
Molecular Detection of Tick-Borne Bacterial and Protozoan Pathogens in Haemaphysalis longicornis (Acari: Ixodidae) Ticks from Free-Ranging Domestic Sheep in Hebei Province, China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
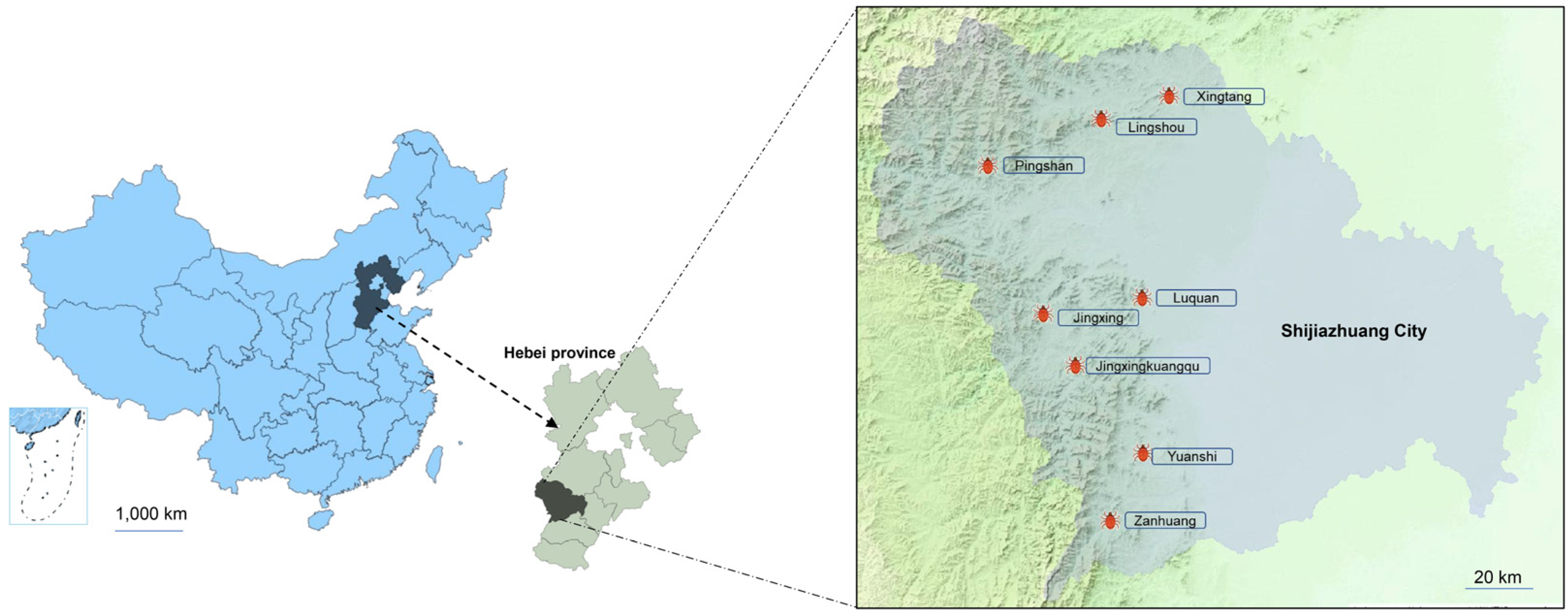
2.1. Study Area and Tick Sampling Protocol
2.2. DNA Extraction
2.3. Detection of Bacteria and Parasites in Ticks
2.4. Phylogenetic Data Analysis
2.5. Accession Numbers of Nucleotide Sequence
3. Results
3.1. Tick Sampling and Identification
3.2. Tick-Borne Pathogens Detected
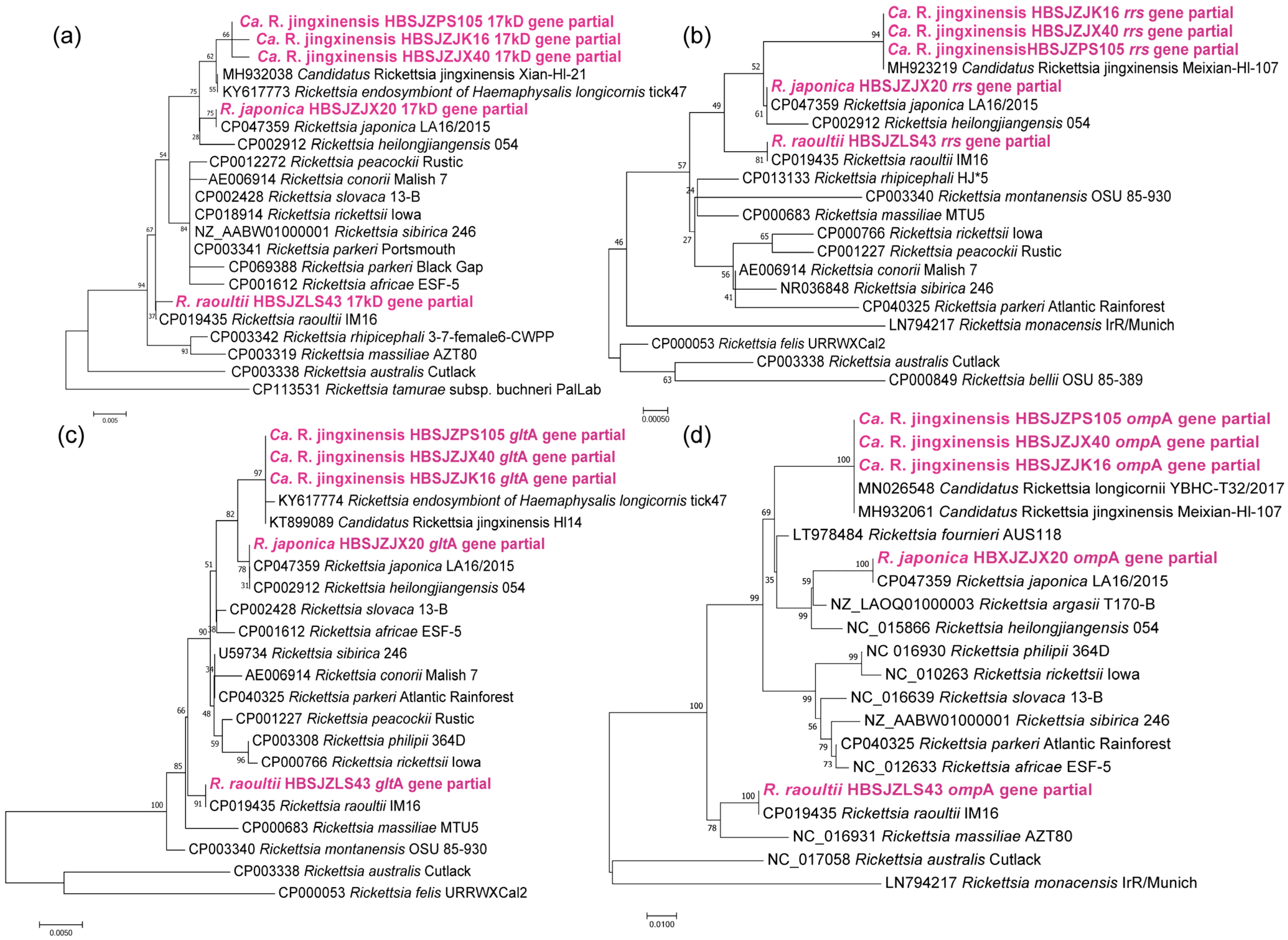
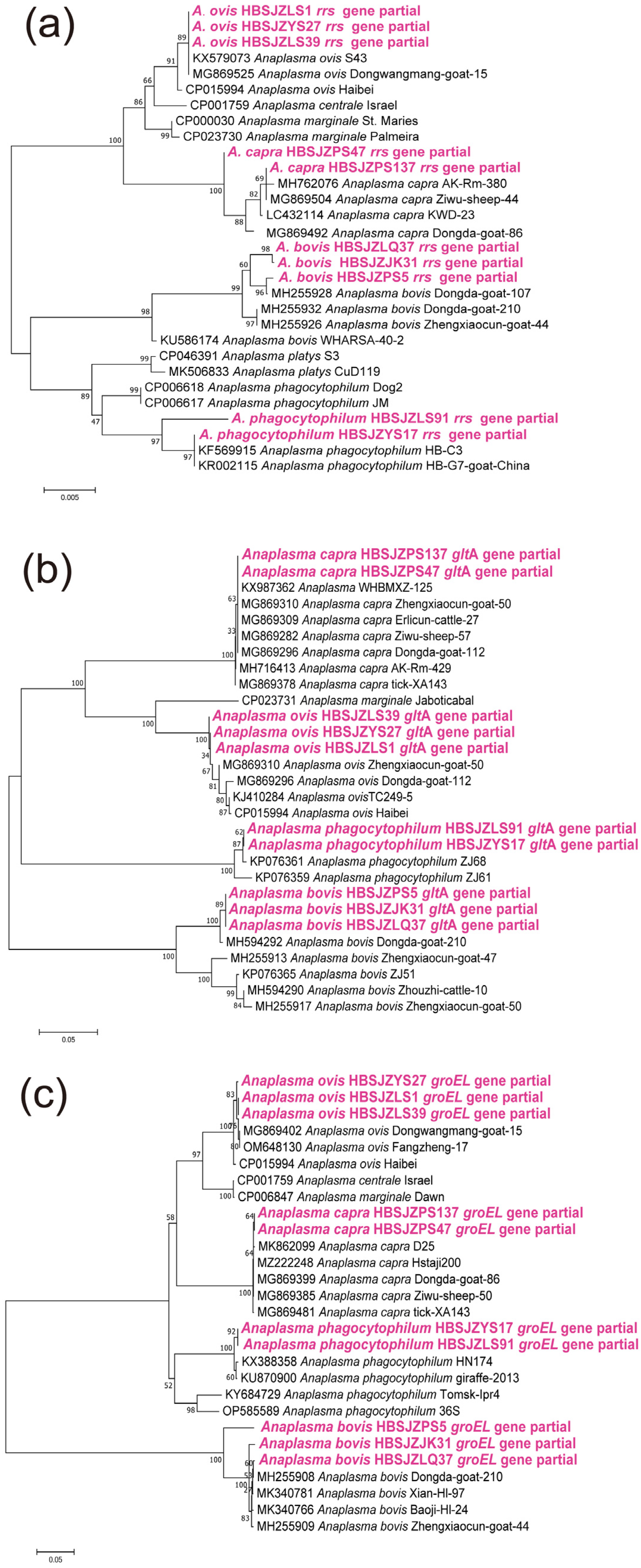
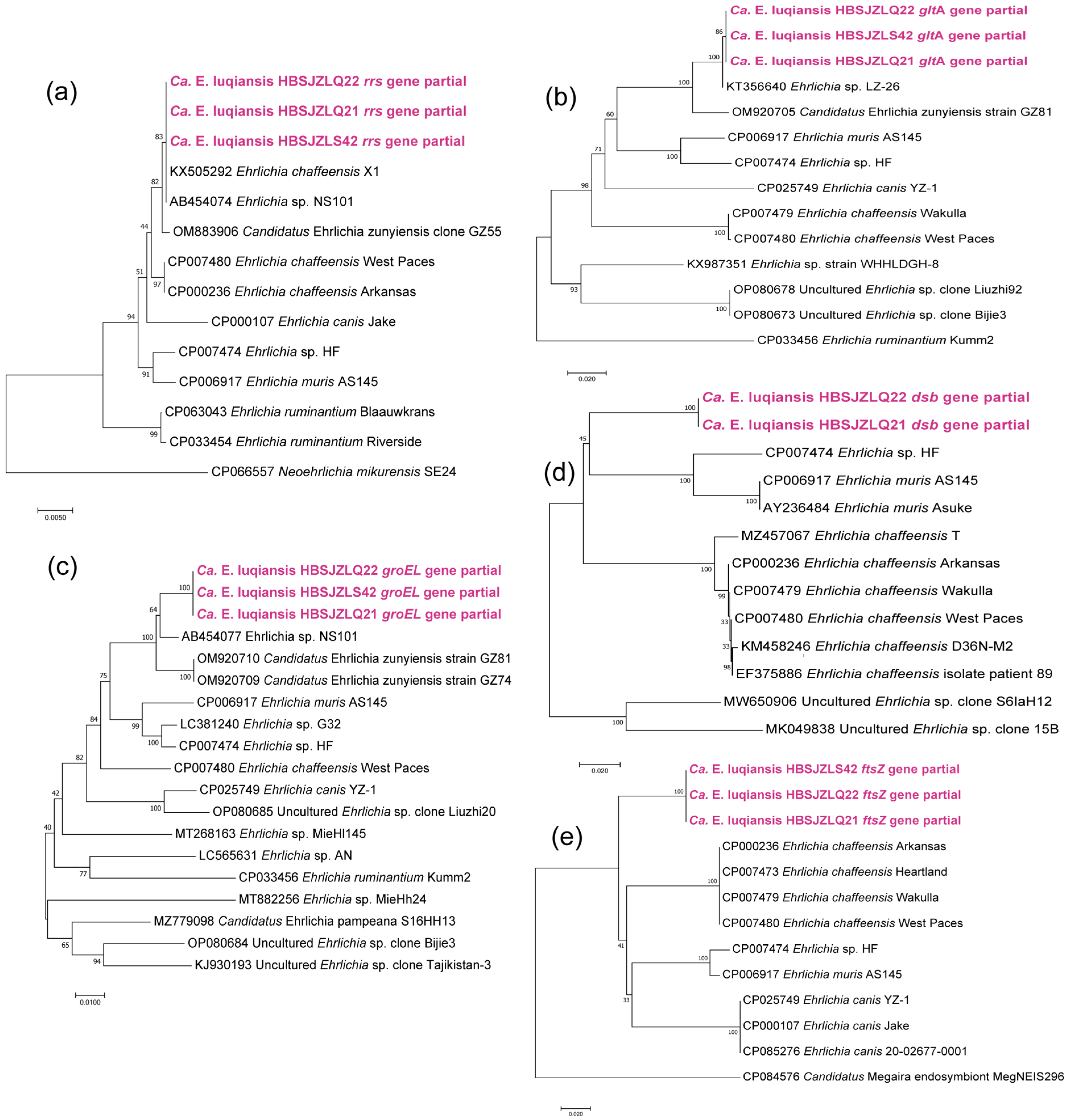
3.3. Phylogenic Analysis of Different Tick-Borne Pathogens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Estrada-Peña, A. Ticks as vectors: Taxonomy, biology and ecology. Rev. Sci. Et. Tech. 2015, 34, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, K.L.; Jizhou, L.; Phipps, L.P.; Johnson, N. Emerging Tick-Borne Viruses in the Twenty-First Century. Front. Cell Infect. Microbiol. 2017, 7, 298. [Google Scholar] [CrossRef] [PubMed]
- Teng, Z.; Zhao, N.; Ren, R.; Zhang, X.; Du, Z.; Wang, P.; Qin, T. Human Rickettsia felis infections in Mainland China. Front. Cell Infect. Microbiol. 2022, 12, 997315. [Google Scholar] [CrossRef] [PubMed]
- Farooq, I.; Moriarty, T.J. The Impact of Tick-Borne Diseases on the Bone. Microorganisms 2021, 9, 663. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; An, H.; Lee, J.S.; O’Guinn, M.L.; Kim, H.C.; Chong, S.T.; Zhang, Y.; Song, D.; Burrus, R.G.; Bao, Y.; et al. Molecular characterization of Haemaphysalis longicornis-borne rickettsiae, Republic of Korea and China. Ticks Tick-Borne Dis. 2018, 9, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.P.; Wang, Y.X.; Fan, Z.W.; Ji, Y.; Liu, M.J.; Zhang, W.H.; Li, X.L.; Zhou, S.X.; Li, H.; Liang, S.; et al. Mapping ticks and tick-borne pathogens in China. Nat. Commun. 2021, 12, 1075. [Google Scholar] [CrossRef]
- Price, K.J.; Witmier, B.J.; Eckert, R.A.; Boyer, C.N. Recovery of Partially Engorged Haemaphysalis longicornis (Acari: Ixodidae) Ticks from Active Surveillance. J. Med. Entomol. 2022, 59, 1842–1846. [Google Scholar] [CrossRef]
- Blisnick, A.A.; Foulon, T.; Bonnet, S.I. Serine Protease Inhibitors in Ticks: An Overview of Their Role in Tick Biology and Tick-Borne Pathogen Transmission. Front. Cell Infect. Microbiol. 2017, 7, 199. [Google Scholar] [CrossRef]
- Madison-Antenucci, S.; Kramer, L.D.; Gebhardt, L.L.; Kauffman, E. Emerging Tick-Borne Diseases. Clin. Microbiol. Rev. 2020, 33, e00083-18. [Google Scholar] [CrossRef]
- Rochlin, I.; Toledo, A. Emerging tick-borne pathogens of public health importance: A mini-review. J. Med. Microbiol. 2020, 69, 781–791. [Google Scholar] [CrossRef]
- Robinson, M.T.; Satjanadumrong, J.; Hughes, T.; Stenos, J.; Blacksell, S.D. Diagnosis of spotted fever group Rickettsia infections: The Asian perspective. Epidemiol. Infect. 2019, 147, e286. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.C.; Zhan, L.; De Vlas, S.J.; Wen, B.H.; Yang, H.; Richardus, J.H.; Habbema, J.D. Molecular detection of spotted fever group Rickettsia in Dermacentor silvarum from a forest area of northeastern China. J. Med. Entomol. 2008, 45, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Teng, Z.; Shi, Y.; Peng, Y.; Zhang, H.; Luo, X.; Lun, X.; Xia, L.; You, Y.; Li, Z.; Zhang, W.; et al. Severe Case of Rickettsiosis Identified by Metagenomic Sequencing, China. Emerg. Infect. Dis. 2021, 27, 1530–1532. [Google Scholar] [CrossRef]
- Qi, Y.; Gong, W.; Xiong, X.; Jiang, J.; Wang, Y.; Jiao, J.; Duan, C.; Wen, B. Microarray of surface-exposed proteins of Rickettsia heilongjiangensis for serodiagnosis of Far-eastern spotted fever. BMC Infect. Dis. 2014, 14, 332. [Google Scholar] [CrossRef] [PubMed]
- Teng, Z.; Gong, P.; Wang, W.; Zhao, N.; Jin, X.; Sun, X.; Zhou, H.; Lu, J.; Lin, X.; Wen, B.; et al. Clinical Forms of Japanese Spotted Fever from Case-Series Study, Zigui County, Hubei Province, China, 2021. Emerg. Infect. Dis. 2023, 29, 202–206. [Google Scholar] [CrossRef]
- Jia, N.; Zheng, Y.C.; Ma, L.; Huo, Q.B.; Ni, X.B.; Jiang, B.G.; Chu, Y.L.; Jiang, R.R.; Jiang, J.F.; Cao, W.C. Human infections with Rickettsia raoultii, China. Emerg. Infect. Dis. 2014, 20, 866–868. [Google Scholar] [CrossRef]
- Lu, M.; Li, F.; Liao, Y.; Shen, J.J.; Xu, J.M.; Chen, Y.Z.; Li, J.H.; Holmes, E.C.; Zhang, Y.Z. Epidemiology and Diversity of Rickettsiales Bacteria in Humans and Animals in Jiangsu and Jiangxi provinces, China. Sci. Rep. 2019, 9, 13176. [Google Scholar] [CrossRef]
- Liu, W.; Li, H.; Lu, Q.B.; Cui, N.; Yang, Z.D.; Hu, J.G.; Fan, Y.D.; Guo, C.T.; Li, X.K.; Wang, Y.W.; et al. Candidatus Rickettsia tarasevichiae Infection in Eastern Central China: A Case Series. Ann. Intern. Med. 2016, 164, 641–648. [Google Scholar] [CrossRef]
- Li, H.; Cui, X.M.; Cui, N.; Yang, Z.D.; Hu, J.G.; Fan, Y.D.; Fan, X.J.; Zhang, L.; Zhang, P.H.; Liu, W.; et al. Human Infection with Novel Spotted Fever Group Rickettsia Genotype, China, 2015. Emerg. Infect. Dis. 2016, 22, 2153–2156. [Google Scholar] [CrossRef]
- Li, H.; Li, X.M.; Du, J.; Zhang, X.A.; Cui, N.; Yang, Z.D.; Xue, X.J.; Zhang, P.H.; Cao, W.C.; Liu, W. Candidatus Rickettsia xinyangensis as Cause of Spotted Fever Group Rickettsiosis, Xinyang, China, 2015. Emerg. Infect. Dis. 2020, 26, 985–988. [Google Scholar] [CrossRef]
- Lu, M.; Chen, Q.; Qin, X.; Lyu, Y.; Teng, Z.; Li, K.; Yu, L.; Jin, X.; Chang, H.; Wang, W.; et al. Anaplasma bovis Infection in Fever and Thrombocytopenia Patients—Anhui Province, China, 2021. China CDC Wkly. 2022, 4, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zheng, Y.C.; Ma, L.; Jia, N.; Jiang, B.G.; Jiang, R.R.; Huo, Q.B.; Wang, Y.W.; Liu, H.B.; Chu, Y.L.; et al. Human infection with a novel tick-borne Anaplasma species in China: A surveillance study. Lancet. Infect. Dis. 2015, 15, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.J.; Dumler, J.S.; Carlyon, J.A. Current management of human granulocytic anaplasmosis, human monocytic ehrlichiosis and Ehrlichia ewingii ehrlichiosis. Expert. Rev. Anti-Infect. Ther. 2009, 7, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.K.; Schiffman, E.K.; Davis, J.P.; Neitzel, D.F.; Sloan, L.M.; Nicholson, W.L.; Fritsche, T.R.; Steward, C.R.; Ray, J.A.; Miller, T.K.; et al. Human Infection with Ehrlichia muris-like Pathogen, United States, 2007–2013(1). Emerg. Infect. Dis. 2015, 21, 1794–1799. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.R.; Strle, F.; Wormser, G.P. Comparison of Lyme Disease in the United States and Europe. Emerg. Infect. Dis. 2021, 27, 2017–2024. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.O.; Tolf, C.; Tamba, P.; Stefanache, M.; Radbea, G.; Rubel, F.; Waldenström, J.; Dobler, G.; Chițimia-Dobler, L. Babesia, Theileria, and Hepatozoon species in ticks infesting animal hosts in Romania. Parasitol. Res. 2017, 116, 2291–2297. [Google Scholar] [CrossRef]
- Soares, H.S.; Marcili, A.; Barbieri, A.R.M.; Minervino, A.H.H.; Moreira, T.R.; Gennari, S.M.; Labruna, M.B. Novel piroplasmid and Hepatozoon organisms infecting the wildlife of two regions of the Brazilian Amazon. Int. J. Parasitol. Parasites Wildl. 2017, 6, 115–121. [Google Scholar] [CrossRef]
- Tirloni, L.; Islam, M.S.; Kim, T.K.; Diedrich, J.K.; Yates, J.R., 3rd; Pinto, A.F.; Mulenga, A.; You, M.J.; Da Silva Vaz, I., Jr. Saliva from nymph and adult females of Haemaphysalis longicornis: A proteomic study. Parasites Vectors 2015, 8, 338. [Google Scholar] [CrossRef]
- Li, J.; Kelly, P.; Guo, W.; Zhang, J.; Yang, Y.; Liu, W.; Wang, C. Molecular detection of Rickettsia, Hepatozoon, Ehrlichia and SFTSV in goat ticks. Vet. Parasitol. Reg. Stud. Rep. 2020, 20, 100407. [Google Scholar] [CrossRef]
- Yao, J.; Xu, Z.; Sun, Z.; Zhou, K.; Lu, J.; Mi, R.; Huang, Y.; Han, X.; Ren, K.; Chen, Z.; et al. Response of Leucine-Rich Repeat Domain-Containing Protein in Haemaphysalis longicornis to Babesia microti Infection and Its Ligand Identification. Infect. Immun. 2021, 89, e00268-20. [Google Scholar] [CrossRef]
- Chen, Z.; Yang, X.; Bu, F.; Yang, X.; Yang, X.; Liu, J. Ticks (Acari: Ixodoidea: Argasidae, Ixodidae) of China. Exp. Appl. Acarol. 2010, 51, 393–404. [Google Scholar] [CrossRef]
- Lu, B.; Wu, H. Classification and Identification of Important Medical Insects in China; Henan Science and Technology Press: Zhengzhou, China, 2003. [Google Scholar]
- Kumar, B.; Maharana, B.R.; Thakre, B.; Brahmbhatt, N.N.; Joseph, J.P. 18S rRNA Gene-Based Piroplasmid PCR: An Assay for Rapid and Precise Molecular Screening of Theileria and Babesia Species in Animals. Acta Parasitol. 2022, 67, 1697–1707. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; He, B.; Gong, Z.; Liu, Y.; Wang, Q.; Yan, X.; Zhang, T.; Masoudi, A.; Zhang, X.; Wang, T.; et al. The new Haemaphysalis longicornis genome provides insights into its requisite biological traits. Genomics 2022, 114, 110317. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Li, J.; Cui, X.; Jia, N.; Wei, J.; Xia, L.; Wang, H.; Zhou, Y.; Wang, Q.; Liu, X.; et al. Distribution of Haemaphysalis longicornis and associated pathogens: Analysis of pooled data from a China field survey and global published data. Lancet. Planet. Health 2020, 4, e320–e329. [Google Scholar] [CrossRef] [PubMed]
- Gaywee, J.; Sunyakumthorn, P.; Rodkvamtook, W.; Ruang-areerate, T.; Mason, C.J.; Sirisopana, N. Human infection with Rickettsia sp. related to R. japonica, Thailand. Emerg. Infect. Dis. 2007, 13, 657–659. [Google Scholar] [CrossRef]
- Chung, M.H.; Lee, S.H.; Kim, M.J.; Lee, J.H.; Kim, E.S.; Lee, J.S.; Kim, M.K.; Park, M.Y.; Kang, J.S. Japanese spotted fever, South Korea. Emerg. Infect. Dis. 2006, 12, 1122–1124. [Google Scholar] [CrossRef]
- Li, J.; Hu, W.; Wu, T.; Li, H.B.; Hu, W.; Sun, Y.; Chen, Z.; Shi, Y.; Zong, J.; Latif, A.; et al. Japanese Spotted Fever in Eastern China, 2013. Emerg. Infect. Dis. 2018, 24, 2107–2109. [Google Scholar] [CrossRef]
- Lu, Q.; Yu, J.; Yu, L.; Zhang, Y.; Chen, Y.; Lin, M.; Fang, X. Rickettsia japonica Infections in Humans, Zhejiang Province, China, 2015. Emerg. Infect. Dis. 2018, 24, 2077–2079. [Google Scholar] [CrossRef]
- Li, H.; Zhang, P.H.; Du, J.; Yang, Z.D.; Cui, N.; Xing, B.; Zhang, X.A.; Liu, W. Rickettsia japonica Infections in Humans, Xinyang, China, 2014–2017. Emerg. Infect. Dis. 2019, 25, 1719–1722. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Sun, Y.Q.; Chen, J.J.; Teng, A.Y.; Wang, T.; Li, H.; Hay, S.I.; Fang, L.Q.; Yang, Y.; Liu, W. Mapping the global distribution of spotted fever group rickettsiae: A systematic review with modelling analysis. Lancet. Digit. Health 2023, 5, e5–e15. [Google Scholar] [CrossRef]
- Rydkina, E.; Roux, V.; Rudakov, N.; Gafarova, M.; Tarasevich, I.; Raoult, D. New Rickettsiae in ticks collected in territories of the former soviet union. Emerg. Infect. Dis. 1999, 5, 811–814. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, P.H.; Huang, Y.; Du, J.; Cui, N.; Yang, Z.D.; Tang, F.; Fu, F.X.; Li, X.M.; Cui, X.M.; et al. Isolation and Identification of Rickettsia raoultii in Human Cases: A Surveillance Study in 3 Medical Centers in China. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2018, 66, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, V.; Oteo, J.A.; Portillo, A.; Santibáñez, S.; Blanco, J.R.; Metola, L.; Eiros, J.M.; Pérez-Martínez, L.; Sanz, M. Rickettsia slovaca infection: DEBONEL/TIBOLA. Ann. N. Y. Acad. Sci. 2006, 1078, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Husin, N.A.; Khoo, J.J.; Zulkifli, M.M.S.; Bell-Sakyi, L.; AbuBakar, S. Replication Kinetics of Rickettsia raoultii in Tick Cell Lines. Microorganisms 2021, 9, 1370. [Google Scholar] [CrossRef] [PubMed]
- Igolkina, Y.; Rar, V.; Vysochina, N.; Ivanov, L.; Tikunov, A.; Pukhovskaya, N.; Epikhina, T.; Golovljova, I.; Tikunova, N. Genetic variability of Rickettsia spp. in Dermacentor and Haemaphysalis ticks from the Russian Far East. Ticks Tick-Borne Dis. 2018, 9, 1594–1603. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Guo, W.B.; Pan, Y.S.; Jiang, B.G.; Du, C.H.; Que, T.C.; Zhan, L.; Wu, J.H.; Yu, M.H.; Cui, X.M.; et al. Detection of Novel Spotted Fever Group Rickettsiae (Rickettsiales: Rickettsiaceae) in Ticks (Acari: Ixodidae) in Southwestern China. J. Med. Entomol. 2021, 58, 1363–1369. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Wang, Q.; Fu, Z.; Liu, P.; Jin, H.; Yang, H.; Gao, H.; Xi, Z.; Liu, Q.; Chen, L. Detection of spotted fever group Rickettsia in Haemaphysalis longicornis from Hebei Province, China. J. Parasitol. 2011, 97, 960–962. [Google Scholar] [CrossRef]
- Aktas, M.; Özübek, S. Anaplasma ovis genetic diversity detected by major surface protein 1a and its prevalence in small ruminants. Vet. Microbiol. 2018, 217, 13–17. [Google Scholar] [CrossRef]
- Fishbein, D.B.; Dawson, J.E.; Robinson, L.E. Human ehrlichiosis in the United States, 1985 to 1990. Ann. Intern. Med. 1994, 120, 736–743. [Google Scholar] [CrossRef]
- Chen, S.M.; Dumler, J.S.; Bakken, J.S.; Walker, D.H. Identification of a granulocytotropic Ehrlichia species as the etiologic agent of human disease. J. Clin. Microbiol. 1994, 32, 589–595. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, H.; Xu, B.; Zhang, Z.; Jin, Y.; Li, W.; Lu, Q.; Li, L.; Chang, L.; Zhang, X.; et al. Rural residents in China are at increased risk of exposure to tick-borne pathogens Anaplasma phagocytophilum and Ehrlichia chaffeensis. BioMed Res. Int. 2014, 2014, 313867. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Zhang, Z.; Moldenhauer, S.; Guo, Y.; Yu, Q.; Wang, L.; Chen, M. Detection of Borrelia burgdorferi from ticks (Acari) in Hebei Province, China. J. Med. Entomol. 1998, 35, 95–98. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Zhou, G.; Yao, G.; Ai, C. Morphological Observation of Haemaphysalis congicornis and Borrelia Burgdorferi in the Tick’s Bodyin Xishan Mountain in Beijing. Chin. J. Vector Biol. Control. 2002, 13, 282–285. [Google Scholar]
- Duron, O.; Noël, V.; McCoy, K.D.; Bonazzi, M.; Sidi-Boumedine, K.; Morel, O.; Vavre, F.; Zenner, L.; Jourdain, E.; Durand, P.; et al. The Recent Evolution of a Maternally-Inherited Endosymbiont of Ticks Led to the Emergence of the Q Fever Pathogen, Coxiella burnetii. PLoS Pathog. 2015, 11, e1004892. [Google Scholar] [CrossRef] [PubMed]
- Kawamori, F.; Shimazu, Y.; Sato, H.; Monma, N.; Ikegaya, A.; Yamamoto, S.; Fujita, H.; Morita, H.; Tamaki, Y.; Takamoto, N.; et al. Evaluation of Diagnostic Assay for Rickettsioses Using Duplex Real-Time PCR in Multiple Laboratories in Japan. Jpn. J. Infect. Dis. 2018, 71, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.P.; Tian, J.H.; Lin, X.D.; Ni, X.B.; Chen, X.P.; Liao, Y.; Yang, S.Y.; Dumler, J.S.; Holmes, E.C.; Zhang, Y.Z. Extensive genetic diversity of Rickettsiales bacteria in multiple mosquito species. Sci. Rep. 2016, 6, 38770. [Google Scholar] [CrossRef] [PubMed]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef]
- Roux, V.; Rydkina, E.; Eremeeva, M.; Raoult, D. Citrate synthase gene comparison, a new tool for phylogenetic analysis, and its application for the rickettsiae. Int. J. Syst. Bacteriol. 1997, 47, 252–261. [Google Scholar] [CrossRef]
- Ngnindji-Youdje, Y.; Diarra, A.Z.; Lontsi-Demano, M.; Tchuinkam, T.; Parola, P. Detection of Tick-Borne Pathogens in Ticks from Cattle in Western Highlands of Cameroon. Microorganisms 2022, 10, 1957. [Google Scholar] [CrossRef]
- Jafar Bekloo, A.; Ramzgouyan, M.R.; Shirian, S.; Faghihi, F.; Bakhshi, H.; Naseri, F.; Sedaghat, M.; Telmadarraiy, Z. Molecular Characterization and Phylogenetic Analysis of Anaplasma spp. and Ehrlichia spp. Isolated from Various Ticks in Southeastern and Northwestern Regions of Iran. Vector Borne Zoonotic Dis. 2018, 18, 252–257. [Google Scholar] [CrossRef]
- Guo, W.P.; Huang, B.; Zhao, Q.; Xu, G.; Liu, B.; Wang, Y.H.; Zhou, E.M. Human-pathogenic Anaplasma spp., and Rickettsia spp. in animals in Xi’an, China. PLoS Negl. Trop. Dis. 2018, 12, e0006916. [Google Scholar] [CrossRef] [PubMed]
- Liz, J.S.; Anderes, L.; Sumner, J.W.; Massung, R.F.; Gern, L.; Rutti, B.; Brossard, M. PCR Detection of Granulocytic Ehrlichiae in Ixodes ricinus Ticks and Wild Small Mammals in Western Switzerland. J. Clin. Microbiol. 2000, 38, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.P.; Wang, X.; Li, Y.N.; Xu, G.; Wang, Y.H.; Zhou, E.M. GroEL gene typing and genetic diversity of Anaplasma bovis in ticks in Shaanxi, China. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2019, 74, 103927. [Google Scholar] [CrossRef] [PubMed]
- Remesar, S.; Prieto, A.; García-Dios, D.; López-Lorenzo, G.; Martínez-Calabuig, N.; Díaz-Cao, J.M.; Panadero, R.; López, C.M.; Fernández, G.; Díez-Baños, P.; et al. Diversity of Anaplasma species and importance of mixed infections in roe deer from Spain. Transbound. Emerg. Dis. 2022, 69, e374–e385. [Google Scholar] [CrossRef]
- Lu, M.; Meng, C.; Gao, X.; Sun, Y.; Zhang, J.; Tang, G.; Li, Y.; Li, M.; Zhou, G.; Wang, W.; et al. Diversity of Rickettsiales in Rhipicephalus microplus Ticks Collected in Domestic Ruminants in Guizhou Province, China. Pathogens 2022, 11, 1108. [Google Scholar] [CrossRef]
- Drazenovich, N.; Foley, J.; Brown, R.N. Use of real-time quantitative PCR targeting the msp2 protein gene to identify cryptic Anaplasma phagocytophilum infections in wildlife and domestic animals. Vector Borne Zoonotic Dis. 2006, 6, 83–90. [Google Scholar] [CrossRef]
- Su, H.; Onoda, E.; Tai, H.; Fujita, H.; Sakabe, S.; Azuma, K.; Akachi, S.; Oishi, S.; Abe, F.; Ando, S.; et al. Diversity unearthed by the estimated molecular phylogeny and ecologically quantitative characteristics of uncultured Ehrlichia bacteria in Haemaphysalis ticks, Japan. Sci. Rep. 2021, 11, 687. [Google Scholar] [CrossRef]
- Almeida, A.P.; Souza, T.D.; Marcili, A.; Labruna, M.B. Novel Ehrlichia and Hepatozoon Agents Infecting the Crab-Eating Fox (Cerdocyon thous) in Southeastern Brazil. J. Med. Entomol. 2013, 50, 640–646. [Google Scholar] [CrossRef]
- Michelet, L.; Delannoy, S.; Devillers, E.; Umhang, G.; Aspan, A.; Juremalm, M.; Chirico, J.; van der Wal, F.J.; Sprong, H.; Boye Pihl, T.P.; et al. High-throughput screening of tick-borne pathogens in Europe. Front. Cell. Infect. Microbiol. 2014, 4, 103. [Google Scholar] [CrossRef]
- Diaz, M.H.; Bai, Y.; Malania, L.; Winchell, J.M.; Kosoy, M.Y. Development of a novel genus-specific real-time PCR assay for detection and differentiation of Bartonella species and genotypes. J. Clin. Microbiol. 2012, 50, 1645–1649. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen Species | Prevalence (%) * | |
|---|---|---|
| Rickettsia | 5.1% | |
| R. japonica | 2.0% | |
| R. raoultii | 0.9% | |
| Ca. R. jingxinensis | 2.2% | |
| Anaplasma | 15.9% | |
| A. bovis | 8.0% | |
| A. ovis, | 4.8% | |
| A. phagocytophilum | 1.5% | |
| A. capra | 1.5% | |
| Ehrlichia | 1.2% | |
| Ca. E. luquansis | 1.2% | |
| Borrelia | 0.15% | |
| B. burgdorferi | 0.15% | |
| Theileria | 17.9% | |
| T. luwenshuni | 17.9% | |
| Hepatozoon | 0.15% | |
| H. felis | 0.15% | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teng, Z.; Shi, Y.; Zhao, N.; Zhang, X.; Jin, X.; He, J.; Xu, B.; Qin, T. Molecular Detection of Tick-Borne Bacterial and Protozoan Pathogens in Haemaphysalis longicornis (Acari: Ixodidae) Ticks from Free-Ranging Domestic Sheep in Hebei Province, China. Pathogens 2023, 12, 763. https://doi.org/10.3390/pathogens12060763
Teng Z, Shi Y, Zhao N, Zhang X, Jin X, He J, Xu B, Qin T. Molecular Detection of Tick-Borne Bacterial and Protozoan Pathogens in Haemaphysalis longicornis (Acari: Ixodidae) Ticks from Free-Ranging Domestic Sheep in Hebei Province, China. Pathogens. 2023; 12(6):763. https://doi.org/10.3390/pathogens12060763
Chicago/Turabian StyleTeng, Zhongqiu, Yan Shi, Na Zhao, Xue Zhang, Xiaojing Jin, Jia He, Baohong Xu, and Tian Qin. 2023. "Molecular Detection of Tick-Borne Bacterial and Protozoan Pathogens in Haemaphysalis longicornis (Acari: Ixodidae) Ticks from Free-Ranging Domestic Sheep in Hebei Province, China" Pathogens 12, no. 6: 763. https://doi.org/10.3390/pathogens12060763
APA StyleTeng, Z., Shi, Y., Zhao, N., Zhang, X., Jin, X., He, J., Xu, B., & Qin, T. (2023). Molecular Detection of Tick-Borne Bacterial and Protozoan Pathogens in Haemaphysalis longicornis (Acari: Ixodidae) Ticks from Free-Ranging Domestic Sheep in Hebei Province, China. Pathogens, 12(6), 763. https://doi.org/10.3390/pathogens12060763