

Recent Methods for the Viability Assessment of Bacterial Pathogens: Advances, Challenges, and Future Perspectives



Abstract
:1. Introduction
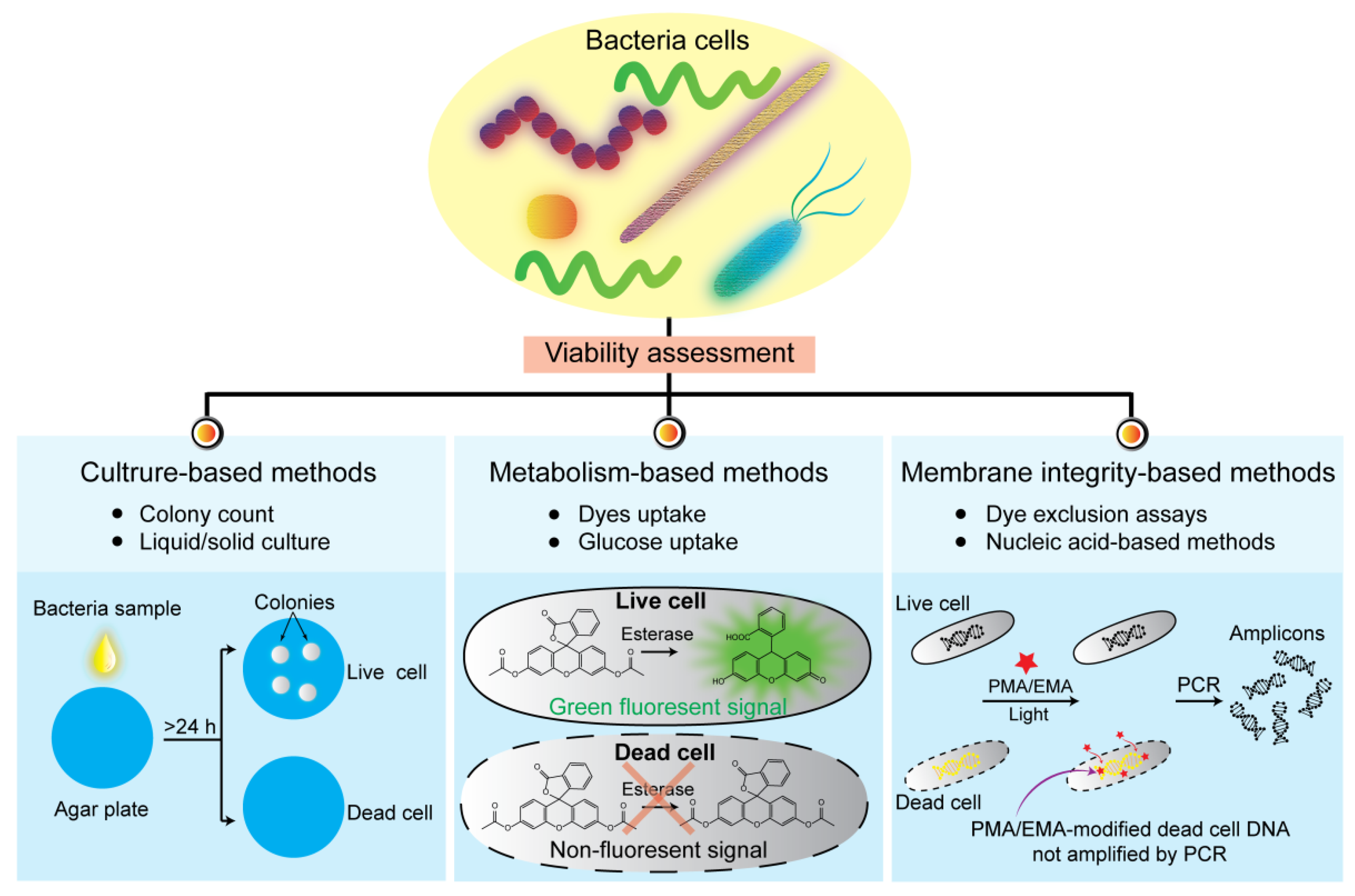
2. Viability Assessments Based on Culturability
3. Viability Assessments Based on Metabolic Activities
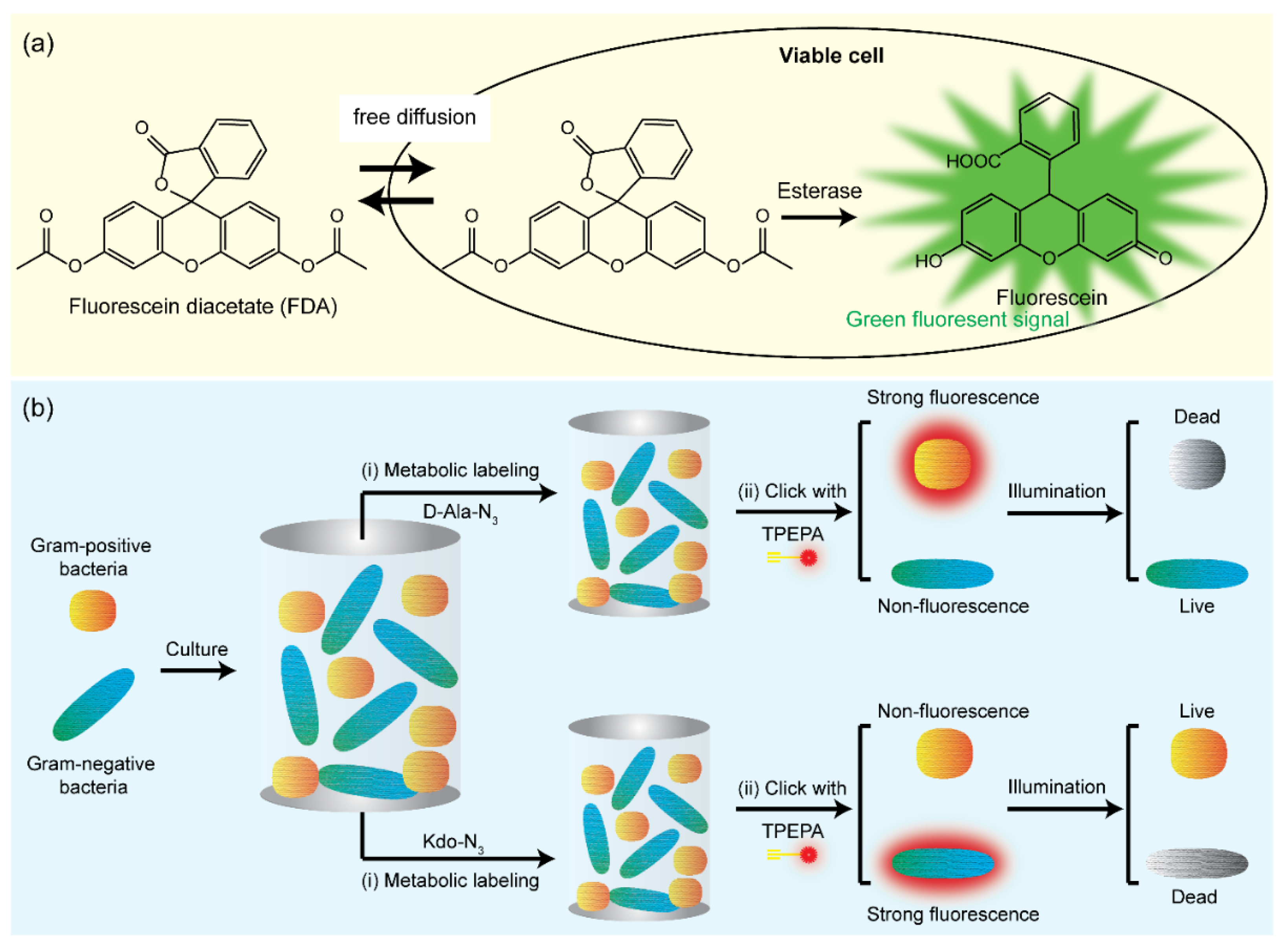
3.1. Dyes Uptake Assay
3.2. Glucose Uptake Assay
3.3. Adenosine Triphosphate (ATP) Assay
4. Viability Assessments Based on Membrane Integrity
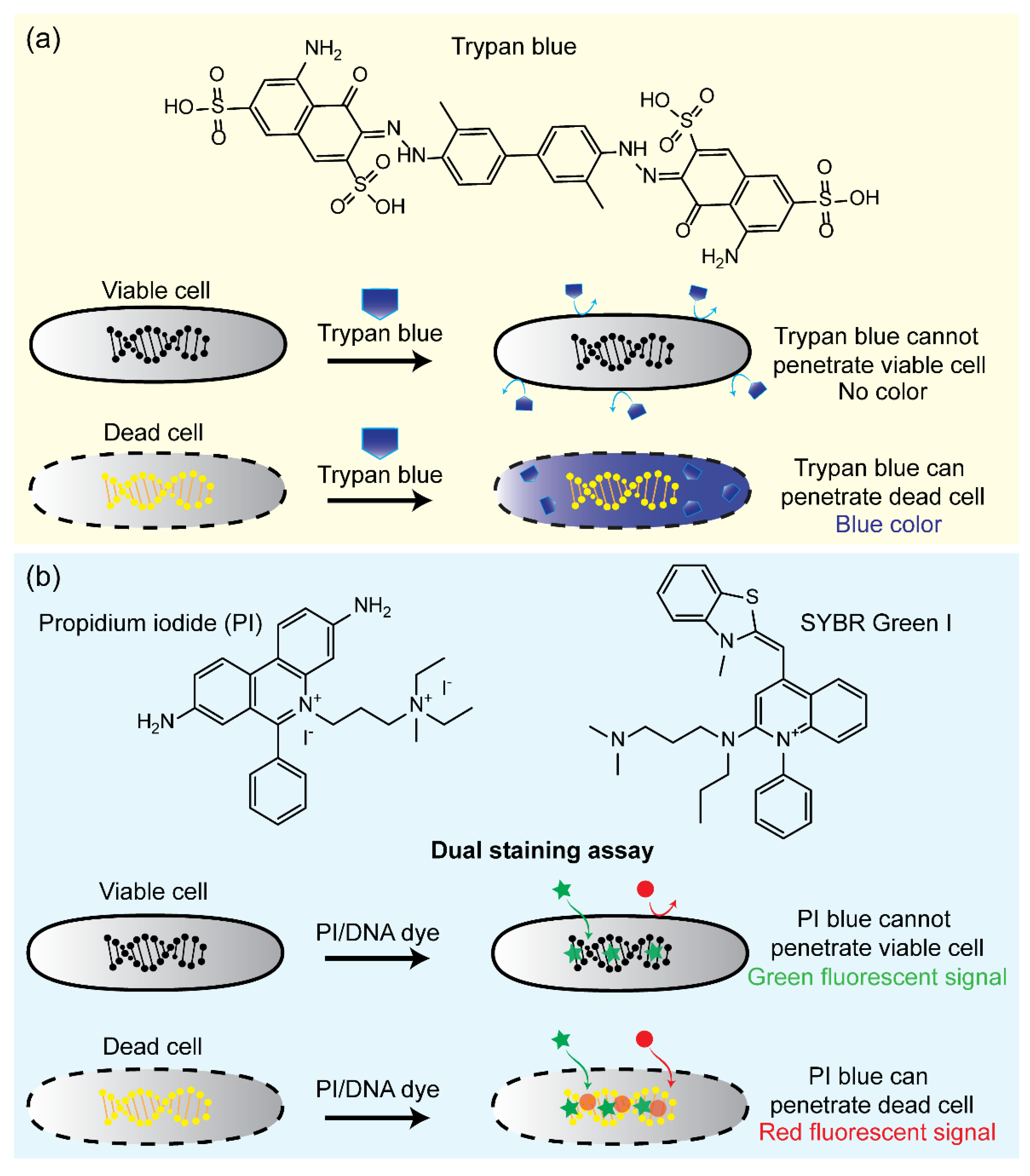
4.1. Dye Exclusion Assays

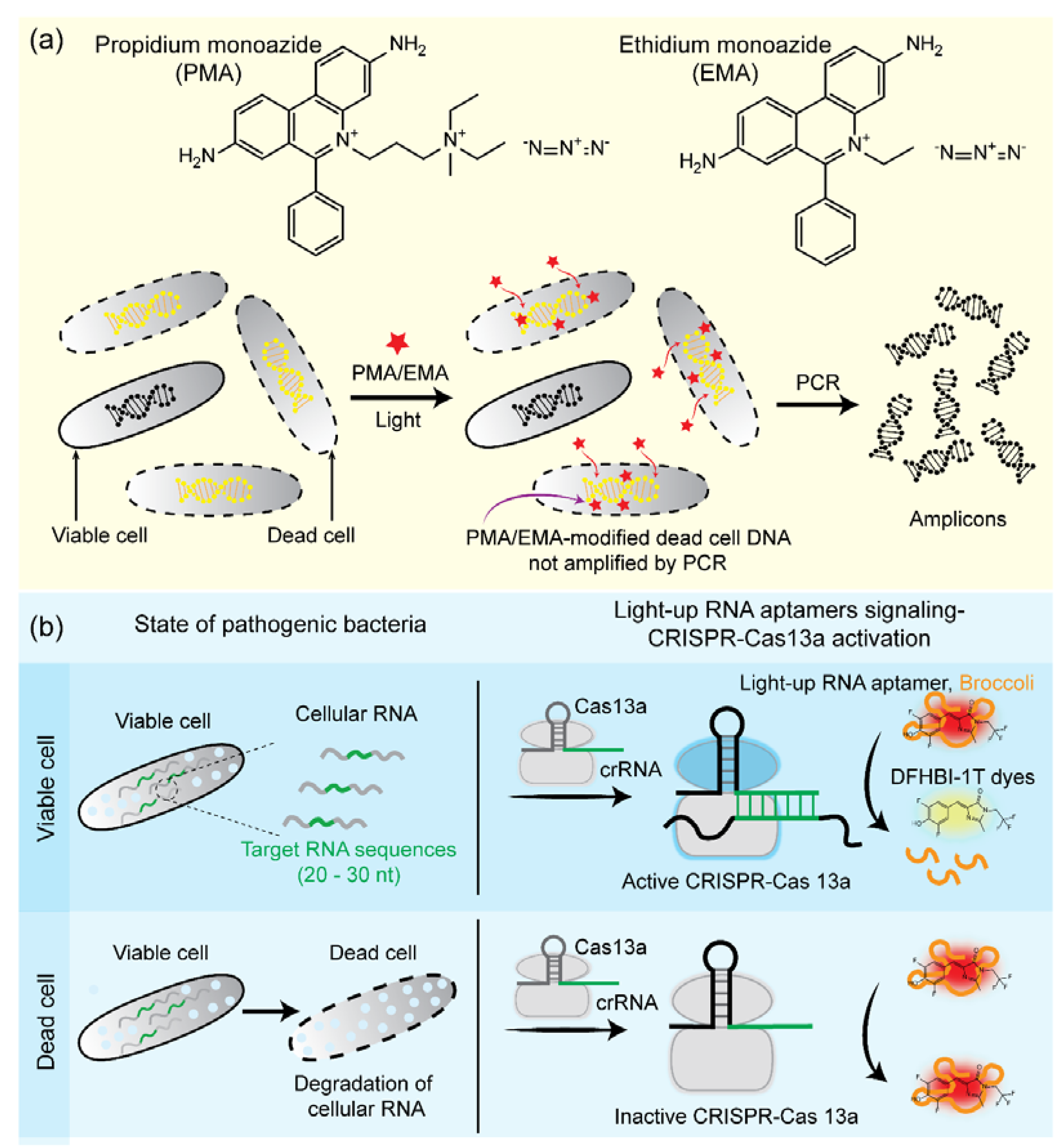
4.2. Nucleic Acid-Based Methods
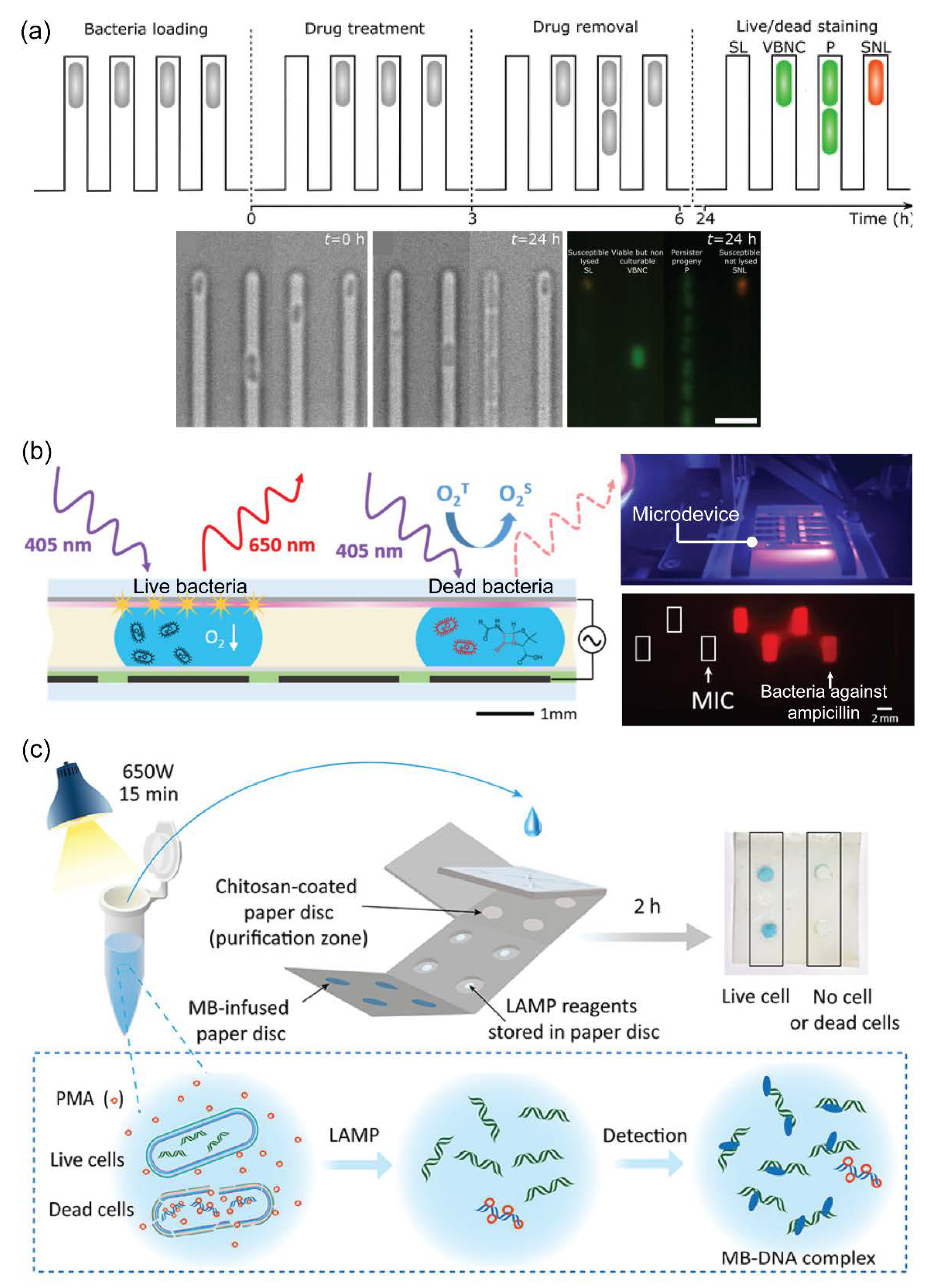
4.3. Microfluidic Technology for Viability Assessments
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nii-Trebi, N.I. Emerging and neglected infectious diseases: Insights, advances, and challenges. BioMed Res. Int. 2017, 2017, 5245021. [Google Scholar] [PubMed]
- Streicher, L.M. Exploring the future of infectious disease treatment in a post-antibiotic era: A comparative review of alternative therapeutics. J. Glob. Antimicrob. Resist. 2021, 24, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Wesche, A.M.; Gurtler, J.B.; Marks, B.P.; Ryser, E.T. Stress, sublethal injury, resuscitation, and virulence of bacterial foodborne pathogens. J. Food Prot. 2009, 72, 1121–1138. [Google Scholar] [CrossRef] [PubMed]
- Verraes, C.; Boxstael, S.V.; Meervenne, E.V.; Coillie, E.V.; Butaye, P.; Catry, B.; Schaetzen, M.-A.; Huffel, X.V.; Imberechts, H.; Dierick, K.; et al. Antimicrobial resistance in the food chain: A review. Int. J. Environ. Res. Public Health 2013, 10, 2643–2669. [Google Scholar] [CrossRef] [PubMed]
- Nocker, A.; Camper, A.K. Novel approaches toward preferential detection of viable cells using nucleic acid amplifcation techniques. FEMS Microbiol. Lett. 2009, 291, 137–142. [Google Scholar] [CrossRef]
- Barbau-Piednoir, E.; Mahillon, J.; Pillyser, J.; Coucke, W.; Nancy, H.; Roosens, N.H.; Botteldoorn, N. Evaluation of viability-qPCR detection system on viable and dead Salmonella serovar Enteritidis. J. Microbiol. Methods 2014, 103, 131–137. [Google Scholar] [CrossRef]
- Biosca, E.G.; Amaro, C.; Marco-Noales, E.; Oliver, J.D. Effect of low temperature on starvation-survival of the eel pathogen Vibrio vulnificus Biotype 2. Appl. Environ. Microbiol. 1996, 62, 450–455. [Google Scholar] [CrossRef]
- Du, M.; Chen, J.; Zhang, X.; Li, A.; Li, Y.; Wang, Y. Retention of virulence in a viable but nonculturable edwardsiella tarda isolate. Appl. Environ. Microbiol. 2007, 73, 1349–1354. [Google Scholar] [CrossRef]
- Fleischmann, S.; Robben, C.; Alter, T.; Rossmanith, P.; Mester, P. How to evaluate non-growing cells—current strategies for determining antimicrobial resistance of VBNC bacteria. Antibiotics 2021, 10, 115. [Google Scholar] [CrossRef]
- Li, L.; Mendis, N.; Trigui, H.; Oliver, J.D.; Sebastien, P.; Faucher, S.P. The importance of the viable but non-culturable state in human bacterial pathogens. Front. Microbiol. 2014, 5, 258. [Google Scholar] [CrossRef] [Green Version]
- Ou, A.; Wang, K.; Mao, Y.; Yuan, L.; Ye, Y.; Chen, L.; Zou, Y.; Huang, T. First report on the rapid detection and identification of methicillin-resistant Staphylococcus aureus (MRSA) in viable but non-culturable (VBNC) under food storage conditions. Front. Microbiol. 2021, 11, 615875. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-S.; Roberts, N.; Singleton, F.L.; Attwell, R.W.; Grimes, D.J.; Colwell, R.R. Survival and viability of nonculturable Escherichia coli and Vibrio cholerae in the estuarine and marine environment. Microb. Ecol. 1982, 8, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yea, C.; Cui, L.; Wan, K.; Chen, S.; Zhang, S.; Yu, X. Population and single cell metabolic activity of UV-induced VBNC bacteria determined by CTC-FCM and D2O-labeled Raman spectroscopy. Environ. Int. 2019, 130, 104883. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Shahamat, M.; Kirchman, P.A.; Russek-Cohen, E.; Colwell, R.R. Methionine uptake and cytopathogenicity of viable but nonculturable Shigella dysenteriae Type 1. Appl. Environ. Microbiol. 1994, 60, 3573–3578. [Google Scholar] [CrossRef]
- Wideman, N.E.; Oliver, J.D.; Crandall, P.G.; Jarvis, N.A. Detection and potential virulence of viable but non-culturable (VBNC) Listeria monocytogenes: A review. Microorganisms 2021, 9, 194. [Google Scholar] [CrossRef]
- Zeng, B.; Zhao, G.; Cao, X.; Yang, Z.; Wang, C.; Hou, L. Formation and resuscitation of viable but nonculturable Salmonella typhi. BioMed Res. Int. 2013, 2013, 907170. [Google Scholar] [CrossRef]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent bacterial infections and persister cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef]
- Wood, T.K.; Knabel, S.J.; Kwan, B.W. Bacterial persister cell formation and dormancy. Appl. Environ. Microbiol. 2013, 79, 7116–7121. [Google Scholar] [CrossRef]
- Grégori, G.; Citterio, S.; Ghiani, A.; Labra, M.; Sgorbati, S.; Brown, S.; Denis, M. Resolution of viable and membrane-compromised bacteria in freshwater and marine waters based on analytical flow cytometry and nucleic acid double staining. Appl. Environ. Microbiol. 2001, 67, 4662–4670. [Google Scholar] [CrossRef]
- Lv, X.-C.; Li, Y.; Qiu, W.-W.; Wu, X.-Q.; Xu, B.-X.; Liang, Y.-T.; Liu, B.; Chen, S.-J.; Rao, P.-F.; Ni, L. Development of propidium monoazide combined with real-time quantitative PCR (PMA-qPCR) assays to quantify viable dominant microorganisms responsible for the traditional brewing of Hong Qu glutinous rice wine. Food Control 2016, 66, 69–78. [Google Scholar] [CrossRef]
- Tan, G.; Zhou, R.; Zhang, W.; Hu, Y.; Ruan, Z.; Li, J.; Zhang, C.; Shen, D.; Peng, N.; Liang, Y.; et al. Detection of viable and total bacterial community in the pit mud of chinese strong-flavor liquor using propidium monoazide combined with quantitative PCR and 16S rRNA gene sequencing. Front. Microbiol. 2020, 11, 896. [Google Scholar] [CrossRef] [PubMed]
- Yun, K.; Park, D.; Kang, M.; Song, J.; Chung, Y.; Bang, H.; Jeon, N.L. A petri-dish with micromolded pattern as a coordinate indicator for live-cell time lapse microscopy. BioChip J. 2022, 16, 27–32. [Google Scholar] [CrossRef]
- Bull, A.T.; Quayle, J.R. New dimensions in microbiology: An introduction. Phil. Trans. R. Soc. Lond. B 1982, 297, 447–457. [Google Scholar]
- Lehnig, M.; Glass, S.; Lippmann, N.; Ziganshyna, S.; Eulenburg, V.; Werdehausen, R. Evaluation of a luminometric cell counting system in context of antimicrobial photodynamic inactivation. Microorganisms 2022, 10, 950. [Google Scholar] [CrossRef]
- Sanders, E.R. Aseptic laboratory techniques: Plating methods. J. Vis. Exp. 2012, 63, e3064. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.M.; Machado, I.; Nicolau, A.; Pereira, M.O. Improvements on colony morphology identification towards bacterial profiling. J. Microbiol. Methods 2013, 95, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Franco-Duarte, R.; Černáková, L.; Kadam, S.; Kaushik, K.S.; Salehi, B.; Bevilacqua, A.; Corbo, M.R.; Antolak, H.; Dybka-Stępién, K.; Leszczewicz, M.; et al. Advances in chemical and biological methods to identify microorganisms—from past to present. Microorganisms 2019, 7, 130. [Google Scholar] [CrossRef]
- Pradhan, P.; Tamang, J.P. Phenotypic and genotypic identification of bacteria isolated from traditionally prepared dry starters of the Eastern Himalayas. Front. Microbiol. 2019, 10, 2526. [Google Scholar] [CrossRef]
- Glasson, J.H.; Guthrie, L.H.; Nielsen, D.J.; Bethell, F.A. Evaluation of an automated instrument for inoculating and spreading samples onto agar plates. J. Clin. Microbiol. 2008, 46, 1281–1284. [Google Scholar] [CrossRef]
- King, G.W.; Kath, G.S.; Siciliano, S.; Simpson, N.; Masurekar, P.; Sigmund, J.; Polishook, J.; Skwish, S.; Bills, G.; Genilloud, O.; et al. Automated agar plate streaker: A linear plater on society for biomolecular sciences standard plates. J. Biomol. Screen. 2006, 11, 704–711. [Google Scholar] [CrossRef]
- Zhu, G.; Yan, B.; Xing, M.; Tian, C. Automated counting of bacterial colonies on agar plates based on images captured at near-infrared light. J. Microbiol. Methods 2018, 153, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Molina, F.; Simancas, A.; Ramírez, M.; Tabla, R.; Roa, I.; Rebollo, J.E. A new pipeline for designing phage cocktails based on phage-bacteria infection networks. Front. Microbiol. 2021, 12, 564532. [Google Scholar] [CrossRef] [PubMed]
- Nitsch, A.; Haralambiev, L.; Einenkel, R.; Muzzio, D.O.; Zygmunt, M.T.; Ekkernkamp, A.; Burchardt, M.; Stope, M.B. Determination of in vitro membrane permeability by analysis of intracellular and extracellular fluorescein signals in renal cells. In Vivo 2019, 33, 1767–1771. [Google Scholar] [CrossRef]
- Hong, D.; Lee, G.; Jung, N.C.; Jeon, M. Fast automated yeast cell counting algorithm using bright-field and fluorescence microscopic images. Biol. Proced. Online 2013, 15, 13. [Google Scholar] [CrossRef]
- Sträuber, H.; Müller, S. Viability states of bacteria—specific mechanisms of selected probes. Cytom. Part A 2010, 77, 623–634. [Google Scholar] [CrossRef]
- Breeuwer, P.; Drocourt, J.-L.; Bunschoten, N.; Zwietering, M.H.; Rombouts, F.M.; Abee, T. Characterization of uptake and hydrolysis of fluorescein diacetate and carboxyfluorescein diacetate by intracellular esterases in saccharomyces cerevisiae, which result in accumulation of fluorescent product. Appl. Environ. Microbiol. 1995, 61, 1614–1619. [Google Scholar] [CrossRef] [PubMed]
- Dzionek, A.; Dzik, J.; Wojcieszyńska, D.; Guzik, U. Fluorescein diacetate hydrolysis using the whole biofilm as a sensitive tool to evaluate the physiological state of immobilized bacterial cells. Catalysts 2018, 8, 434. [Google Scholar] [CrossRef]
- Schnürer, J.; Rosswall, T. Fluorescein diacetate hydrolysis as a measure of total microbial activity in soil and litter. Appl. Environ. Microbiol. 1982, 43, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Qi, G.; Kenry; Mao, D.; Zhou, S.; Wu, M.; Wu, W.; Liu, B. Visualization and in situ ablation of intracellular bacterial pathogens through metabolic labeling. Angew. Chem. 2020, 59, 9288–9292. [Google Scholar] [CrossRef] [PubMed]
- Row, R.D.; Prescher, J.A. Constructing new biorthogonal reagents and reactions. Acc. Chem. Res. 2018, 51, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Tang, B.Z.; Liu, B. Specific light-up bioprobes based on AlEgen conjugates. Chem. Soc. Rev. 2015, 44, 2798–2811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Qi, G.; Liu, X.; Duan, Y.; Liu, J.; Liu, B. Bio-othogonal alegen for specific discrimination and elimination of bacteria pathogens via metabolic engineering. Chem. Mater. 2020, 32, 858–865. [Google Scholar] [CrossRef]
- Jeckelmann, J.-M.; Erni, B. Transporters of glucose and other carbohydrates in bacteria. Pflugers Arch. 2020, 472, 1129–1153. [Google Scholar] [CrossRef] [PubMed]
- Sundar, G.S.; Islam, E.; Braza, R.D.; Silver, A.B.; Breton, Y.L.; McIver, K.S. Route of glucose uptake in the group a streptococcus impacts SLS-mediated hemolysis and survival in human blood. Front. Cell. Infect. Microbiol. 2018, 8, 71. [Google Scholar] [CrossRef]
- Matsuoka, H.; Oishi, K.; Watanabe, M.; Kozone, I.; Saito, M.; Igimi, S. Viable cell detection by the combined use of fluorescent glucose and fluorescent glycine. Biosci. Biotechnol. Biochem. 2003, 67, 2459–2462. [Google Scholar] [CrossRef]
- Tao, J.; McCourt, C.; Sultana, H.; Nelson, C.; Driver, J.; Hackmann, T.J. Use of a fluorescent analog of glucose (2-NBDG) to identify uncultured rumen bacteria that take up glucose. Appl. Environ. Microbiol. 2019, 85, e03018-18. [Google Scholar] [CrossRef]
- Yoshioka, K.; Oh, K.-B.; Saito, M.; Nemoto, Y.; Matsuoka, H. Evaluation of 2-[N-(7-nitrobenz-2-oxa-l,3-diazol-4-yl)amino]-2-deoxy-D. glucose, a new fluorescent derivative of glucose, for viability assessment of yeast Candida albicans. Appl. Microbic. Biotechnol. 1996, 46, 400–404. [Google Scholar]
- Braissant, O.; Astasov-Frauenhoffer, M.; Waltimo, T.; Bonkat, G. A review of methods to determine viability, vitality, and metabolic rates in microbiology. Front. Microbiol. 2020, 11, 547458. [Google Scholar] [CrossRef]
- Silva, P.B.M.; Oliveira, K.A.; Coltro, W.K.T. Colorimetric detection of glucose in biological fluids using toner-based microzone plates. J. Braz. Chem. Soc. 2017, 28, 197–201. [Google Scholar] [CrossRef]
- Xiao, J.; Liu, Y.; Su, L.; Zhao, D.; Zhao, L.; Zhang, X. Microfluidic chip-based wearable colorimetric sensor for simple and facile detection of sweat glucose. Anal. Chem. 2019, 91, 14803–14807. [Google Scholar] [CrossRef]
- Liang, M.; Yan, X. Nanozymes: From new concepts, mechanisms, and standards to applications. Acc. Chem. Res. 2019, 52, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, X.; Huang, L.; Zhang, Z.; Dong, S. GOx@ZIF-8(NiPd) nanoflower: An artificial enzyme system for tandem catalysis. Angew. Chem. 2017, 129, 16298–16301. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, X.; Kang, Y.; Sun, H.; Liang, Y.; Liu, J.; Su, Z.; Dan, J.; Luo, L.; Yue, T.; et al. Natural products self-assembled nanozyme for cascade detection of glucose and bacterial viability in food. Foods 2021, 10, 2596. [Google Scholar] [CrossRef]
- McElroy, W.D. The energy source for bioluminescence in an isolated system. Proc. Natl. Acad. Sci. USA 1947, 33, 342–345. [Google Scholar] [CrossRef]
- Knowles, J.R. Enzyme-catalyzed phosphoryl transfer reactions. Annu. Rev. Biochem. 1980, 49, 877–919. [Google Scholar] [CrossRef] [PubMed]
- Bottari, B.; Santarelli, M.; Neviani, E. Determination of microbial load for different beverages and foodstuff by assessment of intracellular ATP. Trends Food Sci. Technol. 2015, 44, 36–48. [Google Scholar] [CrossRef]
- Hammes, F.; Goldschmidt, F.; Vital, M.; Wang, Y.; Egli, T. Measurement and interpretation of microbial adenosine tri-phosphate (ATP) in aquatic environments. Water Res. 2010, 44, 3915–3923. [Google Scholar] [CrossRef]
- Siragusa, G.R.; Cutter, C.N.; Dorsa, W.J.; Koohmaraie, M. Use of a rapid microbial ATP bioluminescence assay to detect contamination on beef and pork carcasses. J. Food Prot. 1995, 58, 770–775. [Google Scholar] [CrossRef]
- Sakakibara, T.; Murakami, S.; Hattori, N.; Nakajima, M.O.; Imai, K. Enzymatic treatment to eliminate the extracellular ATP for improving the detectability of bacterial intracellular ATP. Anal. Biochem. 1997, 250, 157–161. [Google Scholar] [CrossRef]
- Selan, L.; Berlutti, F.; Passariello, C.; Thaller, M.C.; Renzini, G. Reliability of a bioluminescence ATP assay for detection of bacteria. J. Clin. Microbiol. 1992, 30, 1739–1742. [Google Scholar] [CrossRef]
- Yaginuma, H.; Kawai, S.; Tabata, K.V.; Tomiyama, K.; Kakizuka, A.; Komatsuzaki, T.; Noji, H.; Imamura, H. Diversity in ATP concentrations in a single bacterial cell population revealed by quantitative single-cell imaging. Sci. Rep. 2014, 4, 6522. [Google Scholar] [CrossRef] [PubMed]
- Ammendolia, D.A.; Bement, W.M.; Brumell, J.H. Plasma membrane integrity: Implications for health and disease. BMC Biology 2021, 19, 71. [Google Scholar] [CrossRef] [PubMed]
- Breeuwer, P.; Abee, T. Assessment of viability of microorganisms employing fluorescence techniques. Int. J. Food Microbiol. 2000, 55, 193–200. [Google Scholar] [CrossRef]
- Kwizera, R.; Akampurira, A.; Kandole, T.K.; Nielsen, K.; Kambugu, A.; Meya, D.B.; Boulware, D.R.; Rhein, J. Evaluation of trypan blue stain in a haemocytometer for rapid detection of cerebrospinal fluid sterility in HIV patients with cryptococcal meningitis. BMC Microbiol. 2017, 17, 182. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, C.N.; Antczak, C.; Djaballah, H. Cell viability assessment: Toward content-rich platforms. Expert Opin. Drug Discov. 2010, 5, 223–233. [Google Scholar] [CrossRef]
- Kerschbauma, H.H.; Tasa, B.A.; Schürz, M.; Oberascher, K.; Bresgen, N. Trypan blue—Adapting a dye used for labelling dead cells to visualize pinocytosis in viable cells. Cell. Physiol. Biochem. 2021, 55, 171–184. [Google Scholar]
- Tran, S.-L.; Puhar, A.; Ngo-Camus, M.; Ramarao, N. Trypan blue dye enters viable cells incubated with the pore-forming toxin HlyII of Bacillus cereus. PLoS ONE 2011, 6, e22876. [Google Scholar] [CrossRef]
- Bresgen, N.; Ohlenschläger, I.; Wacht, N.; Afazel, S.; Ladurner, G.; Eckl, P.M. Ferritin and FasL (CD95L) mediate density dependent apoptosis in primary rat hepatocytes. J. Cell. Physiol. 2008, 217, 800–808. [Google Scholar] [CrossRef]
- Hu, C.; He, S.; Lee, Y.J.; He, Y.; Kong, E.M.; Li, H.; Anastasio, M.A.; Popescu, G. Live-dead assay on unlabeled cells using phase imaging with computational specificity. Nat. Commun. 2022, 13, 713. [Google Scholar] [CrossRef]
- Louis, K.S.; Siegel, A.C. Cell viability analysis using trypan blue: Manual and automated methods. Methods Mol. Biol. 2011, 740, 7–12. [Google Scholar]
- Franke, J.D.; Braverman, A.L.; Cunningham, A.M.; Eberhard, E.E.; Perry, G.A. Erythrosin B: A versatile colorimetric and fluorescent vital dye for bacteria. BioTechniques 2019, 68, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Silva, K.J.S.; Sabogal-Paz, L.P. Cryptosporidium spp. and Giardia spp. (oo)cysts as target-organism in sanitation and environmental monitoring: A review in microscopy-based viability assays. Water Res. 2021, 189, 116590. [Google Scholar] [CrossRef] [PubMed]
- Davey, H.; Guyot, S. Estimation of microbial viability using flow cytometry. Curr. Protoc. Cytom. 2020, 93, e72. [Google Scholar] [CrossRef] [PubMed]
- McEvoy, B.; Lynch, M.; Rowan, N.J. Opportunities for the application of real-time bacterial cell analysis using flow cytometry for the advancement of sterilization microbiology. J. Appl. Microbiol. 2020, 130, 1794–1812. [Google Scholar] [CrossRef] [PubMed]
- Boulos, L.; Prevost, M.; Barbeau, B.; Coallier, J.; Desjardins, R. LIVE/DEAD® BacLight™: Application of a new rapid staining method for direct enumeration of viable and total bacteria in drinking water. J. Microbiol. Methods 1999, 37, 77–86. [Google Scholar] [CrossRef]
- Stiefel, P.; Schmidt-Emrich, S.; Maniura-Weber, K.; Ren, Q. Critical aspects of using bacterial cell viability assays with the fluorophores SYTO9 and propidium iodide. BMC Microbiol. 2015, 15, 36. [Google Scholar] [CrossRef]
- Khan, M.M.T.; Pyle, B.H.; Camper, A.K. Specific and rapid enumeration of viable but nonculturable and viable-culturable gram-negative bacteria by using flow cytometry. Appl. Environ. Microbiol. 2010, 76, 5088–5096. [Google Scholar] [CrossRef]
- Feng, J.; Yee, R.; Zhang, S.; Tian, L.; Shi, W.; Zhang, W.-H.; Zhang, Y. A Rapid growth-independent antibiotic resistance detection test by SYBR Green/propidium iodide viability assay. Front. Med. 2018, 5, 127. [Google Scholar] [CrossRef]
- Freire, J.M.; Gaspar, D.; Torre, B.G.; Veiga, A.S.; Andreu, D.; Castanho, M.A.R.B. Monitoring antibacterial permeabilization in real time using time-resolved flow cytometry. Biochim. Biophys. Acta 2015, 1848, 554–560. [Google Scholar] [CrossRef]
- Kolek, J.; Branska, B.; Drahokoupil, M.; Patakova, P.; Melzoch, K. Evaluation of viability, metabolic activity and spore quantity in clostridial cultures during ABE fermentation. FEMS Microbiol. Lett. 2016, 363, fnw031. [Google Scholar] [CrossRef]
- Rosenberg, M.; Azevedo, N.F.; Ivask, A. Propidium iodide staining underestimates viability of adherent bacterial cells. Sci. Rep. 2019, 9, 6483. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ateia, M.; Hatano, Y.; Miyanagad, K.; Yoshimura, C. Novel fluorescence-based method for rapid quantification of live bacteria in river water and treated wastewater. Environ. Sci. Adv. 2022, 1, 30. [Google Scholar] [CrossRef]
- Kontchou, J.A.; Nocker, A. Optimization of viability qPCR for selective detection of membrane-intact Legionella pneumophila. J. Microbiol. Methods 2019, 156, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, J.N.; Cady, N.C. Surveilling cellular vital signs: Toward label-free biosensors and real-time viability assays for bioprocessing. Curr. Opin. Biotechnol. 2021, 71, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Leifels, M.; Cheng, D.; Sozzi, E.; Shoults, D.C.; Wuertz, S.; Mongkolsuk, S.; Sirikanchana, K. Capsid integrity quantitative PCR to determine virus infectivity in environmental and food applications—A systematic review. Water Res. X 2021, 11, 100080. [Google Scholar] [CrossRef]
- Xie, G.; Zhou, D.; Zhao, G.; Feng, X.; Aguilar, Z.P.; Xu, H. Recombinase aided amplification with photoreactive DNA-binding dye for rapid detection of viable Staphylococcus aureus. LWT 2021, 135, 110249. [Google Scholar] [CrossRef]
- Cawthorn, D.M.; Witthuhn, R.C. Selective PCR detection of viable Enterobacter sakazakii cells utilizing propidium monoazide or ethidium bromide monoazide. J. Appl. Microbiol. 2008, 105, 1178–1185. [Google Scholar] [CrossRef]
- Stinson, L.F.; Keelan, J.A.; Payne, M.S. Characterization of the bacterial microbiome in first-pass meconium using propidium monoazide (PMA) to exclude nonviable bacterial DNA. Lett. Appl. Microbiol. 2019, 68, 378–385. [Google Scholar] [CrossRef]
- Vondrakova, L.; Turonova, H.; Scholtz, V.; Pazlarova, J.; Demnerova, K. Impact of various killing methods on EMA/PMA qPCR efficacy. Food Control 2018, 85, 23–28. [Google Scholar] [CrossRef]
- Cao, X.; Zhao, L.; Zhang, J.; Chen, X.; Shi, L.; Fang, X.; Xie, H.; Chang, Y.; Wang, L. Detection of viable but nonculturable Vibrio parahaemolyticus in shrimp samples using improved real-time PCR and real-time LAMP methods. Food Control 2019, 103, 145–152. [Google Scholar] [CrossRef]
- Gao, S.; Sun, C.; Hong, H.; Gooneratne, R.; Mutukumira, A.; Wu, X. Rapid detection of viable Cronobacter sakazakii in powdered infant formula using improved propidium monoazide (PMAxx) and quantitative recombinase polymerase amplification (qRPA) assay. Food Control 2021, 124, 107899. [Google Scholar] [CrossRef]
- Mu, D.; Zhou, D.; Xie, G.; Liu, J.; Wang, Z.; Xiong, Q.; Xu, H. Real-time recombinase-aided amplification with improved propidium monoazide for the rapid detection of viable Escherichia coli O157:H7 in milk. J. Dairy Sci. 2022, 105, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Hein, I.; Schneeweiss, W.; Stanek, C.; Wagner, M. Ethidium monoazide and propidium monoazide for elimination of unspecific DNA background in quantitative universal real-time PCR. J. Microbiol. Methods 2007, 71, 336–339. [Google Scholar] [CrossRef]
- Nocker, A.; Cheung, C.-Y.; Camper, A.K. Comparison of propidium monoazide with ethidium monoazide for differentiation of live vs. dead bacteria by selective removal of DNA from dead cells. J. Microbiol. Methods 2006, 67, 310–320. [Google Scholar] [CrossRef]
- Lee, A.S.; Lamanna, O.K.; Ishida, K.; Hill, E.; Nguyen, A.; Hsieh, M.H. A novel propidium monoazide-based PCR assay can measure viable uropathogenic E. coli in vitro and in vivo. Front. Cell. Infect. Microbiol. 2022, 12, 794323. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.J.; Bentham, R.H.; Ross, K.E. Limitations of using propidium monoazide with qPCR to discriminate between live and dead legionella in biofilm samples. Microbiol. Insights 2014, 4, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Santander, R.D.; Meredith, C.L.; Aćimović, S.G. Development of a viability digital PCR protocol for the selective detection and quantification of live Erwinia amylovora cells in cankers. Sci. Rep. 2019, 9, 11530. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, W.; Xu, H.; Aguilar, Z.P.; Shah, N.P.; Wei, H. Propidium monoazide combined with real-time PCR for selective detection of viable Staphylococcus aureus in milk powder and meat products. Int. J. Dairy Sci. 2015, 98, 1625–1633. [Google Scholar] [CrossRef]
- Zhang, Z.; Feng, L.; Xu, H.; Liu, C.; Shah, N.P.; Wei, H. Detection of viable enterotoxin-producing Bacillus cereus and analysis of toxigenicity from ready-to-eat foods and infant formula milk powder by multiplex PCR. J. Dairy Sci. 2016, 99, 1047–1055. [Google Scholar] [CrossRef]
- Chang, C.-W.; Lin, M.-H. Optimization of PMA-qPCR for Staphylococcus aureus and determination of viable bacteria in indoor air. Int. J. Indoor Environ. Health 2018, 28, 64–72. [Google Scholar]
- Rey, M.d.l.A.; Racca, A.R.; Ribeiro, L.R.; Cruz, F.D.S.; Cap, M.; Mozgovoj, M.V.; Cristianini, M.; Vaudagna, S.R. High-pressure processing treatment of beef burgers: Effect on Escherichia coli O157 inactivation evaluated by plate count and PMA-qPCR. J. Food Sci. 2022, 87, 2324–2335. [Google Scholar] [CrossRef] [PubMed]
- Golpayegani, A.; Douraghi, M.; Rezaei, F.; Alimohammadi, M.; Nodehi, R.N. Propidium monoazide-quantitative polymerase chain reaction (PMA-qPCR) assay for rapid detection of viable and viable but non-culturable (VBNC) Pseudomonas aeruginosa in swimming pools. J. Environ. Health Sci. Eng. 2019, 17, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, Y.; Shu, Y.; Xia, W.; Xu, R.; Chen, Y. Modified PMA-qPCR method for repaid quantification of viable Lastobacillus spp. in fermented dairy products. Food Anal. Methods 2021, 14, 1908–1918. [Google Scholar] [CrossRef]
- Dorn-In, S.; Gareis, M.; Schwaiger, K. Differentiation of live and dead Mycobacterium tuberculosis complex in meat samples using PMA-qPCR. Food Microbiol. 2019, 84, 103275. [Google Scholar] [CrossRef]
- Zhou, P.; Xie, G.; Liang, T.; Aguilar, Z.; Xu, H. Rapid and quantitative detection of viable emetic Bacillus cereus by PMA-qPCR assay in milk. Mol. Cell. Probes 2019, 47, 101437. [Google Scholar] [CrossRef]
- Gou, J.; Wang, W.; Zhao, H.; Luo, Y.; Wan, M.; Li, Y. A new PMA-qPCR method for rapid and accurate detection of viable bacteria and spores of marine-derived Bacillus velezensis B-9987. J. Microbiol. Methods 2022, 199, 106537. [Google Scholar]
- Li, L.; Fu, J.; Bae, S. Changes in physiology states of Salmonella Typhimurium measured by qPCR with PMA and DyeTox13 Green Azide after pasteurization and UV treatment. Appl. Microbiol. Biotechnol. 2022, 106, 2739–2750. [Google Scholar] [CrossRef]
- Han, L.; Wang, K.; Ma, L.; Delaquis, P.; Bach, S.; Feng, J.; Lu, X. Viable but nonculturable Escherichia coli O157:H7 and Salmonella enterica in fresh produce: Rapid determination by loop-mediated isothermal amplification couple with a propidium monoazide treatment. Food Microbiol. 2020, 86, e02566-19. [Google Scholar] [CrossRef]
- Li, R.; Tun, H.M.; Jahan, M.; Zhang, Z.; Kumar, A.; Dilantha Fernando, W.G.; Farenhorst, A.; Khafpour, E. Comparison of DNA-, PMA-, and RNA-based 16S rRNA illumine sequencing for detection of live bacteria in water. Sci. Res. 2017, 7, 5752. [Google Scholar]
- Lee, S.; Bae, S. Molecular viability testing of viable by non-culturable bacteria induced by antibiotic exposure. Microb. Biotechnol. 2018, 11, 1008–1016. [Google Scholar] [CrossRef]
- Trinh, T.N.D.; Lee, N.Y. Colorimetric detection of viable antibiotic resistant Enterococcus mediated by cordless operation of reverse transcription loop-mediated isothermal amplification. J. Biotechnol. 2022, 357, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhou, W.; Lin, X.; Khan, M.R.; Deng, S.; Zhou, M.; He, G.; Wu, C.; Deng, R.; He, Q. Light-up RNA aptamer signaling-CRISPE-Cas13a-based mix-and-read assays for profiling viable pathogenic bacteria. Biosens. Bioelectron. 2021, 176, 112906. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Tao, Z.; Wan, L.; Zong, C.; Wu, J.; Tan, X.; Wang, B.; Gou, Z.; Zhang, L.; Yuan, H.; et al. Aptamer-based Cas14a1 biosensor for amplification-free live pathogenic detection. Biosens. Bioelectron. 2022, 211, 114282. [Google Scholar] [CrossRef] [PubMed]
- Yue, M.; Schmieder, R.; Edwards, R.A.; Rankin, S.C.; Schifferli, D.M. Microfluidic PCR combined with pyrosequencing for identification of allelic variants with phenotypic associations among targeted Salmonella genes. Appl. Environ. Microbiol. 2012, 78, 7480–7482. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liu, Y.; Yin, J.; Yue, M.; Mu, Y. Microfluidic devices for multiplexed detection of foodborne pathogens. Food Res. Int. 2021, 143, 110246. [Google Scholar] [CrossRef]
- Zhao, X.; Li, M.; Liu, Y. Microfluidic-based approaches for foodborne pathogen detection. Microorganisms 2019, 7, 381. [Google Scholar] [CrossRef]
- Zhang, D.; Bi, H.; Liu, B.; Qiao, L. Detection of pathogenic microorganisms by microfluidics based analytical methods. Anal. Chem. 2018, 90, 5512–5520. [Google Scholar] [CrossRef]
- Kang, S.-M. Recent advances in microfluidic-based microphysiological systems. BioChip J. 2022, 16, 13–26. [Google Scholar] [CrossRef]
- Chang, W.-H.; Wang, C.-H.; Yang, S.-Y.; Lin, Y.-C.; Wu, J.-J.; Lee, M.S.; Lee, G.-B. Rapid isolation and diagnosis of live bacteria from human joint fluids by using an integrated microfluidic system. Lab Chip 2014, 14, 3376–3384. [Google Scholar] [CrossRef]
- Wang, C.-H.; Chang, C.-J.; Wu, J.-J.; Lee, G.-B. An integrated microfluidic device utilizing vancomycin conjugated magnetic beads and nanogold-labeled specific nucleotide probes for rapid pathogen diagnosis. Nanomedicine 2014, 10, 809–818. [Google Scholar] [CrossRef]
- Bamford, R.A.; Smith, A.; Metz, J.; Glover, G.; Titball, R.W.; Pagliara, S. Investigating the physiology of viable but non-culturable bacteria by microfluidics and time-lapse microscopy. BMC Biol. 2017, 15, 121. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Nagl, S. Automated miniaturized digital microfluidic antimicrobial susceptibility test using a chip-integrated optical oxygen sensor. ACS Sens. 2021, 6, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-H.; Wang, C.-H.; Lin, C.-L.; Wu, J.-J.; Lee, M.S.; Lee, G.-B. Rapid detection and typing of live bacteria from human joint fluid samples by utilizing an integrated microfluidic system. Biosens. Bioelectron. 2015, 66, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Etayash, H.; Khan, M.F.; Kaur, K.; Thundat, T. Microfluidic cantilever detects bacteria and measures their susceptibility to antibiotics in small confined volumes. Nat. Commun. 2016, 7, 12947. [Google Scholar] [CrossRef] [PubMed]
- Trieu, P.T.; Lee, N.Y. Paper-based all-in-one origami microdevice for nucleic acid amplification testing for rapid colorimetric identification of live cells for point-of-care testing. Anal. Chem. 2019, 91, 11013–11022. [Google Scholar] [CrossRef]
- Zoval, J.V.; Madou, M.J. Centrifuge-based fluidic platforms. Proc. IEEE 2004, 92, 140–153. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, X.; Xie, X.; Bahnemann, J.; Lin, X.; Wu, X.; Wang, S.; Hoffmann, M.R. Propidium monoazide pretreatment on a 3D-printed microfluidic device for efficient PCR determination of “live versus dead” microbial cells. Environ. Sci. Water Res. Technol. 2018, 4, 956–963. [Google Scholar] [CrossRef] [Green Version]
- Iacobino, A.; Plccaro, G.; Glannonl, F.; Mustazzolu, A.; Fattorlnl, L. Mycobacterium tuberculosis is selectively killed by rifampin and rifapentine in hypoxia at neutral pH. Antimicrob. Agents Chemother. 2017, 61, e02296-16. [Google Scholar] [CrossRef]
- Wang, C.-H.; Chang, J.-R.; Hung, S.-C.; Dou, H.-Y.; Lee, G.-B. Rapid molecular diagnosis of live Mycobacterium tuberculosis on an integrated microfluidic system. Sens. Actuators B Chem. 2022, 365, 131968. [Google Scholar] [CrossRef]
- Ou, F.; McGoverin, C.; Swift, S.; Vanholsbeeck, F. Rapid and cost-effective evaluation of bacterial viability using fluorescence spectroscopy. Anal. Bioanal. Chem. 2019, 411, 3653–3663. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Principle | Advantages | Disadvantages | Cost Estimation | Ref. |
|---|---|---|---|---|---|
| Culture-based method |
|
|
|
| [24,25,26] |
| Metabolism-based method |
|
|
|
| [33,42,45,46,47,130] |
| Membrane integrity-based method |
|
|
|
| [70,71,72,73,74,97,98,99,100,101,102,103,130] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trinh, K.T.L.; Lee, N.Y. Recent Methods for the Viability Assessment of Bacterial Pathogens: Advances, Challenges, and Future Perspectives. Pathogens 2022, 11, 1057. https://doi.org/10.3390/pathogens11091057
Trinh KTL, Lee NY. Recent Methods for the Viability Assessment of Bacterial Pathogens: Advances, Challenges, and Future Perspectives. Pathogens. 2022; 11(9):1057. https://doi.org/10.3390/pathogens11091057
Chicago/Turabian StyleTrinh, Kieu The Loan, and Nae Yoon Lee. 2022. "Recent Methods for the Viability Assessment of Bacterial Pathogens: Advances, Challenges, and Future Perspectives" Pathogens 11, no. 9: 1057. https://doi.org/10.3390/pathogens11091057
APA StyleTrinh, K. T. L., & Lee, N. Y. (2022). Recent Methods for the Viability Assessment of Bacterial Pathogens: Advances, Challenges, and Future Perspectives. Pathogens, 11(9), 1057. https://doi.org/10.3390/pathogens11091057