

Prevalence and Molecular Characterization of Shiga Toxin-Producing Escherichia coli from Food and Clinical Samples

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. DNA Isolation from Bacteria
2.3. Serotyping of STEC Strains
2.4. Antimicrobial Susceptibility Testing by Disk Diffusion
2.5. Pulsed-Field Gel Electrophoresis (PFGE)
2.6. Identification of Shiga-Toxin Subtype and Other Genes Associated with Virulence
2.7. Plasmid Profiling
2.8. PCR Detection of Plasmid Replicon Typing
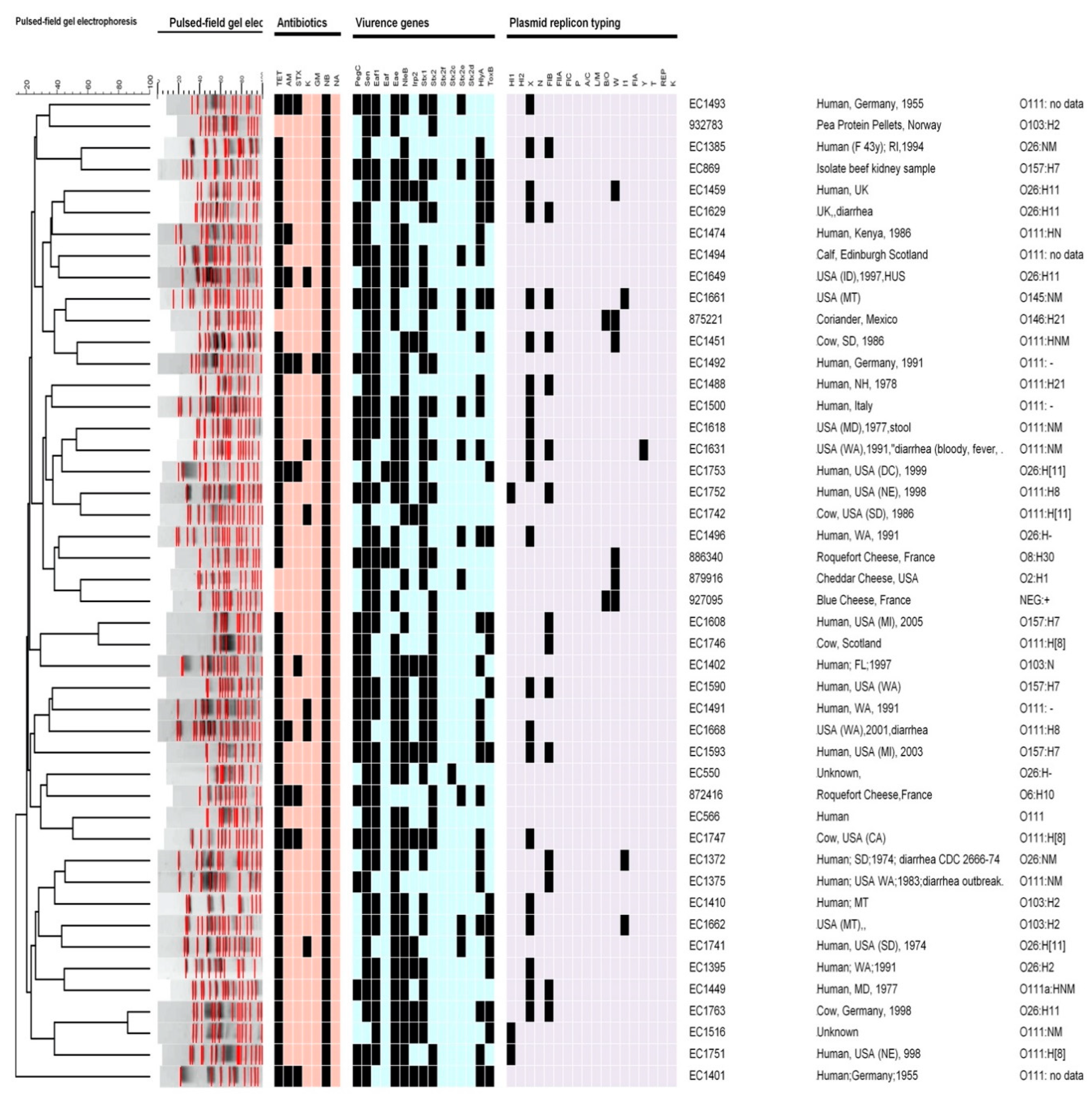
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Griffin, P.M.; Tauxe, R.V. The epidemiology of infections caused by Escherichia coli O157:H7, other enterohemorrhagic E. coli, and the associated hemolytic uremic syndrome. Epidemiol. Rev. 1991, 13, 60–98. [Google Scholar] [CrossRef] [PubMed]
- Law, D. Virulence factors of Escherichia coli O157 and other Shiga toxin-producing E. coli. J. Appl. Microbiol. 2000, 88, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Monaghan, A.M.; Byrne, B.; McDowell, D.; Carroll, A.M.; McNamara, E.B.; Bolton, D.J. Characterization of farm, food, and clinical Shiga toxin-producing Escherichia coli (STEC) O113. Foodborne Pathog. Dis. 2012, 9, 1088–1096. [Google Scholar] [CrossRef] [PubMed]
- Douellou, T.; Delannoy, S.; Ganet, S.; Fach, P.; Loukiadis, E.; Montel, M.C.; Sergentet-Thevenot, D. Molecular characterization of O157:H7, O26:H11 and O103:H2 Shiga toxin-producing Escherichia coli isolated from dairy products. Int. J. Food Microbiol. 2017, 253, 59–65. [Google Scholar] [CrossRef]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar]
- Balière, C.; Rincé, A.; Blanco, J.; Dahbi, G.; Harel, J.; Vogeleer, P.; Giard, J.C.; Mariani-Kurkdjian, P.; Gourmelon, M. Prevalence and Characterization of Shiga Toxin-Producing and Enteropathogenic Escherichia coli in Shellfish-Harvesting Areas and Their Watersheds. Front. Microbiol. 2015, 6, 1356. [Google Scholar] [CrossRef]
- O’Brien, A.D.; Newland, J.W.; Miller, S.F.; Holmes, R.K.; Smith, H.W.; Formal, S.B. Shiga-like toxin-converting phages from Escherichia coli strains that cause hemorrhagic colitis or infantile diarrhea. Science 1984, 226, 694–696. [Google Scholar] [CrossRef]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Piérard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef]
- Friedrich, A.W.; Bielaszewska, M.; Zhang, W.-L.; Pulz, M.; Kuczius, T.; Ammon, A.; Karch, H. Escherichia coli harboring Shiga toxin 2 gene variants: Frequency and association with clinical symptoms. J. Infect. Dis. 2002, 185, 74–84. [Google Scholar] [CrossRef]
- Melton-Celsa, A.R. Shiga Toxin (Stx) Classification, Structure, and Function. Microbiol. Spectr. 2014, 2, EHEC-0024-2013. [Google Scholar]
- Ethelberg, S.; Olsen, K.E.P.; Scheutz, F.; Jensen, C.; Schiellerup, P.; Engberg, J.; Petersen, A.M.; Olesen, B.; Gerner-Smidt, P.; Mølbak, K. Virulence factors for hemolytic uremic syndrome, Denmark. Emerg. Infect. Dis. 2004, 10, 842–847. [Google Scholar] [CrossRef] [PubMed]
- Toma, C.; Espinosa, E.M.; Song, T.; Miliwebsky, E.; Chinen, I.; Iyoda, S.; Iwanaga, M.; Rivas, M. Distribution of putative adhesins in different seropathotypes of Shiga toxin-producing Escherichia coli. J. Clin. Microbiol. 2004, 42, 4937–4946. [Google Scholar] [CrossRef] [PubMed]
- Luna-Gierke, R.E.; PM Griffin LH Gould KHerman CA Bopp NStrockbine RK, M.o.d.y. Outbreaks of non-O157 Shiga toxin-producing Escherichia coli infection: USA. Epidemiol. Infect. 2014, 142, 2270–2280. [Google Scholar] [CrossRef] [PubMed]
- Fratamico, P.M.; Bhaduri, S.; Buchanan, R.L. Studies on Escherichia coli serotype O157:H7 strains containing a 60-MDa plasmid and on 60-MDa plasmid-cured derivatives. J. Med. Microbiol. 1993, 39, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Fratamico, P.M.; Yan, X.; Caprioli, A.; Esposito, G.; Needleman, D.S.; Pepe, T.; Tozzoli, R.; Cortesi, M.L.; Morabito, S. The complete DNA sequence and analysis of the virulence plasmid and of five additional plasmids carried by Shiga toxin-producing Escherichia coli O26:H11 strain H30. Int. J. Med. Microbiol. 2011, 301, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Idelevich, E.A.; Zhang, W.; Bauwens, A.; Schaumburg, F.; Mellmann, A.; Peters, G.; Karch, H. Effects of antibiotics on Shiga toxin 2 production and bacteriophage induction by epidemic Escherichia coli O104:H4 strain. Antimicrob. Agents Chemother. 2012, 56, 3277–3282. [Google Scholar] [CrossRef]
- Hendriksen, R.S.; Bortolaia, V.; Tate, H.; Tyson, G.H.; Aarestrup, F.M.; McDermott, P.F. Using Genomics to Track Global Antimicrobial Resistance. Front. Public Health 2019, 7, 242. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention (CDC). Reports of Selected E. coli Outbreak Investigations. E. coli. CDC n.d. Available online: https://www.cdc.gov/ecoli/outbreaks.html (accessed on 10 May 2021).
- Smith, K.E.; Wilker, P.R.; Reiter, P.L.; Hedican, E.B.; Bender, J.B.; Hedberg, C.W. Antibiotic treatment of Escherichia coli O157 infection and the risk of hemolytic uremic syndrome, Minnesota. Pediatr. Infect. Dis. J. 2012, 31, 37–41. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Karmali, M.A.; Mascarenhas, M.; Shen, S.; Ziebell, K.; Johnson, S.; Reid-Smith, R.; Isaac-Renton, J.; Clark, C.; Rahn, K.; Kaper, J.B. Association of genomic O island 122 of Escherichia coli EDL 933 with verocytotoxin-producing Escherichia coli seropathotypes that are linked to epidemic and/or serious disease. J. Clin. Microbiol. 2003, 41, 4930–4940. [Google Scholar] [CrossRef]
- Coombes, B.K.; Wickham, M.E.; Mascarenhas, M.; Gruenheid, S.; Finlay, B.B.; Karmali, M.A. Molecular analysis as an aid to assess the public health risk of non-O157 Shiga toxin-producing Escherichia coli strains. Appl. Environ. Microbiol. 2008, 74, 2153–2160. [Google Scholar] [CrossRef]
- Karch, H.; Schubert, S.; Zhang, D.; Zhang, W.; Schmidt, H.; Olschlager, T.; Hacker, J. A genomic island, termed high-pathogenicity island, is present in certain non-O157 Shiga toxin-producing Escherichia coli clonal lineages. Infect. Immun. 1999, 67, 5994–6001. [Google Scholar] [CrossRef] [PubMed]
- Gerrish, R.S.; Lee, J.E.; Reed, J.; Williams, J.; Farrell, L.D.; Spiegel, K.M.; Sheridan, P.P.; Shields, M.S. PCR versus hybridization for detecting virulence genes of enterohemorrhagic Escherichia coli. Emerg. Infect. Dis. 2007, 13, 1253–1255. [Google Scholar] [CrossRef] [PubMed]
- Ponce, E.; Khan, A.A.; Cheng, C.-M.; Summage-West, C.; Cerniglia, C.E. Prevalence and characterization of Salmonella enterica serovar Weltevreden from imported seafood. Food Microbiol. 2008, 25, 29–35. [Google Scholar] [PubMed]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar]
- Levine, M.M.; McEwen, J.; Losonsky, G.; Reymann, M.; Harari, I.; Brown, J.E.; Taylor, D.N.; Donohue-Rolfe, A.; Cohen, D.; Bennish, M. Antibodies to shiga holotoxin and to two synthetic peptides of the B subunit in sera of patients with Shigella dysenteriae 1 dysentery. J. Clin. Microbiol. 1992, 30, 1636–1641. [Google Scholar] [CrossRef]
- Tarr, P.I.; Gordon, C.A.; Chandler, W.L. Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet 2005, 365, 1073–1086. [Google Scholar]
- Karmali, M.A. Prospects for preventing serious systemic toxemic complications of Shiga toxin-producing Escherichia coli infections using Shiga toxin receptor analogues. J. Infect. Dis. 2004, 189, 355–359. [Google Scholar] [CrossRef]
- Murinda, S.E.; Ibekwe, A.M.; Rodriguez, N.G.; Quiroz, K.L.; Mujica, A.P.; Osmon, K. Shiga Toxin-Producing Escherichia coli in Mastitis: An International Perspective. Foodborne Pathog. Dis. 2019, 16, 229–243. [Google Scholar]
- Moyo, S.J.; Maselle, S.Y.; Matee, M.I.; Langeland, N.; Mylvaganam, H. Identification of diarrheagenic Escherichia coli isolated from infants and children in Dar es Salaam, Tanzania. BMC Infect. Dis. 2007, 7, 92. [Google Scholar]
- Momtaz, H.; Dehkordi, F.S.; Hosseini, M.J.; Sarshar, M.; Heidari, M. Serogroups, virulence genes and antibiotic resistance in Shiga toxin-producing Escherichia coli isolated from diarrheic and non-diarrheic pediatric patients in Iran. Gut Pathog. 2013, 5, 39. [Google Scholar] [CrossRef]
- Ogden, I.D.; MacRae, M.; Strachan, N.J. Concentration and prevalence of Escherichia coli O157 in sheep faeces at pasture in Scotland. J. Appl. Microbiol. 2005, 98, 646–651. [Google Scholar] [PubMed]
- Hamed, O.M.; Sabry, M.A.; Hassanain, N.A.; Hamza, E.; Hegazi, A.G.; Salman, M.B. Occurrence of virulent and antibiotic-resistant Shiga toxin-producing Escherichia coli in some food products and human stool in Egypt. Vet. World 2017, 10, 1233–1240. [Google Scholar] [PubMed]
- Piérard, D.; Van Damme, L.; Moriau, L.; Stevens, D.; Lauwers, S. Isolation and virulence factors of verocytotoxin-producing Escherichia coli in human stool samples. Clin. Microbiol. Infect. 1997, 3, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Johura, F.-T.; Parveen, R.; Islam, A.; Sadique, A.; Rahim, N.; Monira, S.; Khan, A.R.; Ahsan, S.; Ohnishi, M.; Watanabe, H.; et al. Occurrence of Hybrid Escherichia coli Strains Carrying Shiga Toxin and Heat-Stable Toxin in Livestock of Bangladesh. Front. Public. Health 2016, 4, 287. [Google Scholar]
- Gupta, M.D.; Sen, A.; Das, A. Occurrence of Escherichia coli carrying Shiga toxin-producing genes in buffaloes on smallholdings in Bangladesh. Vet. World 2018, 11, 1454–1458. [Google Scholar]
- Fan, R.; Shao, K.; Yang, X.; Bai, X.; Fu, S.; Sun, H.; Xu, Y.; Wang, H.; Li, Q.; Hu, B.; et al. High prevalence of non-O157 Shiga toxin-producing Escherichia coli in beef cattle detected by combining four selective agars. BMC Microbiol. 2019, 19, 213. [Google Scholar]
- Vaz, T.M.I.; Irino, K.; Nishimura, L.S.; Cergole-Novella, M.C.; Guth, B.E.C. Genetic heterogeneity of Shiga toxin-producing Escherichia coli strains isolated in Sao Paulo, Brazil, from 1976 through 2003, as revealed by pulsed-field gel electrophoresis. J. Clin. Microbiol. 2006, 44, 798–804. [Google Scholar] [CrossRef]
- Fratamico, P.M.; Bagi, L.K.; Bush, E.J.; Solow, B.T. Prevalence and characterization of shiga toxin-producing Escherichia coli in swine feces recovered in the National Animal Health Monitoring System’s Swine 2000 study. Appl. Environ. Microbiol. 2004, 70, 7173–7178. [Google Scholar]
- Tseng, M.; Fratamico, P.M.; Bagi, L.; Delannoy, S.; Fach, P.; Manning, S.D.; Funk, J.A. Diverse virulence gene content of Shiga toxin-producing Escherichia coli from finishing swine. Appl. Environ. Microbiol. 2014, 80, 6395–6402. [Google Scholar]
- Karch, H.; Bielaszewska, M. Sorbitol-fermenting Shiga toxin-producing Escherichia coli O157:H− strains: Epidemiology, phenotypic and molecular characteristics, and microbiological diagnosis. J. Clin. Microbiol. 2001, 39, 2043–2049. [Google Scholar]
- Tseng, M.; Fratamico, P.M.; Manning, S.D.; Funk, J.A. Shiga toxin-producing Escherichia coli in swine: The public health perspective. Anim. Health Res. Rev. 2014, 15, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Ju, W.; Shen, J.; Toro, M.; Zhao, S.; Meng, J. Distribution of pathogenicity islands OI-122, OI-43/48, and OI-57 and a high-pathogenicity island in Shiga toxin-producing Escherichia coli. Appl. Environ. Microbiol. 2013, 79, 3406–3412. [Google Scholar] [CrossRef] [PubMed]
- Schubert, S.; Picard, B.; Gouriou, S.; Heesemann, J.; Denamur, E. Yersinia high-pathogenicity island contributes to virulence in Escherichia coli causing extraintestinal infections. Infect. Immun. 2002, 70, 5335–5337. [Google Scholar] [CrossRef] [PubMed]
- Nishio, M.; Okada, N.; Miki, T.; Haneda, T.; Danbara, H. Identification of the outer-membrane protein PagC required for the serum resistance phenotype in Salmonella enterica serovar Choleraesuis. Microbiology 2005, 151, 863–873. [Google Scholar] [CrossRef]
- Schmidt, H.; Hensel, M. Pathogenicity islands in bacterial pathogenesis. Clin. Microbiol. Rev. 2004, 17, 14–56. [Google Scholar]
- Cadona, J.S.; Burgán, J.; González, J.; Bustamante, A.V.; Sanso, A.M. Differential expression of the virulence gene nleB among Shiga toxin-producing Escherichia coli strains. Heliyon 2020, 6, e04277. [Google Scholar] [CrossRef]
- Regua-Mangia, A.H.; Irino, K.; Pacheco, R.d.S.; Bezerra, R.M.P.; Périssé, A.R.S.; Teixeira, L.M. Molecular characterization of uropathogenic and diarrheagenic Escherichia coli pathotypes. J. Basic. Microbiol. 2010, 50 (Suppl. S1), S107–S115. [Google Scholar] [CrossRef]
- Tatsuno, I.; Horie, M.; Abe, H.; Miki, T.; Makino, K.; Shinagawa, H.; Taguchi, H.; Kamiya, S.; Hayashi, T.; Sasakawa, C. toxB gene on pO157 of enterohemorrhagic Escherichia coli O157:H7 is required for full epithelial cell adherence phenotype. Infect. Immun. 2001, 69, 6660–6669. [Google Scholar] [CrossRef]
- Nicholls, L.; Grant, T.H.; Robins-Browne, R.M. Identification of a novel genetic locus that is required for in vitro adhesion of a clinical isolate of enterohaemorrhagic Escherichia coli to epithelial cells. Mol. Microbiol. 2000, 35, 275–288. [Google Scholar] [CrossRef]
- Klapproth, J.-M.A.; Scaletsky, I.C.A.; McNamara, B.P.; Lai, L.-C.; Malstrom, C.; James, S.P.; Donnenberg, M.S. A large toxin from pathogenic Escherichia coli strains that inhibits lymphocyte activation. Infect. Immun. 2000, 68, 2148–2155. [Google Scholar]
- Johnson, T.J.; Nolan, L.K. Pathogenomics of the virulence plasmids of Escherichia coli. Microbiol. Mol. Biol. Rev. 2009, 73, 750–774. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Miriagou, V.; Bertini, A.; Loli, A.; Colinon, C.; Villa, L.; Whichard, J.M.; Rossolini, G.M. Replicon typing of plasmids encoding resistance to newer beta-lactams. Emerg. Infect. Dis. 2006, 12, 1145–1148. [Google Scholar] [CrossRef]
- Marcadé, G.; Deschamps, C.; Boyd, A.; Gautier, V.; Picard, B.; Branger, C.; Denamur, E.; Arlet, G. Replicon typing of plasmids in Escherichia coli producing extended-spectrum beta-lactamases. J. Antimicrob. Chemother. 2009, 63, 67–71. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene | Sequence of Nucleotides | Target Gene | Size (bp) |
|---|---|---|---|
| eae | F: ACGTTGCAGCATGGGTAACTC R: GATCGGCAACAGTTTCACCTG | eae | (818) |
| hlyA | F: GGTGCAGCAGAAAAAGTTGTAG R: TCTCGCCTGATAGTGTTTGGTA | hlyA | (1551) |
| stx1 | F: TCTCAGTGGGCGTTCTTATG R: TACCCCCTCAACTGCTAATA | stx1 | (338) |
| stx2 | F: GCGGTTTTATTTGCATTAGC R: TCCCGTCAACCTTCACTGTA | stx2 | (115) |
| stx2f | F: TGTCTTCAGCATCTTATGCAG R: CATGATTAATTACTGAAACAGAAAC | stx2f | (150) |
| stx2c | F: GCGGTTTTATTTGCATTAGT R: AGTACTCTTTTCCGGCCACT | stx2c | (124) |
| stx2e | F: ATGAAGTGTATATTGTTAAAGTGGA R: AGCCACATATAAATTATTTCGT | stx2e | (303) |
| stx2d | F: GGTAAAATTGAGTTCTCTAAGTAT R: CAGCAAATCCTGAACCTGACG | stx2d | (175) |
| nel B | F: GGAAGTTTGTTTACAGAGACG R: AAAATGCCGCTTGATACC | nleB | (297) |
| pagC | F: ATGAGTGGTTCAAGACTGG R: CCAACTCCAACAGTAAATCC | pagC | (521) |
| sen | F: GGATGGAACCATACCTGG R: CGCAATCAATTGCTAATGC | sen | (551) |
| efa | F: CTCCCAGAGATAATTTTGAGG R: CAACTGTATGCGAATAGTACTC | efa1 | (504) |
| efa1 | F: CTGTCAGACGATGACATTGG R: GAAGGATGGGCATTGTGTC | efa1 | (547) |
| irp2 | R: GAAGGATGGGCATTGTGTC F: AAGGATTCGCTGTTACCGGAC | irp2 | (280) |
| toxB | F: TGGCCTTGCGCTCTATAAGAACCT R: ACCACGCCGTGAGAATAATGTCCA | toxB | (823) c |
| Replicon | Sequence of Nucleotides | Target Site | Size (bp) |
|---|---|---|---|
| HI1 | F-GGAGCGATGGATTACTTCAGTAC R-TGCCGTTTCACCTCGTGAGTA | parA-parB | 471 |
| HI2 | F-TTTCTCCTGAGTCACCTGTTAACAC R-GGCTCACTACCGTTGTCATCCT | iterons | 644 |
| I1 | F-CGAAAGCCGGACGGCAGAA R-TCGTCGTTCCGCCAAGTTCGT | RNAI | 139 |
| X | F-AACCTTAGAGGCTATTTAAGTTGCTGAT R-TGAGAGTCAATTTTTATCTCATGTTTTAGC | Ori γ | 376 |
| L/M | F-GGATGAAAACTATCAGCATCTGAAG R-CTGCAGGGGCGATTCTTTAGG | rep A, B, C | 785 |
| N | F-GTCTAACGAGCTTACCGAAG R-GTTTCAACTCTGCCAAGTTC | rep A | 559 |
| FIA | F-CCATGCTGGTTCTAGAGAAGGTG R-GTATATCCTTACTGGCTTCCGCAG | iterons | 462 |
| FIIB | F-GGAGTTCTGACACACGATTTTCTG R-CTCCCGTCGCTTCAGGGCATT | rep A | 702 |
| W | F-CCTAAGAACAACAAAGCCCCCG R-GGTGCGCGGCATAGAACCGT | rep A | 242 |
| FIC | F-GTGAACTGGCAGATGAGGAAGG R-TTCTCCTCGTCGCCAAACTAGAT | rep A2 | 262 |
| A/C | F-GAGAACCAAAGACAAAGACCTGGA R-ACGACAAACCTGAATTGCCTCCTT | rep A | 465 |
| T | F-TTGGCCTGTTTGTGCCTAAACCAT R-CGTTGATTACACTTAGCTTTGGAC | rep A | 750 |
| FIIA | F-CTGTCGTAAGCTGATGGC R-CTCTGCCACAAACTTCAGC | rep A | 270 |
| K/B | F-GCGGTCCGGAAAGCCAGAAAAC R-TCTTTCACGAGCCCGCCAAA | RNAI | 160 |
| B/O | F-GCGGTCCGGAAAGCCAGAAAAC R-TCTGCGTTCCGCCAAGTTCGA | RNAI | 159 |
| Y | F-AATTCAAACAACACTGTGCAGCCTG R-CGAGAATGGACGATTACAAAACTTT | repA | 765 |
| P | F-CTATGGCCCTGCAAACGCGCCAGAAA R-TCACGCGCCAGGGCGCAGCC | iterons | 534 |
| Rep | F-TGATCGTTTAAGGAATTTTG R-GAAGATCAGTCACACCATCC | RNAI/repA | 270 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alotaibi, K.; Khan, A.A. Prevalence and Molecular Characterization of Shiga Toxin-Producing Escherichia coli from Food and Clinical Samples. Pathogens 2023, 12, 1302. https://doi.org/10.3390/pathogens12111302
Alotaibi K, Khan AA. Prevalence and Molecular Characterization of Shiga Toxin-Producing Escherichia coli from Food and Clinical Samples. Pathogens. 2023; 12(11):1302. https://doi.org/10.3390/pathogens12111302
Chicago/Turabian StyleAlotaibi, Khulud, and Ashraf A. Khan. 2023. "Prevalence and Molecular Characterization of Shiga Toxin-Producing Escherichia coli from Food and Clinical Samples" Pathogens 12, no. 11: 1302. https://doi.org/10.3390/pathogens12111302
APA StyleAlotaibi, K., & Khan, A. A. (2023). Prevalence and Molecular Characterization of Shiga Toxin-Producing Escherichia coli from Food and Clinical Samples. Pathogens, 12(11), 1302. https://doi.org/10.3390/pathogens12111302