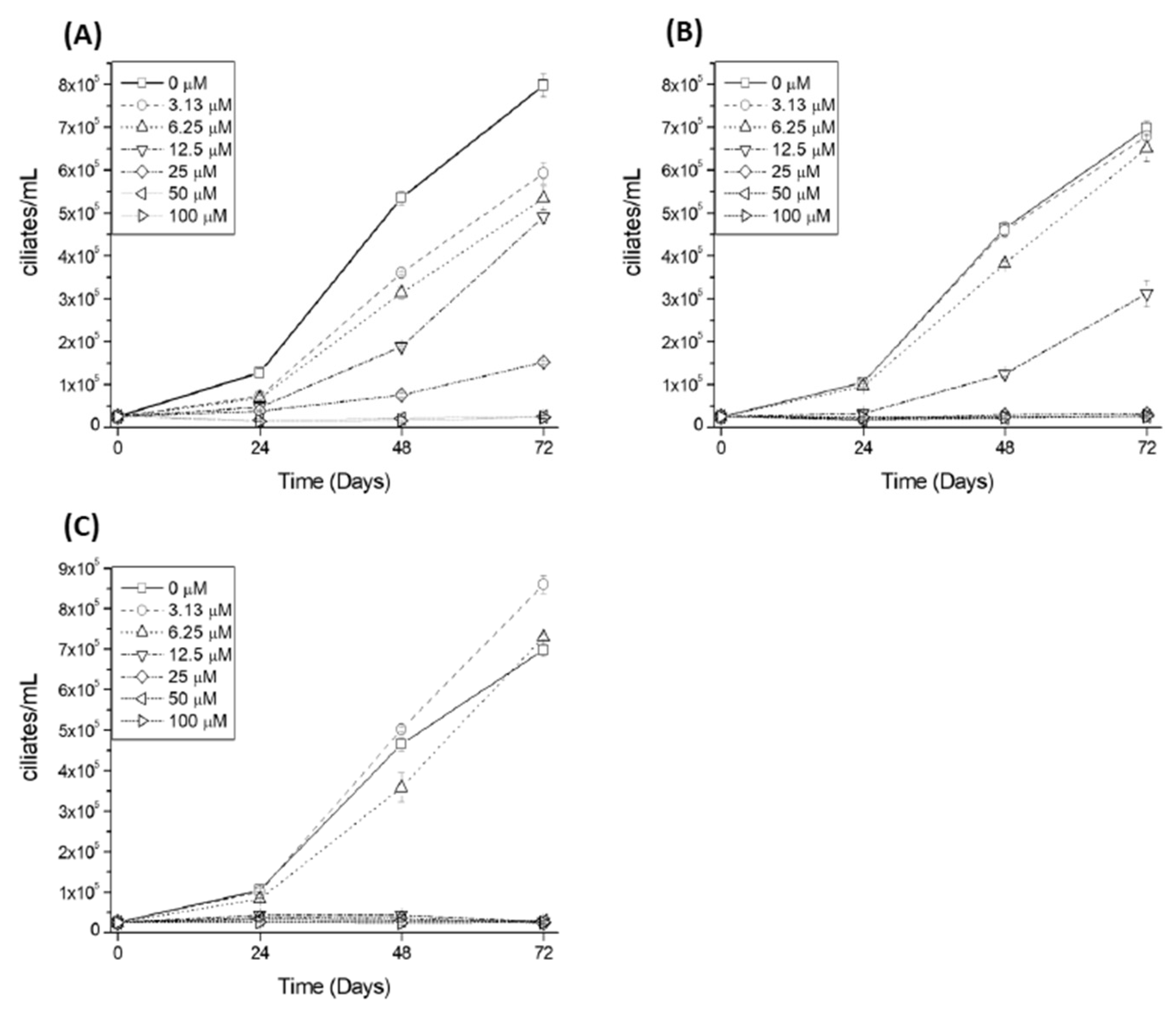{kind=link}
{kind=link}
| Compound | Supplier and Ref. Number | Ciliate Mortality 3 h (%) | Growth Inhibition 24 h (%) |
|---|---|---|---|
| 2′,4′-dihydroxychalcone | SC (sc-266263) | 99 ± 0.71 | 100 |
| 4-hexylresorcinol | S-A (209465) | 95.3 ± 4.05 | 100 |
| 7-hydroxyflavone | S-A (H4530) | 10 ± 1.52 | 67.39 ± 2.4 |
| Benzoic acid | S-A (33045) | 1.64 ± 0.56 | 17.39 ± 1.2 |
| Biotin | S-A (B4501) | 0 | 36.96 ± 0.72 |
| Butyl 4-hydroxybenzoate | S-A (54680) | 52.88 ± 2.39 | 80.65 ± 2.42 |
| Camphor (1r) | S-A (857300) | 0 | 77.42 ± 2.7 |
| Cedrol | S-A (W521418) | 64.25 ± 3.7 | 66.67 ± 0.33 |
| Conessine | SB (11700) | 50.25 ± 4.5 | 84.42 ± 2.42 |
| Coumarin | S-A (C4261) | 8 ± 5.90 | 23.47 ± 4.12 |
| Diallyl sulfide | S-A (W204218) | 22.3 ± 5.1 | 38.76 ± 7.32 |
| Esculetin | S-A (246573) | 8.2 ± 5.32 | 15.31 ± 6.32 |
| Eucalyptol | S-A (C80601) | 17 ± 8.42 | 35.62 ± 4.21 |
| Garlicin 80% | S-A (317691) | 30.02 ± 0.9 | 42.47 ± 5.62 |
| Harmalol hydrochloride dihydrate | S-A (H125-1G-A) | 4.8 ± 1.12 | 40.08 ± 4.9 |
| Monocrolatine | S-A (C2401) | 6.61 ± 1.06 | 9.8 ± 2.11 |
| Palmatine chloride | S-A (sc-205788) | 6.8 ± 0.59 | 28.46 ± 4.25 |
| Phenylethyl alcohol | S-A (W285811) | 2.3 ± 1.4 | 30.14 ± 4.63 |
| Piperine | S-A (P49007) | 42.6 ± 9.1 | 89.04 ± 3.37 |
| Plumbagin | SC (sc-253283) | 100 | 100 |
| Rosmarinic acid | S-A (536954) | 27.12 ± 7.27 | 35 ± 5.51 |
| Sclareolide | S-A (W379401) | 62.64 ± 4.6 | 71.67 ± 6.92 |
| Sodium lactate | S-A (71718) | 13.76 ± 4.1 | 28.33 ± 3.93 |
| Tomatine | SC (sc-296548) | 93.33 ± 2.31 | 100 |
| Umbelliferone | S-A (H24003) | 2.9 ± 1.89 | 44.94 ± 7.23 |
| Usnic acid | S-A (329967) | 38.33 ± 5.52 | 68.3 ± 3.34 |
| Compound | GI50 24 h (µM) | GI50 48 h (µM) |
|---|---|---|
| 2′-4′-Hydroxychalcone | 7.54 ± 1.31 | 5.68 ± 0.16 |
| 4′-Hexylresorcinol | 63.31 ± 3.61 | 40.80 ± 0.11 |
| Butyl-4-hydroxybenzoate | 43.92 ± 6.60 | 29.22 ± 0.35 |
| Piperine | 60.70 ± 6.31 | 43.97 ± 1.83 |
| Plumbagin | 12.89 ± 1.16 | 9.65 ± 0.67 |
| Tomatine | 10.14 ± 1.27 | 8.51 ± 0.20 |
| Conessine | 12.02 ± 0.80 | 11.21 ± 0.65 |
| Compound | GI50 (24 h) | EPC Cell Viability (%) Mean ± SD |
|---|---|---|
| 2′,4′-dihydroxychalcone | 7.54 μM | |
| 3 h | 92.04 ± 2.50 | |
| 24 h | 89.69 ± 1.40 | |
| Plumbagin | 12.89 μM | |
| 3 h | 78.69 ± 1.40 * | |
| 24 h | 74.16 ± 3.94 * | |
| Tomatine | 10.14 μM | |
| 3 h | 62.99 ± 6.82 * | |
| 24 h | 57.63 ± 7.70 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sueiro, R.A.; Leiro, J.M.; Blanco-Abad, V.; Raaijmakers, J.; de Bruijn, I.; Dirks, R.P.H.; Lamas, J. Plant- and Bacteria-Derived Compounds with Anti-Philasterides dicentrarchi Activity. Pathogens 2022, 11, 267. https://doi.org/10.3390/pathogens11020267
Sueiro RA, Leiro JM, Blanco-Abad V, Raaijmakers J, de Bruijn I, Dirks RPH, Lamas J. Plant- and Bacteria-Derived Compounds with Anti-Philasterides dicentrarchi Activity. Pathogens. 2022; 11(2):267. https://doi.org/10.3390/pathogens11020267
Chicago/Turabian StyleSueiro, Rosa Ana, José Manuel Leiro, Verónica Blanco-Abad, Jos Raaijmakers, Irene de Bruijn, Ron P. H. Dirks, and Jesús Lamas. 2022. "Plant- and Bacteria-Derived Compounds with Anti-Philasterides dicentrarchi Activity" Pathogens 11, no. 2: 267. https://doi.org/10.3390/pathogens11020267
APA StyleSueiro, R. A., Leiro, J. M., Blanco-Abad, V., Raaijmakers, J., de Bruijn, I., Dirks, R. P. H., & Lamas, J. (2022). Plant- and Bacteria-Derived Compounds with Anti-Philasterides dicentrarchi Activity. Pathogens, 11(2), 267. https://doi.org/10.3390/pathogens11020267