Transcriptome Analysis of The Inflammatory Responses of Bovine Mammary Epithelial Cells: Exploring Immunomodulatory Target Genes for Bovine Mastitis

 ,
,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Results
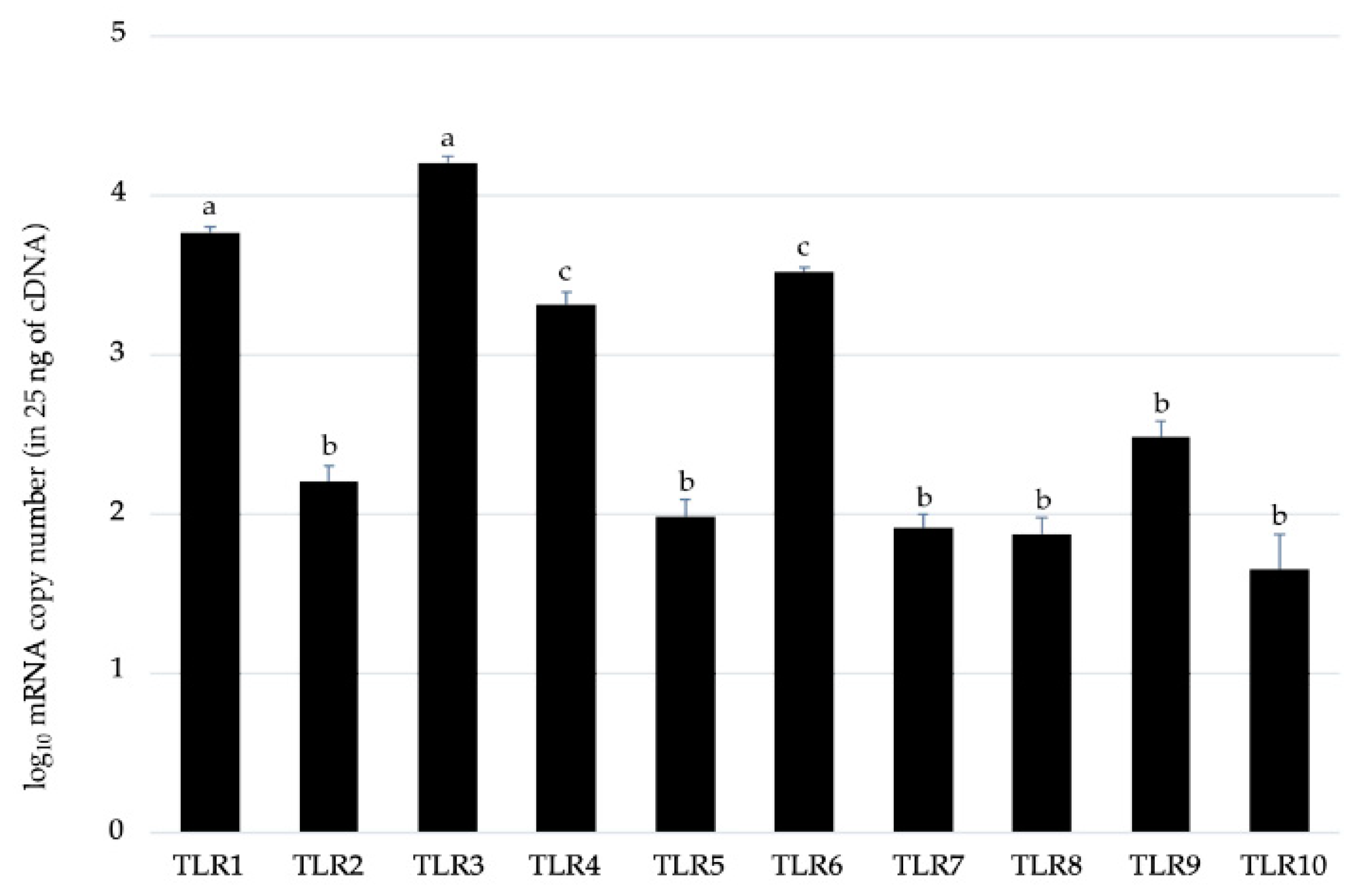
2.1. Expression Patterns of Toll-like Receptors in BME Cells
2.2. Differential Gene Expressions in BME Cells after Stimulation with LPS and Heat-killed S. auresus Strains
2.3. Gene Ontology (GO) Terms Enriched by DEGs
2.4. Differential Responses in Inflammatory Gene Gxpression in BME Cells after Stimulation with LPS and Heat-Killed and S. aureus Strains
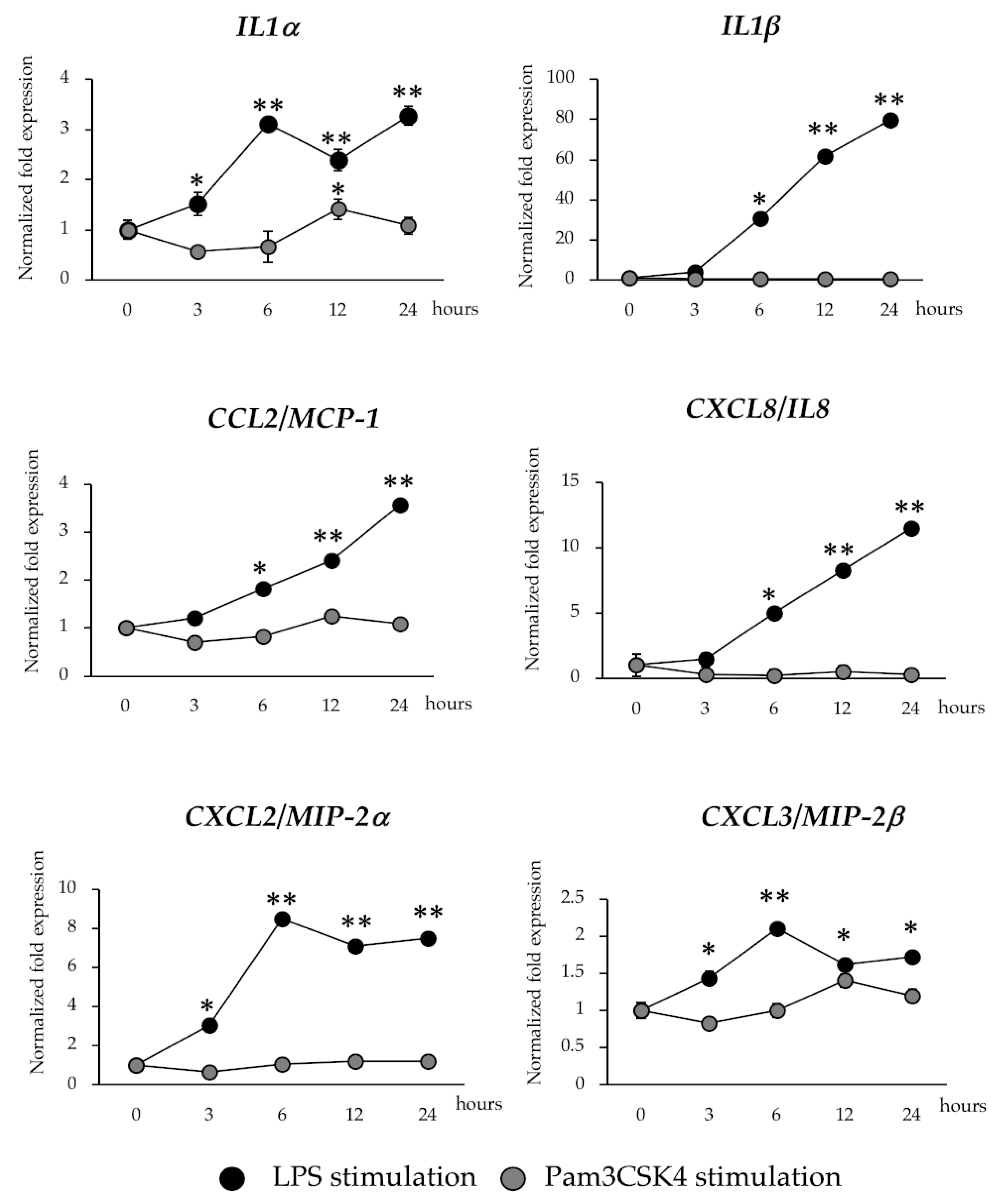
2.5. Inflammatory Gene Expression in BME Cells after LPS or Pam3CSK4 Stimulation
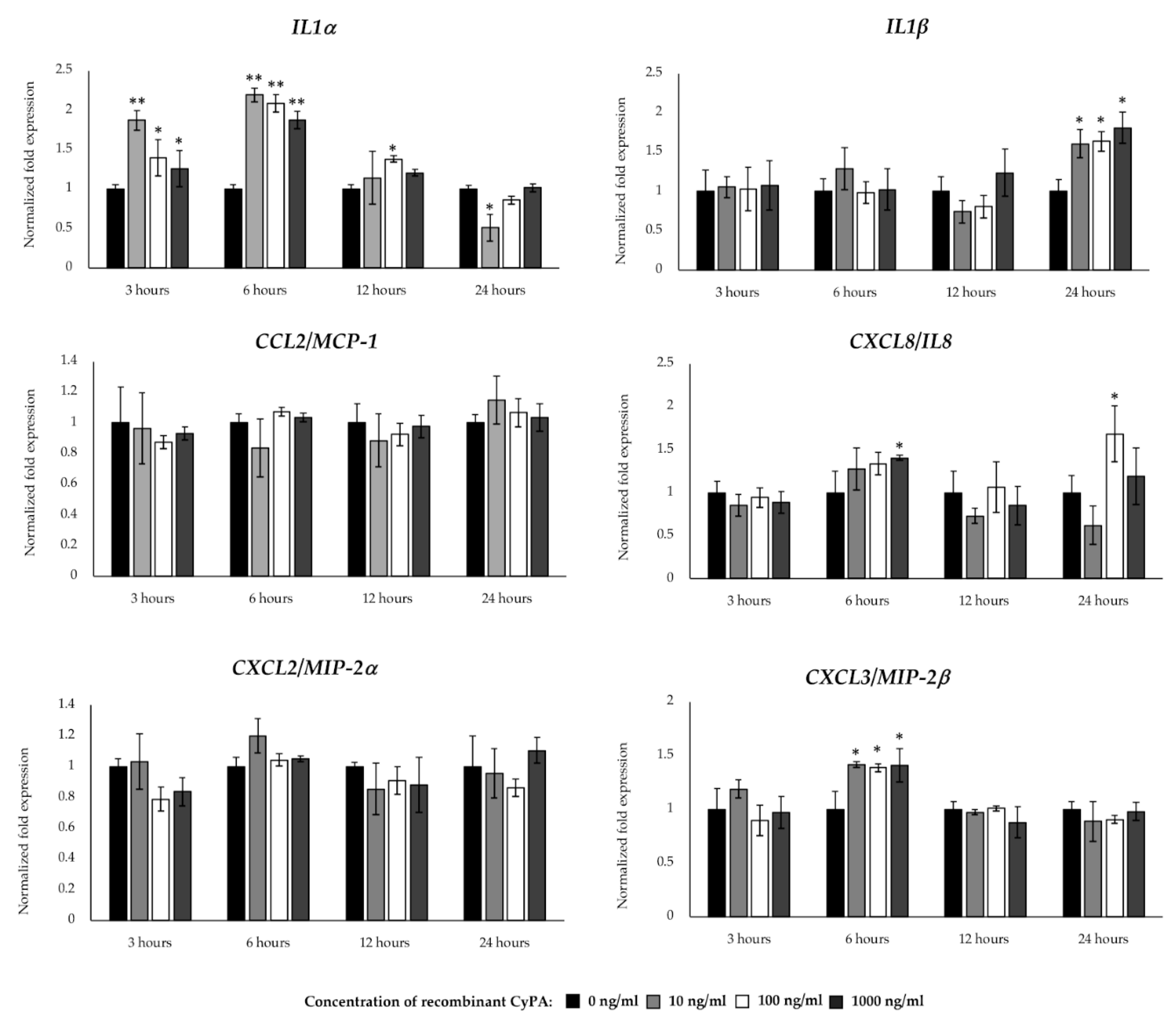
2.6. Inflammatory Gene Expression in BME Cells after CyPA Stimulation
3. Discussion
4. Materials and Methods
4.1. Cell Line
4.2. Growth and Maintenance of BME Cell Line
4.3. TLRs Copy Number Analysis
4.4. Preparations of Stimulants
4.5. Experimental Challenge to BME Cells and Sampling for Gene Expression Study
4.6. RNA Isolation, Labeling and Microarray Hybridization
4.7. Statistical Analysis of Microarray Data
4.8. Gene Ontology Analyses
4.9. RT-qPCR Validation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Seegers, H.; Fourichon, C.; Beaudeau, F. Production effects related to mastitis and mastitis economics in dairy cattle herds. Vet. Res. 2003, 34, 475–491. [Google Scholar] [CrossRef] [Green Version]
- Dos Reis, C.B.M.; Barreiro, J.R.; Mestieri, L.; de Felício Porcionato, M.A.; dos Santos, M.V. Effect of somatic cell count and mastitis pathogens on milk composition in Gyr cows. BMC Vet. Res. 2013, 9, 67. [Google Scholar]
- Pumipuntu, N.; Kulpeanprasit, S.; Santajit, S.; Tunyong, W.; Kong-Ngoen, T.; Hinthong, W.; Indrawattana, N. Screening method for Staphylococcus aureus identification in subclinical bovine mastitis from dairy farms. Vet. World 2017, 10, 721–726. [Google Scholar] [CrossRef] [Green Version]
- Jensen, K.; Günther, J.; Talbot, R.; Petzl, W.; Zerbe, H.; Schuberth, H.J.; Seyfert, H.M.; Glass, E.J. Escherichia coli- and Staphylococcus aureus-induced mastitis differentially modulate transcriptional responses in neighbouring uninfected bovine mammary gland quarters. BMC Genom. 2013, 14, 36. [Google Scholar] [CrossRef] [Green Version]
- Burvenich, C.; Van Merrid, V.; Mehrzad, J.; ez-Fraile, A.; Duchateau, L. Severity of E. coli mastitis is mainly determined by cow factors. Vet. Res. 2003, 34, 521–564. [Google Scholar] [CrossRef] [Green Version]
- Schukken, Y.H.; Günther, J.; Fitzpatrick, J.; Fontaine, M.C.; Goetze, L.; Holst, O.; Leigh, J.; Petzl, W.; Schuberth, H.-J.; Sipka, A.; et al. Host-response patterns of intramammary infections in dairy cows. Vet. Immunol. Immunopathol. 2011, 144, 270–289. [Google Scholar] [CrossRef]
- Mitterhuemer, S.; Petzl, W.; Krebs, S.; Mehne, D.; Klanner, A.; Wolf, E.; Zerbe, H.; Blum, H. Escherichia coli infection induces distinct local and systemic transcriptome responses in the mammary gland. BMC Genom. 2010, 11, 138. [Google Scholar] [CrossRef] [Green Version]
- Bannerman, D.D. Pathogen-dependent induction of cytokines and other soluble inflammatory mediators during intramammary infection of dairy cows. J. Anim. Sci. 2009, 87, 10–25. [Google Scholar] [CrossRef] [Green Version]
- Gresham, H.D.; Lowrance, J.H.; Caver, T.E.; Wilson, B.S.; Cheung, A.L.; Lindberg, F.P. Survival of Staphylococcus aureus inside neutrophils contributes to infection. J. Immunol. 2000, 164, 3713–3722. [Google Scholar] [CrossRef] [Green Version]
- Paape, M.J.; Mehrzad, J.; Zhao, X.; Detilleux, J.; Burvenich, C. Defense of the bovine mammary gland by polymorphonuclear neutrophil leukocytes. J. Mammary Gland Biol. 2002, 7, 109–121. [Google Scholar] [CrossRef]
- Paape, M.J.; Bannerman, D.D.; Zhao, X.; Lee, J.W. The bovine neutrophil: Structure and function in blood and milk. Vet. Res. 2003, 34, 597–627. [Google Scholar] [PubMed] [Green Version]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Aitken, S.L.; Corl, C.M.; Sordillo, L.M. Immunopathology of mastitis: Insights into disease recognition and resolution. J. Mammary Gland Biol. Neoplasia 2011, 16, 291–304. [Google Scholar] [CrossRef]
- Gilbert, F.B.; Cunha, P.; Jensen, K.; Glass, E.J.; Foucras, G.; Robert-Granié, C.; Rupp, R.; Rainard, P. Differential response of bovine mammary epithelial cells to Staphylococcus aureus or Escherichia coli agonists of the innate immune system. Vet. Res. 2013, 44, 40. [Google Scholar] [CrossRef] [Green Version]
- Strandberg, Y.; Gray, C.; Vuocolo, T.; Donaldson, L.; Broadway, M.; Tellam, R. Lipopolysaccharide and lipoteichoic acid induce different innate immune responses in bovine mammary epithelial cells. Cytokine 2005, 31, 72–86. [Google Scholar] [CrossRef]
- Günther, J.; Koczan, D.; Yang, W.; Nurnberg, G.; Repsilber, D.; Schuberth, H.J.; Park, Z.; Maqbool, N.; Molenaar, A.; Seyfert, H.M. Assessment of the immune capacity of mammary epithelial cells: Comparison with mammary tissue after challenge with Escherichia coli. Vet. Res. 2009, 40, 31. [Google Scholar] [CrossRef] [Green Version]
- Griesbeck-Zilch, B.; Meyer, H.H.; Kuhn, C.H.; Schwerin, M.; Wellnitz, O. Staphylococcus aureus and Escherichia coli cause deviating expression profiles of cytokines and lactoferrin messenger ribonucleic acid in mammary epithelial cells. J. Dairy Sci. 2008, 91, 2215–2224. [Google Scholar] [CrossRef] [Green Version]
- Genini, S.; Badaoui, B.; Sclep, G.; Bishop, S.C.; Waddington, D.; van der Pinard, L.M.H.; Klopp, C.; Cabau, C.; Seyfert, H.M.; Petzl, W.; et al. Strengthening insights into host responses to mastitis infection in ruminants by combining heterogeneous microarray data sources. BMC Genom. 2011, 12, 225. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xiu, L.; Hu, Q.; Cui, X.; Liu, B.; Tao, L.; Wang, T.; Wu, J.; Chen, Y.; Chen, Y. Deep Sequencing-Based Transcriptional Analysis of Bovine Mammary Epithelial Cells Gene Expression in Response to In Vitro Infection with Staphylococcus aureus Stains. PLoS ONE 2013, 8, e82117. [Google Scholar] [CrossRef]
- McGrath, M.F. A novel system for mammary epithelial cell culture. J. Dairy Sci. 1987, 70, 1967–1980. [Google Scholar] [CrossRef]
- Rose, T.M.; Aso, H.; Yonekura, S.; Komatsu, T.; Hagino, A.; Ozutsumi, K.; Obara, Y. In Vitro differentiation of a cloned bovine mammary epithelial cell. J. Dairy Res. 2002, 69, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, V.; Constant, S.; Eisenmesser, E.; Bukrinsky, M. Cyclophilin-CD147 interactions: A new target for anti-inflammatory therapeutics. Clin. Exp. Immunol. 2010, 160, 305–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigro, P.; Pompilio, G.; Capogrossi, M.C. Cyclophilin A: A key player for human disease. Cell Death Dis. 2013, 4, e888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handschumacher, R.E.; Harding, M.W.; Rice, J.; Drugge, R.J.; Speicher, D.W. Cyclophilin: A specific cytosolic binding protein for cyclosporin A. Science 1984, 226, 544–547. [Google Scholar] [CrossRef]
- Takanashi, S.; Nochi, T.; Abe, M.; Itaya, N.; Urakawa, M.; Sato, K.; Zhuang, T.; Umemura, S.; Hayashi, T.; Kiku, Y.; et al. Extracellular cyclophilin A possesses chemotaxic activity in cattle. Vet. Res. 2015, 46, 80. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.Y.; Wang, H.; Qi, S.; Wang, X.; Li, X.; Zhou, K.; Zhang, Y.; Gao, M.Q. CYP1A1 Relieves Lipopolysaccharide-Induced Inflammatory Responses in Bovine Mammary Epithelial Cells. Mediat. Inflamm. 2018, 2018, 4093285. [Google Scholar] [CrossRef]
- McClenahan, D.J.; Sotos, J.P.; Czuprynski, C.J. Cytokine response of bovine mammary gland epithelial cells to Escherichia coli, coliform culture filtrate, or lipopolysaccharide. Am. J. Vet. Res. 2005, 66, 1590–1597. [Google Scholar] [CrossRef]
- Petzl, W.; Zerbe, H.; Günther, J.; Yang, W.; Seyfert, H.M.; Nürnberg, G.; Schuberth, H.J. Escherichia coli, but not Staphylococcus aureus, triggers an early increased expression of factors contributing to the innate immune defense in the udder of the cow. Vet. Res. 2008, 39, 18. [Google Scholar] [CrossRef] [Green Version]
- Günther, J.; Esch, K.; Poschadel, N.; Petzl, W.; Zerbe, H.; Mitterhuemer, S.; Blum, H.; Seyfert, H.M. Comparative kinetics of Escherichia coli-and Staphylococcus aureus-specific activation of key immune pathways in mammary epithelial cells demonstrates that S. aureus elicits a delayed response dominated by interleukin-6 (IL-6) but not by IL-1A or tumor necrosis factor alpha. Infect. Immun. 2011, 79, 695–707. [Google Scholar]
- Brand, B.; Hartmann, A.; Repsilber, D.; Griesbeck-Zilch, B.; Wellnitz, O.; Kuhn, C.; Ponsuksili, S.; Meyer, H.H.; Schwerin, M. Comparative expression profiling of E. coli and S. aureus inoculated primary mammary gland cells sampled from cows with different genetic predispositions for somatic cell score. Genet. Sel. Evol. 2011, 43, 24. [Google Scholar] [CrossRef] [Green Version]
- Rainard, P.; Riollet, C. Innate immunity of the bovine mammary gland. Vet. Res. 2006, 37, 369–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellnitz, O.; Bruckmaier, R.M. The innate immune response of the bovine mammary gland to bacterial infection. Vet. J. 2012, 192, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Waller, K.P. Modulation of endotoxin-induced inflammation in the bovine teat using antagonists/inhibitors to leukotrienes, platelet activating factor and interleukin 1 beta. Vet. Immunol. Immunopathol. 1997, 57, 239–251. [Google Scholar] [CrossRef]
- Griesbeck-Zilch, B.; Osman, M.; Kühn, C.; Schwerin, M.; Bruckmaier, R.H.; Pfaffl, M.W.; Hammerle-Fickinger, A.; Meyer, H.H.; Wellnitz, O. Analysis of key molecules of the innate immune system in mammary epithelial cells isolated from marker-assisted and conventionally selected cattle. J. Dairy Sci. 2009, 92, 4621–4633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinard, P.; Riollet, C.; Berthon, P.; Cunha, P.; Fromageau, A.; Rossingnol, C.; Gilbert, F.B. The chemokine CXCL3 is responsible for the constitutive chemotactic activity of bovine milk for neutrophils. Mol. Immunol. 2008, 45, 4020–4027. [Google Scholar] [CrossRef] [PubMed]
- Struyf, S.; Gouwy, M.; Dillen, C.; Proost, P.; Opdenakker, G.; Van Damme, J. Chemokines synergize in the recruitment of circulating neutrophils into inflamed tissue. Eur. J. Immunol. 2005, 35, 1583–1591. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Q.; Hoover, D.M.; Staley, P.; Tucker, K.D.; Lubkowski, J.; Oppenheim, J.J. Many chemokines including CCL20/MIP-3alpha display antimicrobial activity. J. Leukoc. Biol. 2003, 74, 448–455. [Google Scholar] [CrossRef]
- Liew, F.Y.; Xu, D.; Brint, E.K.; O’Neill, L.A. Negative regulation of toll-like receptor-mediated immune responses. Nat. Rev. Immunol. 2005, 5, 446–458. [Google Scholar] [CrossRef]
- Liu, M.; Song, S.; Li, H.; Jiang, X.; Yin, P.; Wan, C.; Liu, X.; Liu, F.; Xu, J. The protective effect of caffeic acid against inflammation injury of primary bovine mammary epithelial cells induced by lipopolysaccharide. J. Dairy Sci. 2014, 97, 2856–2865. [Google Scholar] [CrossRef] [Green Version]
- Bannerman, D.D.; Paape, M.J.; Lee, J.W.; Zhao, X.; Hope, J.C.; Rainard, P. Escherichia coli and Staphylococcus aureus elicit differential innate immune responses following intramammary infection. Clin. Diagn. Lab. Immunol. 2004, 11, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Schröder, N.W.; Morath, S.; Alexander, C.; Hamann, L.; Hartung, T.; Zahringer, U.; Gobel, U.B.; Weber, J.R.; Schumann, R.R. Lipoteichoic acid (Pam3CSK4) of Streptococcus pneumoniae and Staphylococcus aureus activates immune cells via Toll-like receptor (TLR)-2, lipopolysaccharide binding protein (LBP), and CD14, whereas TLR-4 and MD-2 are not involved. J. Biol. Chem. 2003, 278, 15587–15594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.W.; Bannerman, D.D.; Paape, M.J.; Huang, M.K.; Zhao, X. Characterization of cytokine expression in milk somatic cells during intramammary infections with Escherichia coli or Staphylococcus aureus by real-time PCR. Vet. Res. 2006, 37, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, K.; Nigro, P.; Berk, B.C. Oxidative stress and vascular smooth muscle cell growth: A mechanistic linkage by cyclophilin A. Antioxid. Redox Signal. 2010, 12, 675–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, K.; Nigro, P.; Matoba, T.; O’Dell, M.R.; Cui, Z.; Shi, X.; Mohan, A.; Yan, C.; Abe, J.; Illig, K.A.; et al. Cyclophilin A enhances vascular oxidative stress and the development of angiotensin II-induced aortic aneurysms. Nat. Med. 2009, 15, 649–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tohno, M.; Shimosato, T.; Kawai, Y.; Ikegami, S.; Taketomo, N.; Saito, T.; Kitazawa, H. Advanced molecular immunoassay system for immunobiotic lactic acid bacteria using a transfectant of Toll-like receptor 2. Anim. Sci. J. 2007, 78, 195–205. [Google Scholar] [CrossRef]
- Mudunuri, U.; Che, A.; Yi, M.; Stephens, R.M. bioDBnet: The biological database network. Bioinformatics 2009, 25, 555–556. [Google Scholar] [CrossRef] [Green Version]
- Elsik, C.G.; Unni, D.R.; Diesh, C.M.; Tayal, A.; Emery, M.L.; Nguyen, H.N.; Hagen, D.E. Bovine Genome Database: New tools for gleaning function from the Bos taurus genome. Nucleic Acids Res. 2016, 44, D834–D839. [Google Scholar] [CrossRef] [Green Version]
- Breuer, K.; Foroushani, A.K.; Laird, M.R.; Chen, C.; Sribnaia, A.; Lo, R.; Winsor, G.L.; Hancock, R.E.; Brinkman, F.S.; Lynn, D.J. InnateDB: Systems biology of innate immunity and beyond--recent updates and continuing curation. Nucleic Acids Res. 2013, 41, 24. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control vs. LPS | Control vs. S. aureus JE2 | Control vs. S. aureus SA003 | |
|---|---|---|---|
| Upregulated | 222 | 227 | 226 |
| Downregulated | 207 | 238 | 294 |
| Total | 447 | 465 | 520 |
| GO Terms and IDs | Genes Involved | p-Value |
|---|---|---|
| LPS stimulation: | ||
| Cell chemotaxis [GO:0060326] | AGTR1, CCL2, CXCL2, CXCL3, CXCL5, CXCL8, CXCL9 | 0.0001 |
| Inflammatory response [GO:0006954] | CCL20, CCL2, CXCL2, CXCL8, CXCL9, GGT5, IL1α, IL1β | 0.002 |
| Positive regulation of nitric-oxide synthetase production [GO:0051770] | CCL20, CCL2, NOD2 | 0.0002 |
| Response molecules of bacterial origin [GO:0002237] | CXCL2, CXCL8 | 0.003 |
| Negative regulation of extrinsic apoptotic signaling pathway [GO:2001240] | IL1α, IL1β, IL17, UNC5B | 0.006 |
| S. aureus JE2 stimulation: | ||
| Cell differentiation [GO:0030154] | CCDC135, DLL1, HEMGN, MGP, NEUROD6, NKX2-2, PDX1, PIWIL3, SEMA4G, TCF23 | 0.001 |
| Cell adhesion [GO:0007155] | ABL1, CLDN11, ITGBL1, MAG, MPDZ, NCAN, PRKCE | 0.02 |
| Inflammatory response [GO:0006954] | CCL26, CNR2, CRP, CXCR2, IL22 | 0.04 |
| Blood coagulation [GO:0007596] | ABL1, EFEMP2, ESAM, KIF2B, MAG, PRKCE, PRKG2, SERPINB2 | 0.08 |
| Chemotaxis [GO:0006935] | CCL26, CXCR2, CXCR5 | 0.19 |
| S. aureus SA003 stimulation: | ||
| Innate immune response [GO:0045087] | AHSG, APOA1, C8A, F11, FREM1, KLRD1, MAFB, MAPK10, OTUB1, PRKCE, SFTPA1 | 0.01 |
| Cell surface receptor signaling pathway [GO:0007166] | GPR97, KLRD1, LEPR, LIFR | 0.03 |
| Blood coagulation [GO:0007596] | APOA1, ESAM, F11, KIF3C, PRKCE, TRPC3 | 0.03 |
| Signal transduction [GO:0007165] | CAPN3, ECM1, GDF3, MAPK10, NPAS2, PDE4B, PRKCE, SYNGAP1 | 0.04 |
| Positive regulation of NF-kB signaling [GO:0043123] | ECM1, HTR2B, PRKCE | 0.06 |
| Gene Symbol | Gene Name | LPS | S. aureus JE2 | S. aureus SA003 |
|---|---|---|---|---|
| IL1α | Interleukin 1 Alpha | 1.463 | N.M | N.M |
| IL1β | Interleukin 1 Beta | 2.94 | N.M | N.M |
| CCL2 | C-C Motif Chemokine Ligand 2 (also called monocyte chemoattractant protein -1, MCP-1) | 1.682 | N.M | N.M |
| CX3CL1 | C-X3-C Motif Chemokine Ligand 1 | 1.146 | N.M | N.M |
| CXCL2 | C-X-C Motif Chemokine Ligand 2 | 2.953 | N.M | N.M |
| CXCL3 | C-X-C Motif Chemokine Ligand 3 | 1.194 | N.M | N.M |
| CXCL5 | C-X-C Motif Chemokine Ligand 5 | 2.007 | N.M | N.M |
| CXCL8 | C-X-C Motif Chemokine Ligand 8 (also called Interleukin 8, IL8) | 3.013 | N.M | N.M |
| CXCL9 | C-X-C Motif Chemokine Ligand 9 | 4.379 | N.M | N.M |
| C6 | Complement component 6 | 1.313 | N.M | N.M |
| CRP | C-Reactive Protein | N.M | 3.012 | N.M |
| M-SAA3.2 | Mammary Serum Amyloid A3.2 | 3.413 | N.M | N.M |
| SAA3 | Serum Amyloid A3 | 3.869 | N.M | N.M |
| PPIA | Peptidylprolyl Isomerase A (also called Cyclophilin A, CyPA) | N.M | N.M | N.M |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.A.; Takagi, M.; Fukuyama, K.; Komatsu, R.; Albarracin, L.; Nochi, T.; Suda, Y.; Ikeda-Ohtsubo, W.; Rutten, V.; Eden, W.v.; et al. Transcriptome Analysis of The Inflammatory Responses of Bovine Mammary Epithelial Cells: Exploring Immunomodulatory Target Genes for Bovine Mastitis. Pathogens 2020, 9, 200. https://doi.org/10.3390/pathogens9030200
Islam MA, Takagi M, Fukuyama K, Komatsu R, Albarracin L, Nochi T, Suda Y, Ikeda-Ohtsubo W, Rutten V, Eden Wv, et al. Transcriptome Analysis of The Inflammatory Responses of Bovine Mammary Epithelial Cells: Exploring Immunomodulatory Target Genes for Bovine Mastitis. Pathogens. 2020; 9(3):200. https://doi.org/10.3390/pathogens9030200
Chicago/Turabian StyleIslam, Md. Aminul, Michihiro Takagi, Kohtaro Fukuyama, Ryoya Komatsu, Leonardo Albarracin, Tomonori Nochi, Yoshihito Suda, Wakako Ikeda-Ohtsubo, Victor Rutten, Willem van Eden, and et al. 2020. "Transcriptome Analysis of The Inflammatory Responses of Bovine Mammary Epithelial Cells: Exploring Immunomodulatory Target Genes for Bovine Mastitis" Pathogens 9, no. 3: 200. https://doi.org/10.3390/pathogens9030200
APA StyleIslam, M. A., Takagi, M., Fukuyama, K., Komatsu, R., Albarracin, L., Nochi, T., Suda, Y., Ikeda-Ohtsubo, W., Rutten, V., Eden, W. v., Villena, J., Aso, H., & Kitazawa, H. (2020). Transcriptome Analysis of The Inflammatory Responses of Bovine Mammary Epithelial Cells: Exploring Immunomodulatory Target Genes for Bovine Mastitis. Pathogens, 9(3), 200. https://doi.org/10.3390/pathogens9030200