Physics Comes to the Aid of Medicine—Clinically-Relevant Microorganisms through the Eyes of Atomic Force Microscope

 ,
,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
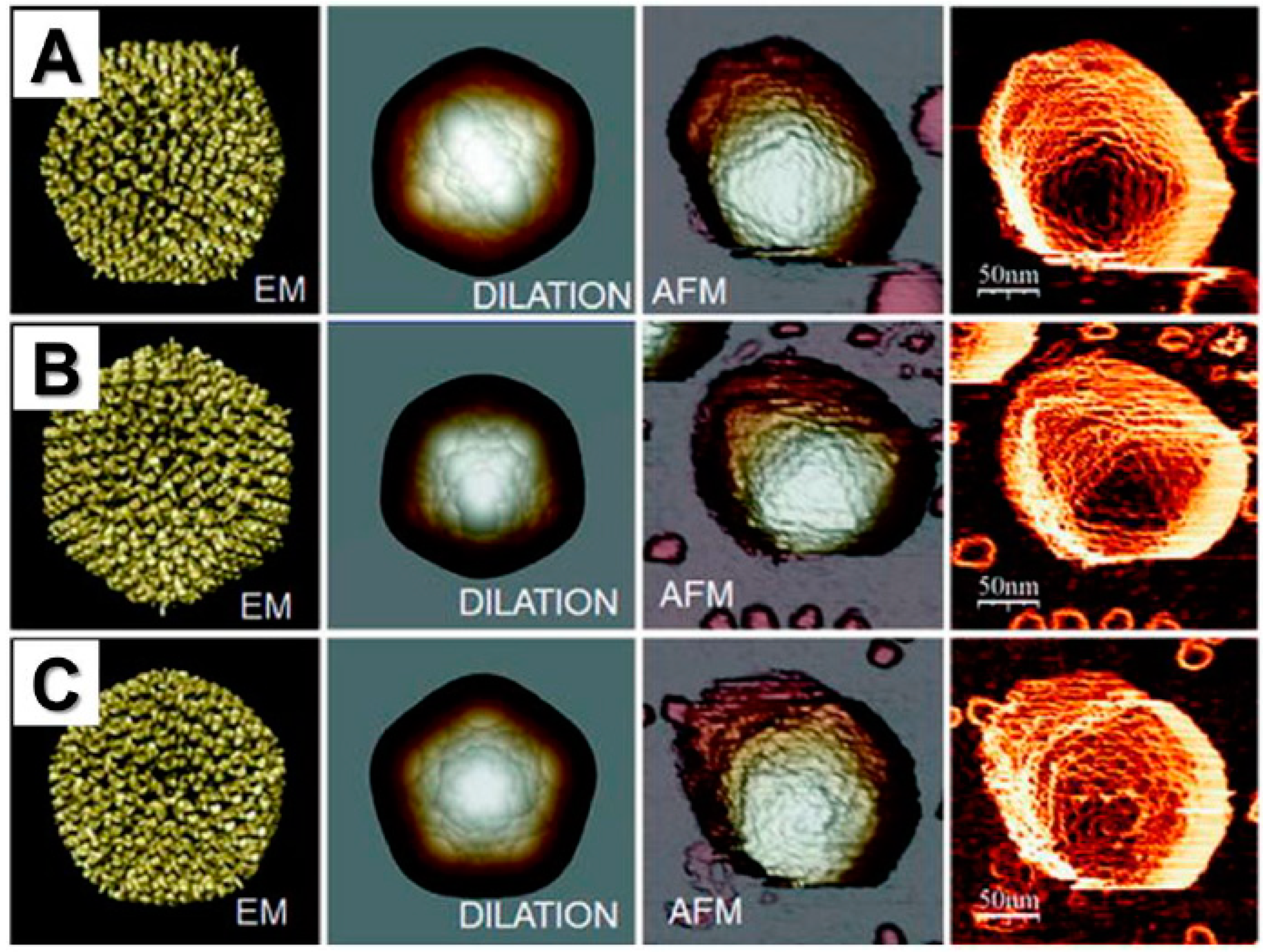{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
Application of Atomic Force Microscopy (AFM) in the Field of Microbiology
2. AFM-Based Investigation as a Novel Approach to Fight with Drug Resistance in Bacteria and Fungi
2.1. Resistance to Antibiotics—An Emerging Problem in Medical and Environmental Microbiology
2.2. Analysis of Nanotopography of Pathogens as an Approach to Elucidate Viability of Microbes and Antibiotics’ Mechanism of Action
2.2.1. Bacteria
2.2.2. Fungi
2.2.3. Viruses
2.3. Alterations in Microbes’ Cellular Stiffness as an Indicator of Antimicrobial Activity of Tested Molecules
2.4. AFM Analysis in Investigation of Microbial Surface Adhesion, Colonization Mechanisms, and Virus–Cell Binding
2.4.1. AFM Analysis of Mechanisms of Pathogens Colonization, Microbe–Microbe Interactions, and Binding to Cells
2.4.2. Utility of AFM in Fabrication of Biomaterials with Anti-Adhesive Properties
2.4.3. AFM Analysis of Material Surfaces for Industrial Purposes
2.4.4. The Employment of Atomic Force Microscopy in the Investigation of Microbial Biofilms
3. Investigation of Bacteria and Fungi Phenotypic and Virulence Features Using AFM
4. Physicochemical Characterization of Developed Antimicrobials Using AFM
5. AFM as Novel Tool to Improve Currently Used Diagnostic Methods
6. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tavenier, A.H.; de Boer, F.J.; Moshaver, B.; van der Leur, S.J.C.M.; Stegeman, C.A.; Groeneveld, P.H.P. Flow cytometric analysis of viable bacteria in urine samples of febrile patients at the emergency department. Cytom. B Clin. Cytom. 2017. [Google Scholar] [CrossRef] [PubMed]
- Durnaś, B.; Piktel, E.; Wątek, M.; Wollny, T.; Góźdź, S.; Smok-Kalwat, J.; Niemirowicz, K.; Savage, P.B.; Bucki, R. Anaerobic bacteria growth in the presence of cathelicidin LL-37 and selected ceragenins delivered as magnetic nanoparticles cargo. BMC Microbiol. 2017, 17, 167. [Google Scholar] [CrossRef] [PubMed]
- Niemirowicz, K.; Durnaś, B.; Tokajuk, G.; Piktel, E.; Michalak, G.; Gu, X.; Kułakowska, A.; Savage, P.B.; Bucki, R. Formulation and candidacidal activity of magnetic nanoparticles coated with cathelicidin LL-37 and ceragenin CSA-13. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Durnaś, B.; Wnorowska, U.; Pogoda, K.; Deptuła, P.; Wątek, M.; Piktel, E.; Głuszek, S.; Gu, X.; Savage, P.B.; Niemirowicz, K.; et al. Candidacidal Activity of Selected Ceragenins and Human Cathelicidin LL-37 in Experimental Settings Mimicking Infection Sites. PLoS ONE 2016, 11, e0157242. [Google Scholar] [CrossRef] [PubMed]
- Piktel, E.; Pogoda, K.; Roman, M.; Niemirowicz, K.; Tokajuk, G.; Wróblewska, M.; Szynaka, B.; Kwiatek, W.M.; Savage, P.B.; Bucki, R. Sporicidal activity of ceragenin CSA-13 against Bacillus subtilis. Sci. Rep. 2017, 7, 44452. [Google Scholar] [CrossRef] [PubMed]
- Hansma, P.K.; Elings, V.B.; Marti, O.; Bracker, C.E. Scanning tunneling microscopy and atomic force microscopy: Application to biology and technology. Science 1988, 242, 209–216. [Google Scholar] [CrossRef]
- Binnig, G.; Quate, C.F.; Gerber, C. Atomic force microscope. Phys. Rev. Lett. 1986, 56, 930–933. [Google Scholar] [CrossRef] [Green Version]
- Gould, S.A.C.; Drake, B.; Prater, C.B.; Weisenhorn, A.L.; Manne, S.; Kelderman, G.L.; Butt, H.J.; Hansma, H.; Hansma, P.K.; Magonov, S.; et al. The atomic force microscope: A tool for science and industry. Ultramicroscopy 1990, 33, 93–98. [Google Scholar] [CrossRef]
- Variola, F.; Vetrone, F.; Richert, L.; Jedrzejowski, P.; Yi, J.H.; Zalzal, S.; Clair, S.; Sarkissian, A.; Perepichka, D.F.; Wuest, J.D.; et al. Improving biocompatibility of implantable metals by nanoscale modification of surfaces: An overview of strategies, fabrication methods, and challenges. Small 2009, 5, 996–1006. [Google Scholar] [CrossRef]
- Rettler, E.; Hoeppener, S.; Sigusch, B.W.; Schubert, U.S. Mapping the mechanical properties of biomaterials on different length scales: Depth-sensing indentation and AFM based nanoindentation. J. Mater. Chem. B 2013, 1, 2789–2806. [Google Scholar] [CrossRef]
- Wang, D.; Nakajima, K.; Liu, F.; Shi, S.; Russell, T.P. Nanomechanical Imaging of the Diffusion of Fullerene into Conjugated Polymer. ACS Nano 2017. [Google Scholar] [CrossRef] [PubMed]
- Eaton, P.; West, P. Atomic Force Microscopy; Oxford Scholarship Online: Oxford, UK, 2010. [Google Scholar]
- Toumey, C. Probing the history of nanotechnology. Nat. Nanotechnol. 2012, 7, 205–206. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Tamm, L.K.; Somlyo, A.P.; Shao, Z. Promises and problems of biological atomic force microscopy. J. Microsc. 1993, 171, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Coleman, A.W.; Lazar, A.N.; Rousseau, C.F.; Cecillon, S.; Shahgaldian, P. Nanosystem Characterization Tools in the Life Sciences; Kumar, C.S.S.R., Ed.; Wiley-VCH: Hoboken, NJ, USA, 2011. [Google Scholar]
- Toca-Herrera, J.L. Atomic Force Microscopy Meets Biophysics, Bioengineering, Chemistry, and Materials Science. ChemSusChem 2019, 12, 603–611. [Google Scholar] [CrossRef] [Green Version]
- Pogoda, K.; Piktel, E.; Deptula, P.; Savage, P.B.; Lekka, M.; Bucki, R. Stiffening of bacteria cells as a first manifestation of bactericidal attack. Micron 2017, 101, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Delguste, M.; Koehler, M.; Alsteens, D. Probing Single Virus Binding Sites on Living Mammalian Cells Using AFM. Methods Mol. Biol. 2018, 1814, 483–514. [Google Scholar] [CrossRef]
- Pleshakova, T.O.; Kaysheva, A.L.; Bayzyanova, J.; Anashkina, A.; Uchaikin, V.F.; Ziborov, V.S.; Konev, V.A.; Archakov, A.I.; Ivanov, Y.D. The detection of hepatitis c virus core antigen using afm chips with immobolized aptamers. J. Virol. Methods 2018, 251, 99–105. [Google Scholar] [CrossRef]
- Piktel, E.; Wnorowska, U.; Ciesluk, M.; Deptula, P.; Pogoda, K.; Misztalewska-Turkowicz, I.; Paprocka, P.; Niemirowicz-Laskowska, K.; Wilczewska, A.Z.; Janmey, P.A.; et al. Inhibition of inflammatory response in human keratinocytes by magnetic nanoparticles functionalized with PBP10 peptide derived from the PIP2-binding site of human plasma gelsolin. J. Nanobiotechnol. 2019, 17, 22. [Google Scholar] [CrossRef]
- Del Valle, A.; Torra, J.; Bondia, P.; Tone, C.M.; Pedraz, P.; Vadillo-Rodriguez, V.; Flors, C. Mechanically Induced Bacterial Death Imaged in Real Time: A Simultaneous Nanoindentation and Fluorescence Microscopy Study. ACS Appl. Mater. Interfaces 2020, 12, 31235–31241. [Google Scholar] [CrossRef]
- Lonergan, N.E.; Britt, L.D.; Sullivan, C.J. Immobilizing live Escherichia coli for AFM studies of surface dynamics. Ultramicroscopy 2014, 137, 30–39. [Google Scholar] [CrossRef]
- Gammoudi, I.; Mathelie-Guinlet, M.; Morote, F.; Beven, L.; Moynet, D.; Grauby-Heywang, C.; Cohen-Bouhacina, T. Morphological and nanostructural surface changes in Escherichia coli over time, monitored by atomic force microscopy. Colloids Surf. B Biointerfaces 2016, 141, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wnorowska, U.; Niemirowicz, K.; Myint, M.; Diamond, S.L.; Wróblewska, M.; Savage, P.B.; Janmey, P.A.; Bucki, R. Bactericidal activities of cathelicidin LL-37 and select cationic lipids against the hypervirulent Pseudomonas aeruginosa strain LESB58. Antimicrob. Agents Chemother. 2015, 59, 3808–3815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Deering, A.J.; Bhunia, A.K.; Yao, Y. Biofilm of Escherichia coli O157:H7 on cantaloupe surface is resistant to lauroyl arginate ethyl and sodium hypochlorite. Int. J. Food Microbiol. 2017, 260, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Anselme, K.; Davidson, P.; Popa, A.M.; Giazzon, M.; Liley, M.; Ploux, L. The interaction of cells and bacteria with surfaces structured at the nanometre scale. Acta Biomater. 2010, 6, 3824–3846. [Google Scholar] [CrossRef]
- Biggs, M.J.; Richards, R.G.; Dalby, M.J. Nanotopographical modification: A regulator of cellular function through focal adhesions. Nanomedicine 2010, 6, 619–633. [Google Scholar] [CrossRef] [Green Version]
- Meincken, M.; Holroyd, D.L.; Rautenbach, M. Atomic force microscopy study of the effect of antimicrobial peptides on the cell envelope of Escherichia coli. Antimicrob. Agents Chemother. 2005, 49, 4085–4092. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Lee, P.Y.; Ho, B.; Ding, J.L.; Lim, C.T. Atomic force microscopy study of the antimicrobial action of Sushi peptides on Gram negative bacteria. Biochim. Biophys. Acta 2007, 1768, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Beaussart, A.; Abellán-Flos, M.; El-Kirat-Chatel, S.; Vincent, S.P.; Dufrêne, Y.F. Force Nanoscopy as a Versatile Platform for Quantifying the Activity of Antiadhesion Compounds Targeting Bacterial Pathogens. Nano Lett. 2016, 16, 1299–1307. [Google Scholar] [CrossRef]
- Canale, C.; Torre, B.; Ricci, D.; Braga, P.C. Recognizing and avoiding artifacts in atomic force microscopy imaging. Methods Mol. Biol. 2011, 736, 31–43. [Google Scholar] [CrossRef]
- Voigtländer, B. Artifacts in AFM. In Atomic Force Microscopy; Springer: Berlin, Germany, 2019; pp. 137–147. [Google Scholar]
- Petrov, L.; Matija, L. Atomic force microscopy as a tool for testing biomedical samples and elimination probe artifacts. Contem. Mater. 2019, X-2, 137–144. [Google Scholar] [CrossRef]
- Moon, D.; Jeon, B.R.; Kim, S. Thermal drift induced artifacts in AFM atomic lattice images. In Proceedings of the APS March Meeting 2020, Denver, CO, USA, 2–6 March 2020. [Google Scholar]
- Bobrov, Y.; Bykov, V.; Leesment, S.; Polyakov, V. Automation of topography and phase contrast measurements in tapping mode. In Proceedings of the Scanning Probe Microscopy, Russia-China Workshop on Dielectric and Ferroelectric Materials, Ekaterinburg, Russia, 25–28 August 2019; pp. 69–70. [Google Scholar]
- Zhavnerko, G.; Nikolaevich Poleschuyk, N. Mycobacterium under AFM tip: Advantages of polyelectrolyte modified substrate. Int. J. Mycobacteriol. 2012, 1, 53–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, X.; Walter, J.; Burke, S.; Stewart, S.; Jericho, M.H.; Pink, D.; Hunter, R.; Beveridge, T.J. Atomic force microscopy and theoretical considerations of surface properties and turgor pressures of bacteria. Colloids Surf. B Biointerfaces 2002, 23, 213–230. [Google Scholar] [CrossRef]
- Benn, G.; Pyne, A.L.B.; Ryadnov, M.G.; Hoogenboom, B.W. Imaging live bacteria at the nanoscale: Comparison of immobilisation strategies. Analyst 2019, 144, 6944–6952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allison, D.P.; Sullivan, C.J.; Mortensen, N.P.; Retterer, S.T.; Doktycz, M. Bacterial immobilization for imaging by atomic force microscopy. J. Vis. Exp. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasas, S.; Ikai, A. A method for anchoring round shaped cells for atomic force microscope imaging. Biophys. J. 1995, 68, 1678–1680. [Google Scholar] [CrossRef] [Green Version]
- Touhami, A.; Jericho, M.H.; Beveridge, T.J. Atomic force microscopy of cell growth and division in Staphylococcus aureus. J. Bacteriol. 2004, 186, 3286–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufrêne, Y.F. Atomic force microscopy and chemical force microscopy of microbial cells. Nat. Protoc. 2008, 3, 1132–1138. [Google Scholar] [CrossRef]
- Alzahrani, K.; Shukla, A.K.; Alam, J.; Niazy, A.A.; Alsouwaileh, A.M.; Alhoshan, M.; Khalid, J.; Alghamadi, H.S. Probing the surface ultrastructure of Brevibacillus laterosporus using atomic force microscopy. Micron 2020, 131, 102827. [Google Scholar] [CrossRef]
- Alam, F.; Balani, K. Adhesion force of staphylococcus aureus on various biomaterial surfaces. J Mech Behav Biomed. Mater. 2017, 65, 872–880. [Google Scholar] [CrossRef]
- Sjollema, J.; van der Mei, H.C.; Hall, C.L.; Peterson, B.W.; de Vries, J.; Song, L.; Jong, E.D.; Busscher, H.J.; Swartjes, J.J.T.M. Detachment and successive re-attachment of multiple, reversibly-binding tethers result in irreversible bacterial adhesion to surfaces. Sci. Rep. 2017, 7, 4369. [Google Scholar] [CrossRef] [Green Version]
- Shlyakhtenko, L.S.; Gall, A.A.; Lyubchenko, Y.L. Mica functionalization for imaging of DNA and protein-DNA complexes with atomic force microscopy. Methods Mol. Biol. 2013, 931, 295–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, C.J.; Venkataraman, S.; Retterer, S.T.; Allison, D.P.; Doktycz, M.J. Comparison of the indentation and elasticity of E. coli and its spheroplasts by AFM. Ultramicroscopy 2007, 107, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Hyldgaard, M.; Mygind, T.; Vad, B.S.; Stenvang, M.; Otzen, D.E.; Meyer, R.L. The antimicrobial mechanism of action of epsilon-poly-l-lysine. Appl. Environ. Microbiol. 2014, 80, 7758–7770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu Quba, A.A.; Schaumann, G.E.; Karagulyan, M.; Diehl, D. A new approach for repeated tip-sample relocation for AFM imaging of nano and micro sized particles and cells in liquid environment. Ultramicroscopy 2020, 211, 112945. [Google Scholar] [CrossRef] [PubMed]
- Müller, D.J.; Fotiadis, D.; Scheuring, S.; Müller, S.A.; Engel, A. Electrostatically balanced subnanometer imaging of biological specimens by atomic force microscope. Biophys. J. 1999, 76, 1101–1111. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.F.; Chopra, T.; Kaye, K.S. Pathogens Resistant to Antibacterial Agents. Med. Clin. N. Am. 2011, 95, 647–676. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention—Anitbiotic/Antimicrobial Resistance. Available online: https://www.cdc.gov/ (accessed on 20 July 2017).
- Willyard, C. The drug-resistant bacteria that pose the greatest health threats. Nat. News 2017, 543, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clutter, D.S.; Jordan, M.R.; Bertagnolio, S.; Shafer, R.W. HIV-1 drug resistance and resistance testing. Infect. Genet. Evol. 2016, 46, 292–307. [Google Scholar] [CrossRef] [Green Version]
- Ikram, A.; Obaid, A.; Awan, F.M.; Hanif, R.; Naz, A.; Paracha, R.Z.; Ali, A.; Janjua, H.A. Identification of drug resistance and immune-driven variations in hepatitis C virus (HCV) NS3/4A, NS5A and NS5B regions reveals a new approach toward personalized medicine. Antiviral. Res. 2017, 137, 112–124. [Google Scholar] [CrossRef]
- Graham, S.V. The human papillomavirus replication cycle, and its links to cancer progression: A comprehensive review. Clin. Sci. (Lond.) 2017, 131, 2201–2221. [Google Scholar] [CrossRef] [Green Version]
- Copin, R.; Sause, W.E.; Fulmer, Y.; Balasubramanian, D.; Dyzenhaus, S.; Ahmed, J.M.; Kumar, K.; Lees, J.; Stachel, A.; Fisher, J.C.; et al. Sequential evolution of virulence and resistance during clonal spread of community-acquired methicillin-resistant Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouloudi, E.; Protonotariou, E.; Zagorianou, A.; Iosifidis, E.; Karapanagiotou, A.; Giasnetsova, T.; Tsioka, A.; Roilides, E.; Sofianou, D.; Gritsi-Gerogianni, N. Bloodstream infections caused by metallo-beta-lactamase/Klebsiella pneumoniae carbapenemase-producing K. pneumoniae among intensive care unit patients in Greece: Risk factors for infection and impact of type of resistance on outcomes. Infect. Control Hosp. Epidemiol. 2010, 31, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Austin, N.; Ordonez, A.A.; Hsu, A.J.; Benson, J.E.; Mahesh, M.; Menachery, E.; Razeq, J.H.; Salfinger, M.; Starke, J.R.; Milstone, A.M.; et al. Extensively drug-resistant tuberculosis in a young child after travel to India. Lancet Infect. Dis. 2015, 15, 1485–1491. [Google Scholar] [CrossRef] [Green Version]
- Plomp, M.; Leighton, T.J.; Wheeler, K.E.; Hill, H.D.; Malkin, A.J. In vitro high-resolution structural dynamics of single germinating bacterial spores. Proc. Natl. Acad. Sci. USA 2007, 104, 9644–9649. [Google Scholar] [CrossRef] [Green Version]
- Butt, H.J.; Wolff, E.K.; Gould, S.A.; Dixon Northern, B.; Peterson, C.M.; Hansma, P.K. Imaging cells with the atomic force microscope. J. Struct. Biol. 1990, 105, 54–61. [Google Scholar] [CrossRef]
- Ding, L.; Yang, L.; Weiss, T.M.; Waring, A.J.; Lehrer, R.I.; Huang, H.W. Interaction of antimicrobial peptides with lipopolysaccharides. Biochemistry 2003, 42, 12251–12259. [Google Scholar] [CrossRef] [Green Version]
- Overton, K.; Greer, H.M.; Ferguson, M.A.; Spain, E.M.; Elmore, D.E.; Núñez, M.E.; Volle, C.B. Qualitative and Quantitative Changes to. Langmuir 2020, 36, 650–659. [Google Scholar] [CrossRef]
- Domingues, M.M.; Silva, P.M.; Franquelim, H.G.; Carvalho, F.A.; Castanho, M.A.; Santos, N.C. Antimicrobial protein rBPI21-induced surface changes on Gram-negative and Gram-positive bacteria. Nanomedicine 2014, 10, 543–551. [Google Scholar] [CrossRef]
- Oh, Y.J.; Plochberger, B.; Rechberger, M.; Hinterdorfer, P. Characterizing the effect of polymyxin B antibiotics to lipopolysaccharide on Escherichia coli surface using atomic force microscopy. J. Mol. Recognit. 2017, 30. [Google Scholar] [CrossRef]
- Daugelavicius, R.; Bakiene, E.; Bamford, D.H. Stages of polymyxin B interaction with the Escherichia coli cell envelope. Antimicrob. Agents Chemother. 2000, 44, 2969–2978. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, M.; Godino, A.; Príncipe, A.; Morales, G.M.; Fischer, S. Effect of a Pseudomonas fluorescens tailocin against phytopathogenic Xanthomonas observed by atomic force microscopy. J. Biotechnol. 2017, 256, 13–20. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Di Bonaventura, I.; Visini, R.; Gan, B.H.; Fu, Y.; Probst, D.; Lüscher, A.; Köhler, T.; van Delden, C.; Stocker, A.; et al. Design, crystal structure and atomic force microscopy study of thioether ligated d,l-cyclic antimicrobial peptides against multidrug resistant Pseudomonas aeruginosa. Chem. Sci. 2017, 8, 7464–7475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carneiro, V.A.; de Oliveira, S.T.; Silva, R.L.; Duarte, H.S.; Silva, M.L.; Matos, M.N.C.; Cavalcante, R.M.B.; Figueira, C.S.; Lorenzón, E.N.; Cilli, E.M.; et al. Antimicrobial and Antibiofilm Activity of Lys-[Trp6]hy-a1 Combined with Ciprofloxacin Against Gram-negative Bacteria. Protein Pept. Lett. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.; Bessa, L.J.; Sousa, C.F.; Eaton, P.; Bongiorno, D.; Stefani, S.; Campanile, F.; Gameiro, P. Fluoroquinolone Metalloantibiotics: A Promising Approach against Methicillin-Resistant. Int. J. Environ. Res. Public Health 2020, 17, 3127. [Google Scholar] [CrossRef] [PubMed]
- Chmielewska, S.J.; Skłodowski, K.; Piktel, E.; Suprewicz, Ł.; Fiedoruk, K.; Daniluk, T.; Wolak, P.; Savage, P.B.; Bucki, R. NDM-1 Carbapenemase-Producing Enterobacteriaceae are Highly Susceptible to Ceragenins CSA-13, CSA-44, and CSA-131. Infect. Drug Resist. 2020, 13, 3277–3294. [Google Scholar] [CrossRef]
- Wnorowska, U.; Piktel, E.; Durnaś, B.; Fiedoruk, K.; Savage, P.B.; Bucki, R. Use of ceragenins as a potential treatment for urinary tract infections. BMC Infect. Dis. 2019, 19, 369. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Hussain, A.; Kumar Singh, S. Morphological transitions of Bacillus subtilis in the presence of food-grade lipidic nanoemulsions. J. Food Sci. 2020, 85, 1223–1230. [Google Scholar] [CrossRef]
- Quilès, F.; Accoceberry, I.; Couzigou, C.; Francius, G.; Noël, T.; El-Kirat-Chatel, S. AFM combined to ATR-FTIR reveals Candida cell wall changes under caspofungin treatment. Nanoscale 2017, 9, 13731–13738. [Google Scholar] [CrossRef]
- Shahina, Z.; El-Ganiny, A.M.; Minion, J.; Whiteway, M.; Sultana, T.; Dahms, T.E.S. Cinnamomum zeylancium bark essential oil induces cell wall remodelling and spindle defects in Candida albicans. Fungal Biol. Biotechnol. 2018, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Hasim, S.; Allison, D.P.; Retterer, S.T.; Hopke, A.; Wheeler, R.T.; Doktycz, M.J.; Reynolds, T.B. β-(1,3)-Glucan Unmasking in Some Candida albicans Mutants Correlates with Increases in Cell Wall Surface Roughness and Decreases in Cell Wall Elasticity. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Luo, M.; Fu, Y.J.; Zu, Y.G.; Wang, W.; Zhang, L.; Yao, L.P.; Zhao, C.J.; Sun, Y. Effect of corilagin on membrane permeability of Escherichia coli, Staphylococcus aureus and Candida albicans. Phytother. Res. 2013, 27, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- Bucki, R.; Niemirowicz-Laskowska, K.; Deptuła, P.; Wilczewska, A.Z.; Misiak, P.; Durnaś, B.; Fiedoruk, K.; Piktel, E.; Mystkowska, J.; Janmey, P.A. Susceptibility of microbial cells to the modified PIP 2-binding sequence of gelsolin anchored on the surface of magnetic nanoparticles. J. Nanobiotechnol. 2019, 17, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucki, R.; Durnaś, B.; Wątek, M.; Piktel, E.; Cruz, K.; Wolak, P.; Savage, P.B.; Janmey, P.A. Targeting polyelectrolyte networks in purulent body fluids to modulate bactericidal properties of some antibiotics. Infect. Drug Resist. 2018, 11, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemirowicz, K.; Surel, U.; Wilczewska, A.Z.; Mystkowska, J.; Piktel, E.; Gu, X.; Namiot, Z.; Akowska, A.K.L.; Savage, P.B.; Bucki, R. Bactericidal activity and biocompatibility of ceragenin-coated magnetic nanoparticles. J. Nanobiotechnol. 2015, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, P.F.N.; Lima, P.G.; Freitas, C.D.T.; Sousa, D.O.B.; Neto, N.A.S.; Dias, L.P.; Vasconcelos, I.M.; Freitas, L.B.N.; Silva, R.G.G.; Sousa, J.S.; et al. Antidermatophytic Activity of Synthetic Peptides: Action mechanisms and clinical application as adjuvants to enhance the activity and decrease the toxicity of Griseofulvin. Mycoses 2020. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Borah, S.N.; Kandimalla, R.; Bora, A.; Deka, S. Efficacy of a rhamnolipid biosurfactant to inhibit Trichophyton rubrum in vitro and in a mice model of dermatophytosis. Exp. Dermatol. 2019, 28, 601–608. [Google Scholar] [CrossRef]
- Godon, C.; Teulon, J.M.; Odorico, M.; Basset, C.; Meillan, M.; Vellutini, L.; Chen, S.W.; Pellequer, J.L. Conditions to minimize soft single biomolecule deformation when imaging with atomic force microscopy. J. Struct. Biol. 2017, 197, 322–329. [Google Scholar] [CrossRef]
- Kämmer, E.; Götz, I.; Bocklitz, T.; Stöckel, S.; Dellith, A.; Cialla-May, D.; Weber, K.; Zell, R.; Dellith, J.; Deckert, V.; et al. Single particle analysis of herpes simplex virus: Comparing the dimensions of one and the same virions via atomic force and scanning electron microscopy. Anal. Bioanal. Chem. 2016, 408, 4035–4041. [Google Scholar] [CrossRef]
- de Pablo, P.J. Atomic force microscopy of viruses. Subcell. Biochem. 2013, 68, 247–271. [Google Scholar] [CrossRef]
- Barinov, N.; Ivanov, N.; Kopylov, A.; Klinov, D.; Zavyalova, E. Direct visualization of the oligomeric state of hemagglutinins of influenza virus by high-resolution atomic force microscopy. Biochimie 2018, 146, 148–155. [Google Scholar] [CrossRef]
- Azinas, S.; Bano, F.; Torca, I.; Bamford, D.H.; Schwartz, G.A.; Esnaola, J.; Oksanen, H.M.; Richter, R.P.; Abrescia, N.G. Membrane-containing virus particles exhibit the mechanics of a composite material for genome protection. Nanoscale 2018, 10, 7769–7779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Pablo, P.J.; Hernando-Pérez, M.; Carrasco, C.; Carrascosa, J.L. Direct visualization of single virus restoration after damage in real time. J. Biol. Phys. 2018, 44, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Longo, G.; Rio, L.M.; Trampuz, A.; Dietler, G.; Bizzini, A.; Kasas, S. Antibiotic-induced modifications of the stiffness of bacterial membranes. J. Microbiol. Methods 2013, 93, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Garrett, T.R.; Bhakoo, M.; Zhang, Z. Bacterial adhesion and biofilms on surfaces. Prog. Nat. Sci. 2008, 18, 1049–1056. [Google Scholar] [CrossRef]
- Rozenbaum, R.T.; van der Mei, H.C.; Woudstra, W.; de Jong, E.D.; Busscher, H.J.; Sharma, P.K. Role of viscoelasticity in bacterial killing by antimicrobials in differently grown Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 2019, 63, e01972-18. [Google Scholar] [CrossRef] [Green Version]
- Sagar, S.; Kaistha, S.; Das, A.J.; Kumar, R. Nanotechnology: A Twenty-First-Century Approach Towards the Control of Antibiotic-Resistant Bacteria. In Antibiotic Resistant Bacteria: A Challenge to Modern Medicine; Springer: Singapore, 2019; pp. 135–151. [Google Scholar] [CrossRef]
- Chhikara, B.S. Current trends in nanomedicine and nanobiotechnology research. J. Mater. NanoScience 2017, 4, 19–24. [Google Scholar]
- da Silva, A.; Teschke, O. Effects of the antimicrobial peptide PGLa on live Escherichia coli. Biochim. Biophys. Acta 2003, 1643, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.; Wang, K.; He, X.; Zhao, X.J.; Drake, T.; Wang, L.; Bagwe, R.P. Bionanotechnology based on silica nanoparticles. Med. Res. Rev. 2004, 24, 621–638. [Google Scholar] [CrossRef]
- Mathelié-Guinlet, M.; Grauby-Heywang, C.; Martin, A.; Février, H.; Morote, F.; Vilquin, A.; Beven, L.; Delville, M.-H.; Cohen-Bouhacina, T. Detrimental impact of silica nanoparticles on the nanomechanical properties of Escherichia coli, studied by AFM. J. Colloid Interface Sci. 2018, 529, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Uzoechi, S.C.; Abu-Lail, N.I. Variations in the Morphology, Mechanics and Adhesion of Persister and Resister. Antibiotics 2020, 9, 235. [Google Scholar] [CrossRef]
- Krce, L.; Šprung, M.; Rončević, T.; Maravić, A.; Čikeš Čulić, V.; Blažeka, D.; Krstulović, N.; Aviani, I. Probing the Mode of Antibacterial Action of Silver Nanoparticles Synthesized by Laser Ablation in Water: What Fluorescence and AFM Data Tell Us. Nanomaterials 2020, 10, 1040. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, S.; Silva, P.M.; Felício, M.R.; De Medeiros, L.N.; Kurtenbach, E.; Santos, N.C. Psd1 effects on Candida albicans planktonic cells and biofilms. Front. Cellular Infect. Microbiol. 2017, 7, 249. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, S.; Teixeira, A.; Abade, J.; de Medeiros, L.N.; Kurtenbach, E.; Santos, N.C. Evaluation of the membrane lipid selectivity of the pea defensin Psd1. Biochim. Biophys. Acta (BBA)-Biomembranes 2012, 1818, 1420–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Rosmalen, M.G.M.; Nemerow, G.R.; Wuite, G.J.L.; Roos, W.H. A single point mutation in precursor protein VI doubles the mechanical strength of human adenovirus. J. Biol. Phys. 2018, 44, 119–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assaidi, A.; Ellouali, M.; Latrache, H.; Mabrouki, M.; Timinouni, M.; Zahir, H.; Tankiouine, S.; Barguigua, A.; Mliji, E.M. Adhesion of Legionella pneumophila on glass and plumbing materials commonly used in domestic water systems. Int. J. Environ. Health Res. 2018. [Google Scholar] [CrossRef]
- Binggeli, M.; Mate, C.M. Influence of capillary condensation of water on nanotribology studied by force microscopy. Appl. Phys. Lett. 1994, 65, 415–417. [Google Scholar] [CrossRef]
- Fang, H.H.; Chan, K.Y.; Xu, L.C. Quantification of bacterial adhesion forces using atomic force microscopy (AFM). J. Microbiol. Methods 2000, 40, 89–97. [Google Scholar] [CrossRef]
- Pembrey, R.S.; Marshall, K.C.; Schneider, R.P. Cell surface analysis techniques: What do cell preparation protocols do to cell surface properties? Appl. Environ. Microbiol. 1999, 65, 2877–2894. [Google Scholar] [CrossRef] [Green Version]
- Robelek, R.; Wegener, J. Label-free and time-resolved measurements of cell volume changes by surface plasmon resonance (SPR) spectroscopy. Biosens. Bioelectron. 2010, 25, 1221–1224. [Google Scholar] [CrossRef]
- Beaussart, A.; El-Kirat-Chatel, S. Microbial adhesion and ultrastructure from the single-molecule to the single-cell levels by Atomic Force Microscopy. Cell Surf. 2019, 5, 100031. [Google Scholar] [CrossRef]
- Wang, Y.; Ren, B.; Zhou, X.; Liu, S.; Zhou, Y.; Li, B.; Jiang, Y.; Li, M.; Feng, M.; Cheng, L. Growth and adherence of Staphylococcus aureus were enhanced through the PGE2 produced by the activated COX-2/PGE2 pathway of infected oral epithelial cells. PLoS ONE 2017, 12, e0177166. [Google Scholar] [CrossRef] [PubMed]
- Viela, F.; Speziale, P.; Pietrocola, G.; Dufrêne, Y.F. Mechanostability of the Fibrinogen Bridge between Staphylococcal Surface Protein ClfA and Endothelial Cell Integrin α. Nano Lett. 2019, 19, 7400–7410. [Google Scholar] [CrossRef]
- Guillaume-Gentil, O.; Potthoff, E.; Ossola, D.; Franz, C.M.; Zambelli, T.; Vorholt, J.A. Force-controlled manipulation of single cells: From AFM to FluidFM. Trends Biotechnol. 2014, 32, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Formosa, C.; Schiavone, M.; Boisrame, A.; Richard, M.L.; Duval, R.E.; Dague, E. Multiparametric imaging of adhesive nanodomains at the surface of Candida albicans by atomic force microscopy. Nanomedicine 2015, 11, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Kirat-Chatel, S.; Beaussart, A.; Derclaye, S.; Alsteens, D.; Kucharíková, S.; Van Dijck, P.; Dufrêne, Y.F. Force nanoscopy of hydrophobic interactions in the fungal pathogen Candida glabrata. ACS Nano 2015, 9, 1648–1655. [Google Scholar] [CrossRef] [PubMed]
- Valotteau, C.; Prystopiuk, V.; Cormack, B.P.; Dufrêne, Y.F. Atomic Force Microscopy Demonstrates that Candida glabrata Uses Three Epa Proteins To Mediate Adhesion to Abiotic Surfaces. mSphere 2019, 4. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Zhou, S.; Xu, X.; Li, D.; Lin, Y.; Lyu, F.; Dong, M. A Multi-Scale Approach to Investigate Adhesion Properties of. Foods 2020, 9, 912. [Google Scholar] [CrossRef]
- Ma, S.; Ge, W.; Yan, Y.; Huang, X.; Ma, L.; Li, C.; Yu, S.; Chen, C. Effects of Streptococcus sanguinis Bacteriocin on Deformation, Adhesion Ability, and Young’s Modulus of Candida albicans. Biomed. Res. Int. 2017, 2017, 5291486. [Google Scholar] [CrossRef]
- Ma, S.; Li, H.; Yan, C.; Wang, D.; Xia, X.; Dong, X.; Zhao, Y.; Sun, T.; Hu, P.; Guan, W. Antagonistic effect of protein extracts from. Exp. Ther. Med. 2014, 7, 1486–1494. [Google Scholar] [CrossRef]
- Newton, R.; Delguste, M.; Koehler, M.; Dumitru, A.C.; Laskowski, P.R.; Müller, D.J.; Alsteens, D. Combining confocal and atomic force microscopy to quantify single-virus binding to mammalian cell surfaces. Nat. Protoc. 2017, 12, 2275–2292. [Google Scholar] [CrossRef]
- Lin, S.; Hong, C.H.; Sheu, B.C.; Wu, L.X.; Huang, W.C.; Guo, C.Y. Dynamic response of HPV16/anti-HPV16 pairs with unbinding events studied by atomic force microscopy. Biochem. Biophys. Rep. 2016, 6, 203–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Petitjean, S.J.L.; Koehler, M.; Zhang, Q.; Dumitru, A.C.; Chen, W.; Derclaye, S.; Vincent, S.P.; Soumillion, P.; Alsteens, D. Molecular interaction and inhibition of SARS-CoV-2 binding to the ACE2 receptor. Nat. Commun. 2020, 11, 4541. [Google Scholar] [CrossRef] [PubMed]
- Aguayo, S.; Marshall, H.; Pratten, J.; Bradshaw, D.; Brown, J.S.; Porter, S.R.; Spratt, D.; Bozec, L. Early Adhesion of Candida albicans onto Dental Acrylic Surfaces. J. Dent. Res. 2017, 96, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Ozel, G.S.; Guneser, M.B.; Inan, O.; Eldeniz, A.U. Evaluation of C. albicans and S. Mutans adherence on different provisional crown materials. J. Adv. Prosthodont. 2017, 9, 335–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Blanco, D.; Lynn, A.; Rosch, J.; Noreldin, R.; Salerni, A.; Lambert, C.; Rao, R.P. A pre-therapeutic coating for medical devices that prevents the attachment of Candida albicans. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carniello, V.; Harapanahalli, A.K.; Busscher, H.J.; van der Mei, H.C. Adhesion force sensing and activation of a membrane-bound sensor to activate nisin efflux pumps in Staphylococcus aureus under mechanical and chemical stresses. J. Colloid Interface Sci. 2018, 512, 14–20. [Google Scholar] [CrossRef]
- van Belkum, A.; Verkaik, N.J.; de Vogel, C.P.; Boelens, H.A.; Verveer, J.; Nouwen, J.L.; Verbrugh, H.A.; Wertheim, H.F. Reclassification of Staphylococcus aureus nasal carriage types. J. Infect. Dis. 2009, 199, 1820–1826. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.C.; Cheng, Y.W.; Liu, C.C.; Peng, X.Y.; Yung, M.C.; Liu, T.Y. Anti-Bacterial and Anti-Fouling Capabilities of Poly(3,4-Ethylenedioxythiophene) Derivative Nanohybrid Coatings on SUS316L Stainless Steel by Electrochemical Polymerization. Polymers 2020, 12, 1467. [Google Scholar] [CrossRef]
- Hu, C.H.; Xu, Z.R.; Xia, M.S. Antibacterial effect of Cu2+-exchanged montmorillonite on Aeromonas hydrophila and discussion on its mechanism. Vet. Microbiol. 2005, 109, 83–88. [Google Scholar] [CrossRef]
- Gião, M.S.; Wilks, S.A.; Keevil, C.W. Influence of copper surfaces on biofilm formation by Legionella pneumophila in potable water. Biometals 2015, 28, 329–339. [Google Scholar] [CrossRef]
- Lehtola, M.J.; Miettinen, I.T.; Keinänen, M.M.; Kekki, T.K.; Laine, O.; Hirvonen, A.; Vartiainen, T.; Martikainen, P.J. Microbiology, chemistry and biofilm development in a pilot drinking water distribution system with copper and plastic pipes. Water Res. 2004, 38, 3769–3779. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, C.K.; Subbiahdoss, G.; Zeng, G.; Salmi, Z.; Kjems, J.; Mygind, T.; Snabe, T.; Meyer, R.L. Antibacterial isoeugenol coating on stainless steel and polyethylene surfaces prevents biofilm growth. J. Appl. Microbiol. 2018, 124, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, L.P.; Miñán, A.; Benítez, G.; de Mele, M.F.L.; Vela, M.E.; Schilardi, P.L.; Ferreira-Neto, E.P.; Noveletto, J.C.; Correr, W.R.; Rodrigues-Filho, U.P. Self-sterilizing ormosils surfaces based on photo-synzthesized silver nanoparticles. Colloids Surf. B Biointerfaces 2017, 164, 144–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wnorowska, U.; Watek, M.; Durnas, B.; Gluszek, K.; Piktel, E.; Niemirowicz, K.; Bucki, R. Extracellular DNA as an essential component and therapeutic target of microbial biofilm. Med. Stud. Studia Med. 2015, 31, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Sportelli, M.C.; Tütüncü, E.; Picca, R.A.; Valentini, M.; Valentini, A.; Kranz, C.; Mizaikoff, B.; Barth, H.; Cioffi, N. Inhibiting P. fluorescens biofilms with fluoropolymer-embedded silver nanoparticles: An in-situ spectroscopic study. Sci. Rep. 2017, 7, 11870. [Google Scholar] [CrossRef]
- Santos, S.M.; Carbajo, J.M.; Gómez, N.; Ladero, M.; Villar, J.C. Modification of Bacterial Cellulose Biofilms with Xylan Polyelectrolytes. Bioengineering 2017, 4, 93. [Google Scholar] [CrossRef] [Green Version]
- Quatrin, P.M.; Verdi, C.M.; de Souza, M.E.; de Godoi, S.N.; Klein, B.; Gundel, A.; Wagner, R.; de Almeida Vaucher, R.; Ourique, A.F.; Santos, R.C.V. Antimicrobial and antibiofilm activities of nanoemulsions containing Eucalyptus globulus oil against Pseudomonas aeruginosa and Candida spp. Microb. Pathog. 2017, 112, 230–242. [Google Scholar] [CrossRef]
- Wang, H.; Wilksch, J.J.; Strugnell, R.A.; Gee, M.L. Role of capsular polysaccharides in biofilm formation: An AFM nanomechanics study. ACS Appl. Mater. Interfaces 2015, 7, 13007–13013. [Google Scholar] [CrossRef]
- Daniels, S.L.; Pressman, J.G.; Wahman, D.G. AFM structural characterization of drinking water biofilm under physiological conditions. RSC Adv. 2016, 6, 5812–5816. [Google Scholar] [CrossRef]
- Kwon, T.-H.; Kim, S. Measuring elastic modulus of bacterial biofilms in a liquid phase using atomic force microscopy. Geomechanics Eng. 2017, 12, 863–870. [Google Scholar]
- de Souza, M.E.; Clerici, D.J.; Verdi, C.M.; Fleck, G.; Quatrin, P.M.; Spat, L.E.; Bonez, P.C.; Santos, C.F.D.; Antoniazzi, R.P.; Zanatta, F.B.; et al. Antimicrobial activity of Melaleuca alternifolia nanoparticles in polymicrobial biofilm in situ. Microb. Pathog. 2017, 113, 432–437. [Google Scholar] [CrossRef] [PubMed]
- De, A.; Raj, H.J.; Haldar, J.; Mukherjee, P.; Maiti, P.K. Biofilm colonization in chronic treatment refractory infections presenting with discharging sinuses: A study in a tertiary care hospital of Eastern India. J. Lab. Phys. 2017, 9, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Birarda, G.; Delneri, A.; Lagatolla, C.; Parisse, P.; Cescutti, P.; Vaccari, L.; Rizzo, R. Multi-technique microscopy investigation on bacterial biofilm matrices: A study on Klebsiella pneumoniae clinical strains. Anal. Bioanal. Chem. 2019, 411, 7315–7325. [Google Scholar] [CrossRef] [PubMed]
- Handorf, O.; Schnabel, U.; Bösel, A.; Weihe, T.; Bekeschus, S.; Graf, A.C.; Riedel, K.; Ehlbeck, J. Antimicrobial effects of microwave-induced plasma torch (MiniMIP) treatment on Candida albicans biofilms. Microb. Biotechnol. 2019, 12, 1034–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, H.; Aguayo, S.; Kilian, M.; Petersen, F.; Bozec, L.; Brown, J. Relationship between the Nano-Biomechanical Properties of Streptococcal Polysaccharide Capsules and Virulence Phenotype. ACS Nano 2020, 14, 1070–1083. [Google Scholar] [CrossRef] [PubMed]
- Odermatt, P.D.; Hannebelle, M.T.M.; Eskandarian, H.A.; Nievergelt, A.P.; McKinney, J.D.; Fantner, G.E. Overlapping and essential roles for molecular and mechanical mechanisms in mycobacterial cell division. Nat. Phys. 2020, 16, 57–62. [Google Scholar] [CrossRef]
- Hassan, A.A.; Vitorino, M.V.; Robalo, T.; Rodrigues, M.S.; Sá-Correia, I. Variation of Burkholderia cenocepacia cell wall morphology and mechanical properties during cystic fibrosis lung infection, assessed by atomic force microscopy. Sci. Rep. 2019, 9, 16118. [Google Scholar] [CrossRef]
- Madeira, A.; dos Santos, S.C.; Santos, P.M.; Coutinho, C.P.; Tyrrell, J.; McClean, S.; Callaghan, M.; Sá-Correia, I. Proteomic profiling of Burkholderia cenocepacia clonal isolates with different virulence potential retrieved from a cystic fibrosis patient during chronic lung infection. PLoS ONE 2013, 8, e83065. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.C.; Huang, C.; Lai, H.C. Revealing the ultrastructure of the membrane pores of intact. Heliyon 2019, 5, e02636. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, X.; Yang, H.; Yuan, J.; Wei, H.; Yu, J.; Fang, X. Study of the interactions between endolysin and bacterial peptidoglycan on S. aureus by dynamic force spectroscopy. Nanoscale 2015, 7, 15245–15250. [Google Scholar] [CrossRef]
- Iqbal, K.M.; Bertino, M.F.; Shah, M.R.; Ehrhardt, C.J.; Yadavalli, V.K. Nanoscale Phenotypic Textures of. Microorganisms 2020, 8, 160. [Google Scholar] [CrossRef] [Green Version]
- Salive, A.F.V.; Prudêncio, C.V.; Baglinière, F.; Oliveira, L.L.; Ferreira, S.O.; Vanetti, M.C.D. Comparison of stress conditions to induce viable but non-cultivable state in Salmonella. Braz. J. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Qiao, H.; Krajcikova, D.; Zhang, Z.; Wang, H.; Barak, I.; Tang, J. Physical interaction and assembly of Bacillus subtilis spore coat proteins CotE and CotZ studied by atomic force microscopy. J. Struct. Biol. 2016, 195, 245–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokajuk, G.; Niemirowicz, K.; Deptula, P.; Piktel, E.; Ciesluk, M.; Wilczewska, A.Z.; Dabrowski, J.R.; Bucki, R. Use of magnetic nanoparticles as a drug delivery system to improve chlorhexidine antimicrobial activity. Int. J. Nanomed. 2017, 12, 7833–7846. [Google Scholar] [CrossRef] [Green Version]
- Niemirowicz, K.; Durnaś, B.; Piktel, E.; Bucki, R. Development of antifungal therapies using nanomaterials. Nanomedicine 2017, 12, 1891–1905. [Google Scholar] [CrossRef]
- Chen, N.; Wang, H.; Huang, Q.; Li, J.; Yan, J.; He, D.; Fan, C.; Song, H. Long-term effects of nanoparticles on nutrition and metabolism. Small 2014, 10, 3603–3611. [Google Scholar] [CrossRef]
- Mendonça, M.C.; Soares, E.S.; de Jesus, M.B.; Ceragioli, H.J.; Irazusta, S.P.; Batista, Â.; Vinolo, M.A.; Maróstica Júnior, M.R.; da Cruz-Höfling, M.A. Reduced graphene oxide: Nanotoxicological profile in rats. J. Nanobiotechnol. 2016, 14, 53. [Google Scholar] [CrossRef] [Green Version]
- Vila, L.; Marcos, R.; Hernández, A. Long-term effects of silver nanoparticles in caco-2 cells. Nanotoxicology 2017, 11, 771–780. [Google Scholar] [CrossRef]
- Michalak, G.; Gluszek, K.; Piktel, E.; Deptula, P.; Puszkarz, I.; Niemirowicz, K.; Bucki, R. Polymeric nanoparticles—A novel solution for delivery of antimicrobial agents. Med. Stud. Studia Med. 2016, 32, 56–62. [Google Scholar] [CrossRef]
- Ertem, E.; Gutt, B.; Zuber, F.; Allegri, S.; Le Ouay, B.; Mefti, S.; Formentin, K.; Stellacci, F.; Ren, Q. Core-Shell Silver Nanoparticles in Endodontic Disinfection Solutions Enable Long-Term Antimicrobial Effect on Oral Biofilms. ACS Appl. Mater. Interfaces 2017. [Google Scholar] [CrossRef]
- Salvioni, L.; Galbiati, E.; Collico, V.; Alessio, G.; Avvakumova, S.; Corsi, F.; Tortora, P.; Prosperi, D.; Colombo, M. Negatively charged silver nanoparticles with potent antibacterial activity and reduced toxicity for pharmaceutical preparations. Int. J. Nanomed. 2017, 12, 2517–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlov, I.A.; Sankova, T.P.; Babich, P.S.; Sosnin, I.M.; Ilyechova, E.Y.; Kirilenko, D.A.; Brunkov, P.N.; Ataev, G.L.; Romanov, A.E.; Puchkova, L.V. New silver nanoparticles induce apoptosis-like process in E. coli and interfere with mammalian copper metabolism. Int. J. Nanomed. 2016, 11, 6561–6574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Shawafi, W.M.; Salah, N.; Alshahrie, A.; Ahmed, Y.M.; Moselhy, S.S.; Hammad, A.H.; Hussain, M.A.; Memic, A. Size controlled ultrafine CeO2 nanoparticles produced by the microwave assisted route and their antimicrobial activity. J. Mater. Sci. Mater. Med. 2017, 28, 177. [Google Scholar] [CrossRef] [PubMed]
- Muniyan, A.; Ravi, K.; Mohan, U.; Panchamoorthy, R. Characterization and in vitro antibacterial activity of saponin-conjugated silver nanoparticles against bacteria that cause burn wound infection. World J. Microbiol. Biotechnol. 2017, 33, 147. [Google Scholar] [CrossRef]
- Alavi, M.; Karimi, N. Characterization, antibacterial, total antioxidant, scavenging, reducing power and ion chelating activities of green synthesized silver, copper and titanium dioxide nanoparticles using Artemisia haussknechtii leaf extract. Artif. Cells Nanomed. Biotechnol. 2017. [Google Scholar] [CrossRef] [Green Version]
- Raj, R.; Das, S. Development and application of anticancer fluorescent CdS nanoparticles enriched Lactobacillus bacteria as therapeutic microbots for human breast carcinoma. Appl. Microbiol. Biotechnol. 2017, 101, 5439–5451. [Google Scholar] [CrossRef]
- Steffy, K.; Shanthi, G.; Maroky, A.S.; Selvakumar, S. Enhanced antibacterial effects of green synthesized ZnO NPs using Aristolochia indica against Multi-drug resistant bacterial pathogens from Diabetic Foot Ulcer. J. Infect. Public Health 2017. [Google Scholar] [CrossRef]
- Rao, Y.; Inwati, G.K.; Singh, M. Green synthesis of capped gold nanoparticles and their effect on Gram-positive and Gram-negative bacteria. Future Sci. OA 2017, 3, FSO239. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.-E.; Xing, L.; Hou, B.; Ge, H.; Guo, X.; Tang, Z. Inorganic nano mental oxides used as anti-microorganism agents for pathogen control. Curr. Res. Technol. Education. Top. Appl. Microbiol. Microbial. 2010, 2, 361–368. [Google Scholar]
- Wang, L.; Hu, C.; Shao, L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Feng, X.; Werber, J.R.; Chu, C.; Zucker, I.; Kim, J.H.; Osuji, C.O.; Elimelech, M. Enhanced antibacterial activity through the controlled alignment of graphene oxide nanosheets. Proc. Natl. Acad. Sci. USA 2017, 114, E9793–E9801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maktedar, S.S.; Mehetre, S.S.; Avashthi, G.; Singh, M. In situ sonochemical reduction and direct functionalization of graphene oxide: A robust approach with thermal and biomedical applications. Ultrason Sonochem. 2017, 34, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, T.I.; Fouda, A. Green approach for one-pot synthesis of silver nanorod using cellulose nanocrystal and their cytotoxicity and antibacterial assessment. Int. J. Biol. Macromol. 2018, 106, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Dobrucka, R.; Dlugaszewska, J.; Kaczmarek, M. Cytotoxic and antimicrobial effects of biosynthesized ZnO nanoparticles using of Chelidonium majus extract. Biomed. Microdevices 2017, 20, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mussap, M.; Molinari, M.; Senno, E.; Gritti, P.; Soro, B.; Mannelli, S.; Fabris, C. New diagnostic tools for neonatal sepsis: The role of a real-time polymerase chain reaction for the early detection and identification of bacterial and fungal species in blood samples. J. Chemother. 2007, 19, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Razik, A.; Eldars, W.; Elhelaly, R.; Eldeeb, A.A.; Abdelsalam, M.; El-Wakeel, N.; Aboulmagd, A. Homocysteine: A new diagnostic marker in spontaneous bacterial peritonitis. Eur. J. Gastroenterol. Hepatol. 2018, 30, 779–785. [Google Scholar] [CrossRef]
- Perez-Guaita, D.; Kochan, K.; Batty, M.; Doerig, C.; Garcia-Bustos, J.; Espinoza, S.; McNaughton, D.; Heraud, P.; Wood, B.R. Multispectral atomic force microscopy-infrared nano-imaging of malaria infected red blood cells. Anal. Chem. 2018, 90, 3140–3148. [Google Scholar] [CrossRef]
- Etayash, H.; Khan, M.F.; Kaur, K.; Thundat, T. Microfluidic cantilever detects bacteria and measures their susceptibility to antibiotics in small confined volumes. Nat. Commun. 2016, 7, 12947. [Google Scholar] [CrossRef]
- Kasas, S.; Ruggeri, F.S.; Benadiba, C.; Maillard, C.; Stupar, P.; Tournu, H.; Dietler, G.; Longo, G. Detecting nanoscale vibrations as signature of life. Proc. Natl. Acad. Sci. USA 2015, 112, 378–381. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cieśluk, M.; Deptuła, P.; Piktel, E.; Fiedoruk, K.; Suprewicz, Ł.; Paprocka, P.; Kot, P.; Pogoda, K.; Bucki, R. Physics Comes to the Aid of Medicine—Clinically-Relevant Microorganisms through the Eyes of Atomic Force Microscope. Pathogens 2020, 9, 969. https://doi.org/10.3390/pathogens9110969
Cieśluk M, Deptuła P, Piktel E, Fiedoruk K, Suprewicz Ł, Paprocka P, Kot P, Pogoda K, Bucki R. Physics Comes to the Aid of Medicine—Clinically-Relevant Microorganisms through the Eyes of Atomic Force Microscope. Pathogens. 2020; 9(11):969. https://doi.org/10.3390/pathogens9110969
Chicago/Turabian StyleCieśluk, Mateusz, Piotr Deptuła, Ewelina Piktel, Krzysztof Fiedoruk, Łukasz Suprewicz, Paulina Paprocka, Patrycja Kot, Katarzyna Pogoda, and Robert Bucki. 2020. "Physics Comes to the Aid of Medicine—Clinically-Relevant Microorganisms through the Eyes of Atomic Force Microscope" Pathogens 9, no. 11: 969. https://doi.org/10.3390/pathogens9110969
APA StyleCieśluk, M., Deptuła, P., Piktel, E., Fiedoruk, K., Suprewicz, Ł., Paprocka, P., Kot, P., Pogoda, K., & Bucki, R. (2020). Physics Comes to the Aid of Medicine—Clinically-Relevant Microorganisms through the Eyes of Atomic Force Microscope. Pathogens, 9(11), 969. https://doi.org/10.3390/pathogens9110969