Witch Hazel Significantly Improves the Efficacy of Commercially Available Teat Dips

 ,
,
Abstract
:1. Introduction
2. Results
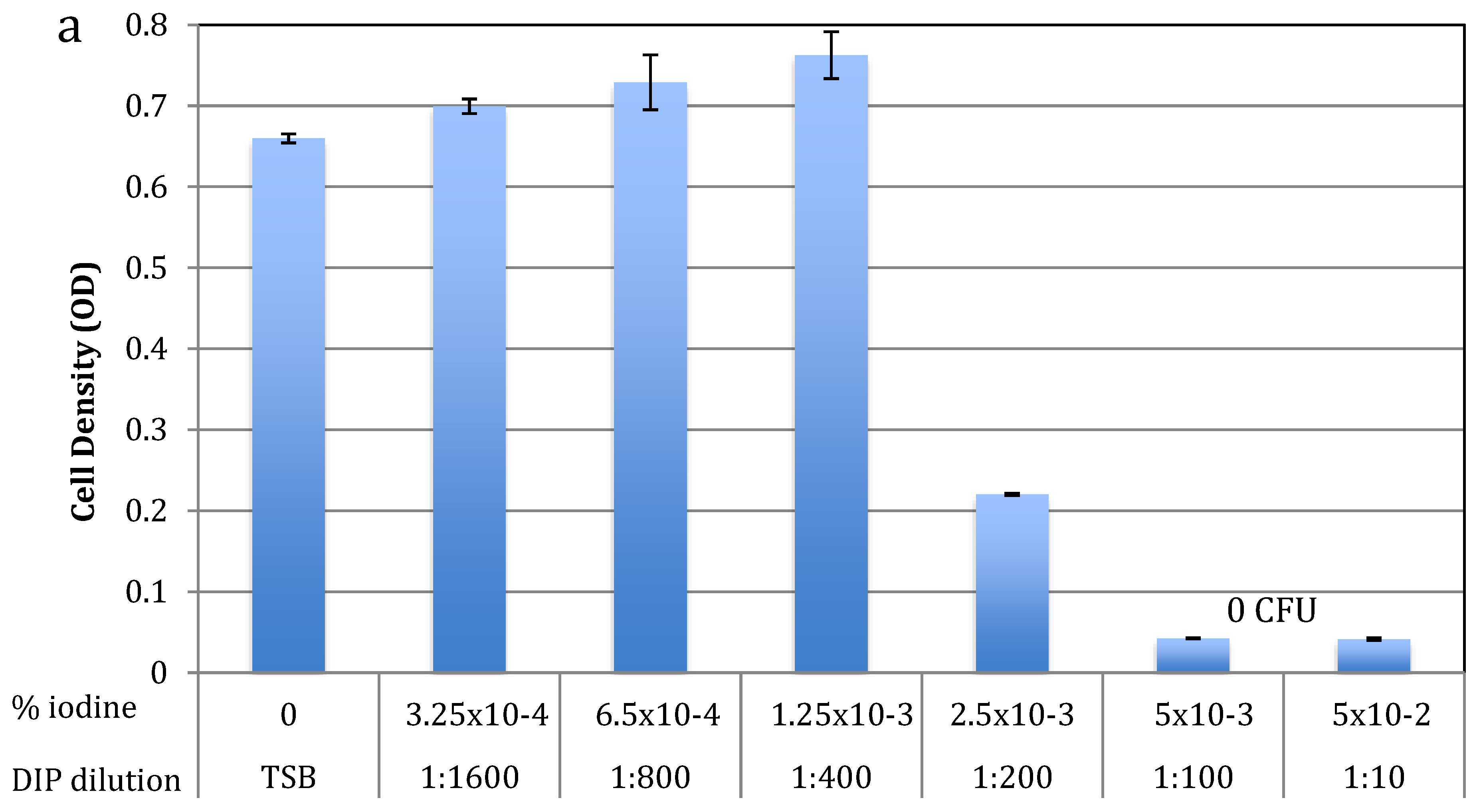
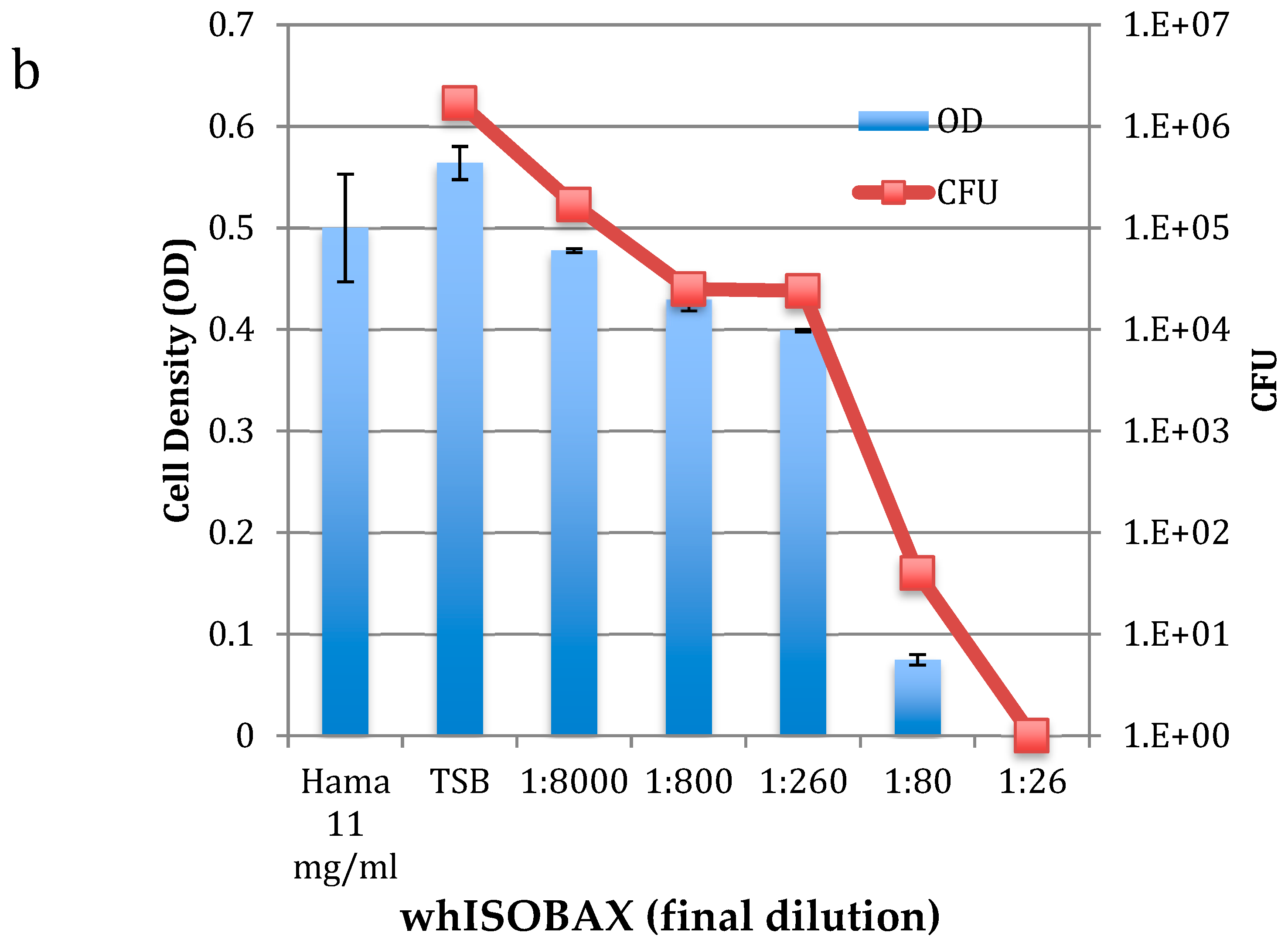
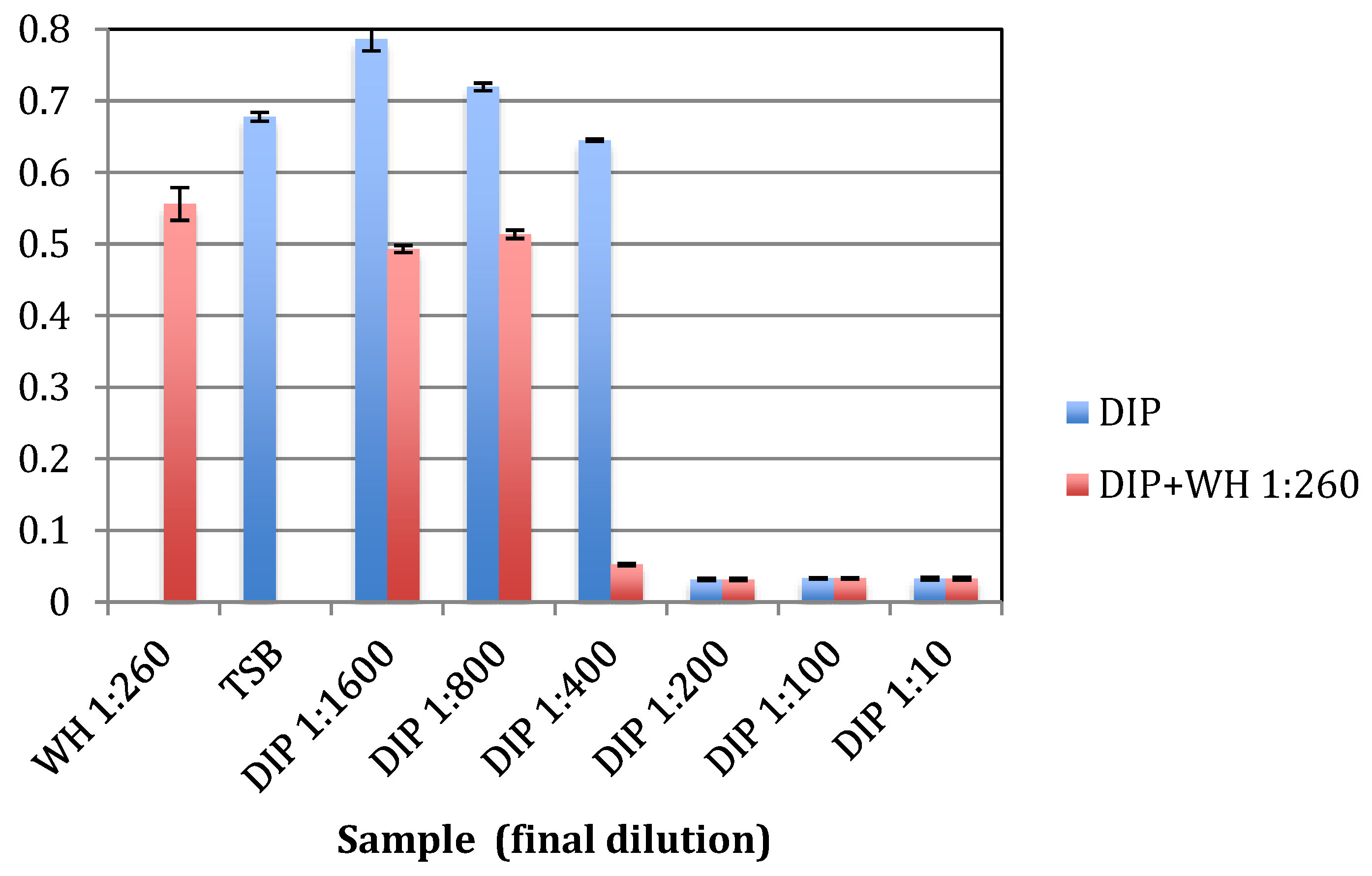
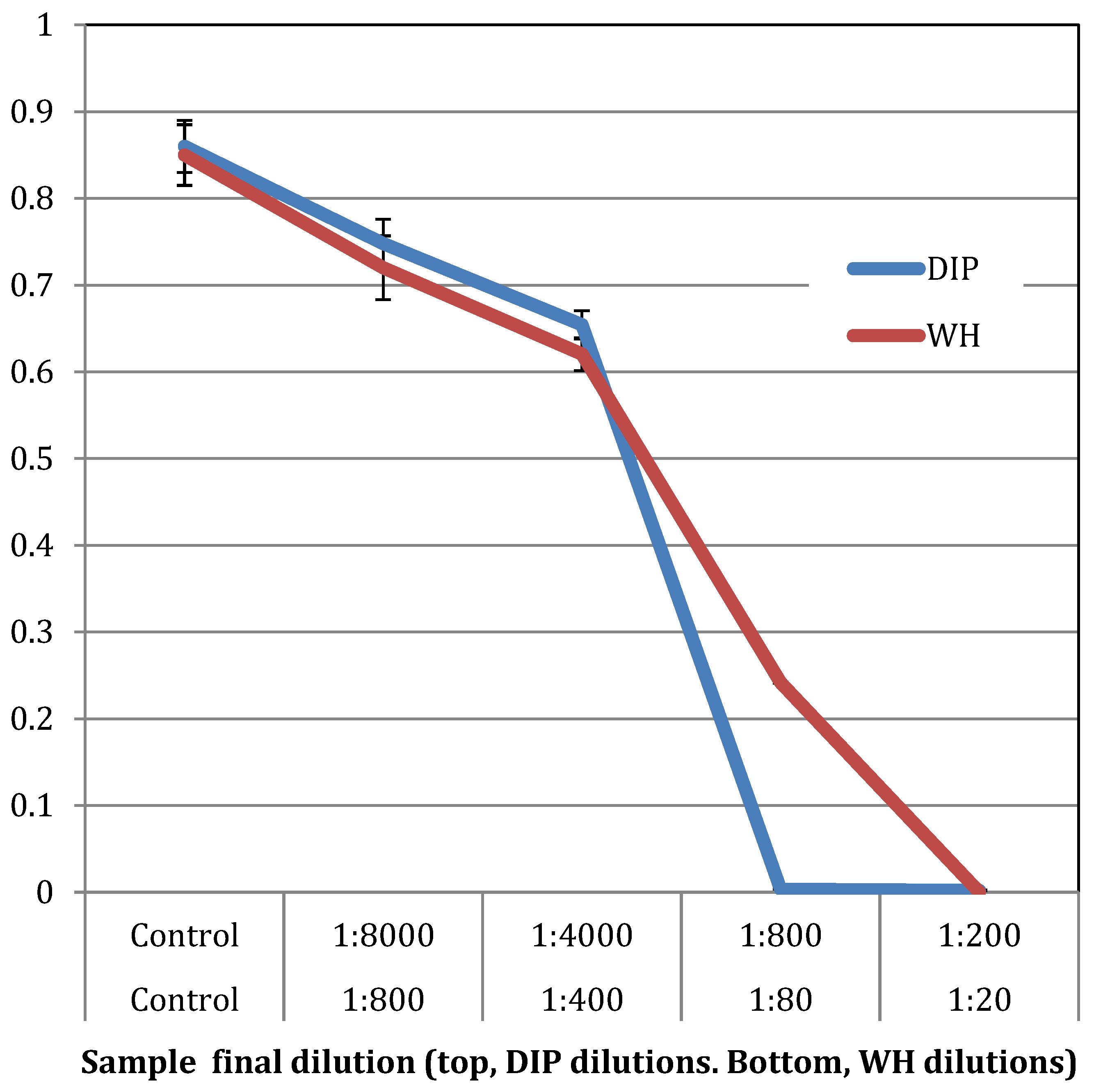
2.1. Antibacterial Activity against Planktonic S. epidermidis
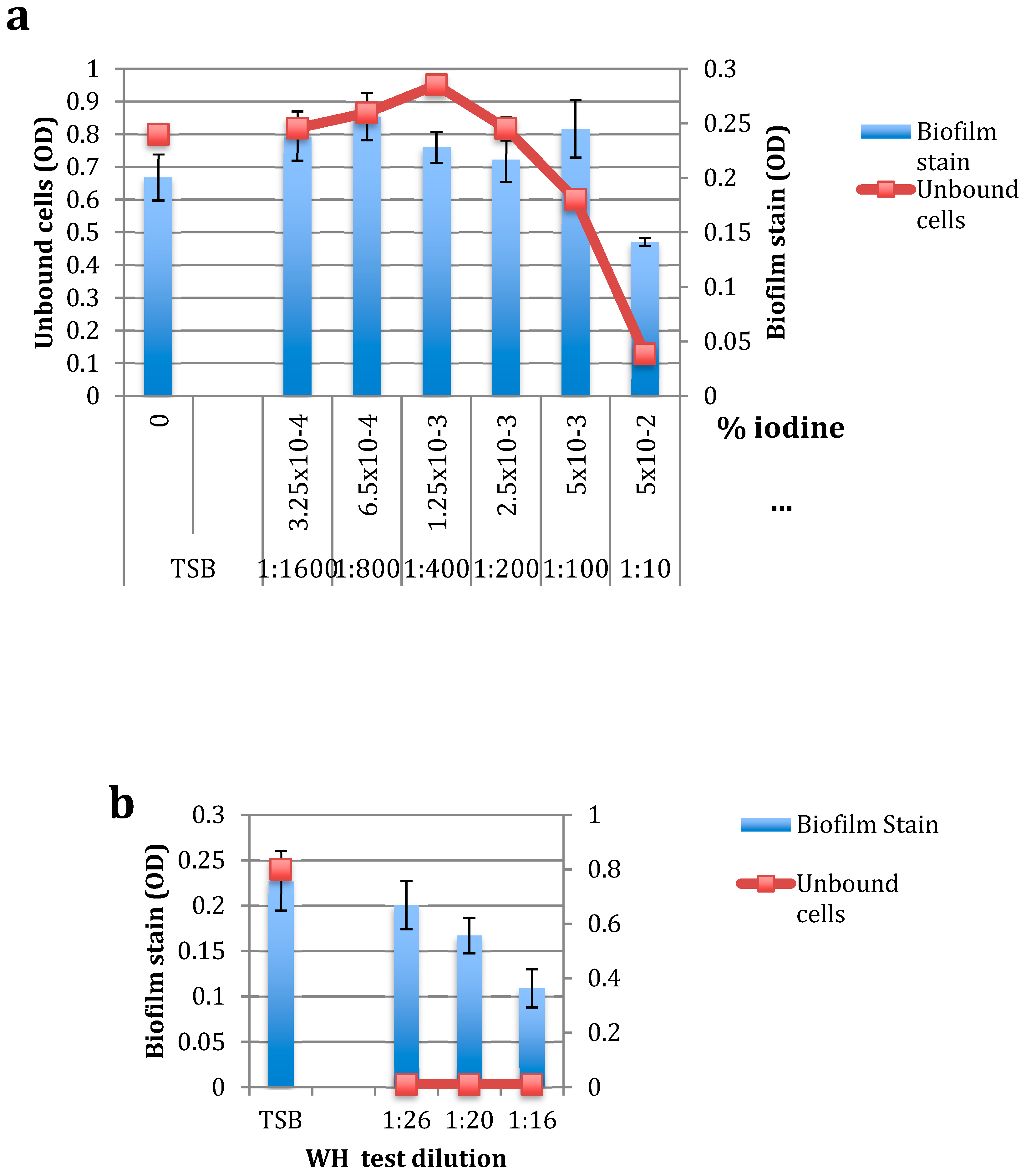
2.2. The Effect of DIP and WH on S. epidermidis Biofilm Formation
2.3. The Effect of DIP and WH on Pre-formed Biofilms
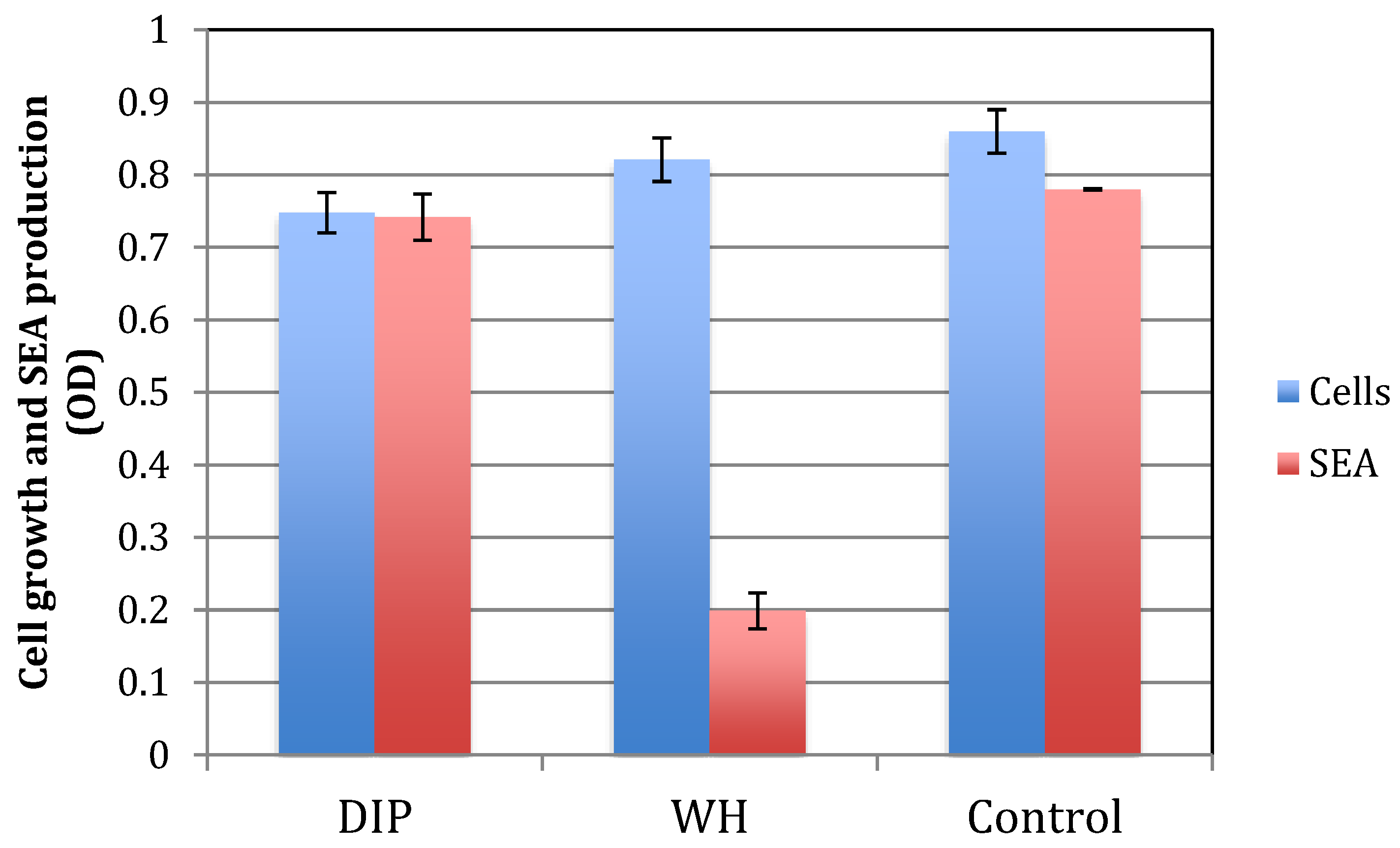
2.4. The Effect of DIP and WH on S. aureus Growth and Toxin Production
2.5. The Effect of DIP and WH on the Growth of Gram-negative Bacteria
3. Discussion
4. Materials and Methods
4.1. Bacteria
4.2. Test Formulations
4.3. MIC Testing on Planktonic Cells
4.4. MIC Testing on Biofilm Cells
4.5. Prevention of Biofilm Formation
4.6. SEA Production
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Progressive Dairy. U.S. Dairy Stats; Progressive Diary: Jerome, ID, USA, 2018. [Google Scholar]
- Halasa, T.; Huijps, K.; Osteras, O.; Hogeveen, H. Economic effects of bovine mastitis and mastitis management: A review. Vet. Q. 2007, 29, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Oliver, S.; Gillespie, B.; Susan, M.; Headrick, J.; Mark, B.; Lewis, M.; Henry, B.; Dowlen, H. Prevalence, Risk Factors, and Strategies for Controlling Mastitis in Heifers During the Periparturient Period. Int. J. Appl. Res. Vet. Med. 2005, 3, 150–162. [Google Scholar]
- Leitner, G.; Shoshani, E.; Krifucks, O.; Chaffer, M.; Saran, A. Milk leucocyte population patterns in bovine udder infection of different aetiology. J. Vet. Med. B Infect. Dis. Vet. Public Health 2000, 47, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Mastitis Control Program for Staph. aureus Infected Dairy Cows. Available online: https://milkquality.webhosting.cals.wisc.edu/wp-content/uploads/sites/212/2011/09/mastitis-control-program_staph.-aureaus.pdf (accessed on 1 March 2001).
- Chaffer, M.; Leitner, G.; Winkler, M.; Glickman, A.; Krifucks, O.; Ezra, E.; Saran, A. Coagulase-negative staphylococci and mammary gland infections in cows. Zentralbl. Veterinarmed. B 1999, 46, 707–712. [Google Scholar] [CrossRef]
- Piessens, V.; De Vliegher, S.; Verbist, B.; Braem, G.; Van Nuffel, A.; De Vuyst, L.; Heyndrickx, M.; Van Coillie, E. Characterization of coagulase-negative staphylococcus species from cows’ milk and environment based on bap, icaA, and mecA genes and phenotypic susceptibility to antimicrobials and teat dips. J. Dairy Sci. 2012, 95, 7027–7038. [Google Scholar] [CrossRef] [Green Version]
- Melchior, M.B.; Vaarkamp, H.; Fink-Gremmels, J. Biofilms: A role in recurrent mastitis infections? Vet. J. 2006, 171, 398–407. [Google Scholar] [CrossRef]
- Balaban, N.; Goldkorn, T.; Nhan, R.T.; Dang, L.B.; Scott, S.; Ridgley, R.M.; Rasooly, A.; Wright, S.C.; Larrick, J.W.; Rasooly, R.; et al. Autoinducer of virulence as a target for vaccine and therapy against Staphylococcus aureus. Science 1998, 280, 438–440. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Irie, Y.; Parsek, M.R. Quorum sensing and microbial biofilms. Curr. Top. Microbiol. Immunol. 2008, 322, 67–84. [Google Scholar]
- Nadell, C.D.; Xavier, J.B.; Levin, S.A.; Foster, K.R. The evolution of quorum sensing in bacterial biofilms. PLoS Biol. 2008, 6, e14. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Provan, G.J.; Helliwell, K. Determination of hamamelitannin, catechins and gallic acid in witch hazel bark, twig and leaf by HPLC. J. Pharm. Biomed. Anal. 2003, 33, 539–544. [Google Scholar] [CrossRef]
- Kiran, M.D.; Adikesavan, N.V.; Cirioni, O.; Giacometti, A.; Silvestri, C.; Scalise, G.; Ghiselli, R.; Saba, V.; Orlando, F.; Shoham, M.; et al. Discovery of a quorum-sensing inhibitor of drug-resistant staphylococcal infections by structure-based virtual screening. Mol. Pharmacol. 2008, 73, 1578–1586. [Google Scholar] [CrossRef] [Green Version]
- Brackman, G.; Breyne, K.; De Rycke, R.; Vermote, A.; Van Nieuwerburgh, F.; Meyer, E.; Van Calenbergh, S.; Coenye, T. The Quorum Sensing Inhibitor Hamamelitannin Increases Antibiotic Susceptibility of Staphylococcus aureus Biofilms by Affecting Peptidoglycan Biosynthesis and eDNA Release. Sci. Rep. 2016, 6, 20321. [Google Scholar] [CrossRef]
- Brackman, G.; Cos, P.; Maes, L.; Nelis, H.J.; Coenye, T. Quorum sensing inhibitors increase the susceptibility of bacterial biofilms to antibiotics in vitro and in vivo. Antimicrob. Agent. Chemother. 2011, 55, 2655–2661. [Google Scholar] [CrossRef] [Green Version]
- Cobrado, L.; Silva-Dias, A.; Azevedo, M.M.; Pina-Vaz, C.; Rodrigues, A.G. In vivo antibiofilm effect of cerium, chitosan and hamamelitannin against usual agents of catheter-related bloodstream infections. J. Antimicrob. Chemother. 2013, 68, 126–130. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Tena, S.; Fernandez-Cachon, M.L.; Carreras, A.; Mateos-Martin, M.L.; Costoya, N.; Moyer, M.P.; Nunez, M.J.; Torres, J.L.; Cascante, M. Hamamelitannin from witch hazel (Hamamelis virginiana) displays specific cytotoxic activity against colon cancer cells. J. Nat. Prod. 2012, 75, 26–33. [Google Scholar] [CrossRef]
- Korting, H.C.; Schafer-Korting, M.; Hart, H.; Laux, P.; Schmid, M. Anti-inflammatory activity of hamamelis distillate applied topically to the skin. Influence of vehicle and dose. Eur. J. Clin. Pharmacol. 1993, 44, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Wolff, H.H.; Kieser, M. Hamamelis in children with skin disorders and skin injuries: Results of an observational study. Eur. J. Pediatr. 2007, 166, 943–948. [Google Scholar] [CrossRef]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef]
- Friedman, M.; Rasooly, R. Review of the inhibition of biological activities of food-related selected toxins by natural compounds. Toxins 2013, 5, 743–775. [Google Scholar] [CrossRef]
- Tshivhandekano, I.; Ntushelo, K.; Ngezimana, W.; Tshikalange, T.E.; Mudau, F.N. Chemical compositions and antimicrobial activities of Athrixia phylicoides DC. (bush tea), Monsonia burkeana (special tea) and synergistic effects of both combined herbal teas. Asian Pac. J. Trop. Med. 2014, 7, S448–S453. [Google Scholar] [CrossRef] [Green Version]
- Foret, C.J.; Corbellini, C.; Young, S.; Janowicz, P. Efficacy of two iodine teat dips based on reduction of naturally occurring new intramammary infections. J. Dairy. Sci. 2005, 88, 426–432. [Google Scholar] [CrossRef]
- Martins, C.; Pinheiro, E.S.C.; Gentilini, M.; Benavides, M.L.; Santos, M.V. Efficacy of a high free iodine barrier teat disinfectant for the prevention of naturally occurring new intramammary infections and clinical mastitis in dairy cows. J. Dairy. Sci. 2017, 100, 3930–3939. [Google Scholar] [CrossRef]
- Azizoglu, R.O.; Lyman, R.; Anderson, K.L. Bovine Staphylococcus aureus: Dose response to iodine and chlorhexidine and effect of iodine challenge on antibiotic susceptibility. J. Dairy. Sci. 2013, 96, 993–999. [Google Scholar] [CrossRef] [Green Version]
- Nickerson, S.C. Choosing the Best Teat Dip for Mastitis Control and Milk Quality. In Proceedings of the NMC-PDPW Milk Quality Conference, Louisiana State University Agricultural Center, Homer, LA, USA, 9 February 2017; pp. 43–54. [Google Scholar]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef]
- Balaban, N.; Rasooly, A. Staphylococcal enterotoxins. Int. J. Food Microbiol. 2000, 61, 1–10. [Google Scholar] [CrossRef]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Rasooly, R.; Molnar, A.; Choi, H.Y.; Do, P.; Racicot, K.; Apostolidis, E. In–Vitro Inhibition of Staphylococcal Pathogenesis by Witch–Hazel and Green Tea Extracts. Antibiotics 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simoes, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug. Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Erskine, R.J. Mastitis in Cattle. In Merck Veterinary Manual; Merck & Co., Inc.: Kenilworth, NJ, USA, 2019. [Google Scholar]
- Dogan, B.; Klaessig, S.; Rishniw, M.; Almeida, R.A.; Oliver, S.P.; Simpson, K.; Schukken, Y.H. Adherent and invasive Escherichia coli are associated with persistent bovine mastitis. Vet. Microbiol. 2006, 116, 270–282. [Google Scholar] [CrossRef]
- Nuesch-Inderbinen, M.; Kappeli, N.; Morach, M.; Eicher, C.; Corti, S.; Stephan, R. Molecular types, virulence profiles and antimicrobial resistance of Escherichia coli causing bovine mastitis. Vet. Rec. Open 2019, 6, e000369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adkins, P.R.F.; Middleton, J.R.; Fox, L.K.; Pighetti, G.; Petersson-Wolfe, C.; Council, N.M. Laboratory Handbook on Bovine Mastitis; National Mastitis Council, Incorporated: New Prague, MN, USA, 2017. [Google Scholar]
- Pillai, S.K.; Moellering, R.C.; Eliopoulos, G.M. Antimicrobial Combinations. In Antibiotics in Laboratory Medicine; Lorian, V., Ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2005. [Google Scholar]
- Brandrick, M.; Newton, J.M.; Henderson, G.; Vickers, J.A. An investigation into the interaction between iodine and bacteria. J. Appl. Bacteriol. 1967, 30, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Sirk, T.W.; Brown, E.F.; Friedman, M.; Sum, A.K. Molecular binding of catechins to biomembranes: Relationship to biological activity. J. Agric. Food Chem. 2009, 57, 6720–6728. [Google Scholar] [CrossRef] [PubMed]
- Sirk, T.W.; Brown, E.F.; Sum, A.K.; Friedman, M. Molecular dynamics study on the biophysical interactions of seven green tea catechins with lipid bilayers of cell membranes. J. Agric. Food Chem. 2008, 56, 7750–7758. [Google Scholar] [CrossRef]
- Melchior, M.B.; Fink-Gremmels, J.; Gaastra, W. Extended antimicrobial susceptibility assay for Staphylococcus aureus isolates from bovine mastitis growing in biofilms. Vet. Microbiol. 2007, 125, 141–149. [Google Scholar] [CrossRef]
- Wolcott, R.; Lopez-Leban, F.; Kiran, M.; Balaban, N. Wound Healing by an Anti-Staphylococcal Biofilm Approach. In Biofilm Highlights; Hans-Curt, F., Wingender, J., Szewzyk, U., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 2011; pp. 141–161. [Google Scholar] [CrossRef]
- Leitner, G.; Krifucks, O.; Kiran, M.D.; Balaban, N. Vaccine development for the prevention of staphylococcal mastitis in dairy cows. Vet. Immunol. Immunopathol. 2011, 142, 25–35. [Google Scholar] [CrossRef]
- Gov, Y.; Borovok, I.; Korem, M.; Singh, V.K.; Jayaswal, R.K.; Wilkinson, B.J.; Rich, S.M.; Balaban, N. Quorum sensing in Staphylococci is regulated via phosphorylation of three conserved histidine residues. J. Biol. Chem. 2004, 279, 14665–14672. [Google Scholar] [CrossRef] [Green Version]
- Henrick, K.; Hirshberg, M. Structure of the signal transduction protein TRAP (target of RNAIII-activating protein). Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2012, 68, 744–750. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.S.; Je, J.Y. Gallic acid-grafted-chitosan inhibits foodborne pathogens by a membrane damage mechanism. J. Agric. Food Chem. 2013, 61, 6574–6579. [Google Scholar] [CrossRef]
- Gomes, F.; Saavedra, M.J.; Henriques, M. Bovine mastitis disease/pathogenicity: Evidence of the potential role of microbial biofilms. Pathog. Dis. 2016, 74, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Younis, A.; Krifucks, O.; Heller, E.D.; Samra, Z.; Glickman, A.; Saran, A.; Leitner, G. Staphylococcus aureus exosecretions and bovine mastitis. J. Vet. Med. B Infect Dis. Vet. Public Health 2003, 50, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirioni, O.; Giacometti, A.; Ghiselli, R.; Dell’Acqua, G.; Orlando, F.; Mocchegiani, F.; Silvestri, C.; Licci, A.; Saba, V.; Scalise, G.; et al. RNAIII-inhibiting peptide significantly reduces bacterial load and enhances the effect of antibiotics in the treatment of central venous catheter-associated Staphylococcus aureus infections. J. Infect Dis. 2006, 193, 180–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, H.; Wang, W.; Xu, L.; Ma, W.; Liu, L.; Wang, L.; Xu, C. Monoclonal antibody-based sandwich ELISA for the detection of staphylococcal enterotoxin A. Int. J. Environ. Res. Public Health 2013, 10, 1598–1608. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MIC DIP | MIC WH | MIC Combination (D-W) | FICI | MIC DIP | MIC WH | MIC Combination (D–W) | FICI |
|---|---|---|---|---|---|---|---|
| E. coli | P. aeruginosa | ||||||
| 1:80 | 1:10 | 1:160/1:160 | 0.562 | 1:160 | 1:20 | 1:320/1:160 | 0.625 |
| S. epidermidis | Planktonic | Planktonic | Biofilm | Biofilm |
|---|---|---|---|---|
| MIC Dilution | MBC Dilution | MIC Dilution | MBC Dilution | |
| whISOBAX | 1:80 | 1:26 | 1:26 | < 1:16 |
| DIP | 1:200 | 1:100 | 1:10 (up to 40% reduction of biofilm) | |
| DIP + WH 1:26 | >1:1600 | >1:1600 | 1:200 (up to 70% reduction of biofilm) | |
| DIP + whISOBAX 1:260 | 1:400 | 1:200 | ||
| whISOBAX + DIP 1:100 | 1:26 | < 1:16 |
| S. aureus | MIC | MBC | Inhibition of SEA Production (Max Dilution) |
|---|---|---|---|
| WH | 1:20 | 1:40 | 1:800 |
| DIP | 1:800 | 1:1600 | No activity |
| Final Tested Dilution of DIP | % Iodine in Final Dilution |
|---|---|
| 1:10 | >5 × 10−2 |
| 1:100 * | >5 × 10−3 |
| 1:200 ** | >2.5 × 10−3 |
| 1:400 | >1.25 × 10−3 |
| 1:800 | >6.5 × 10−4 |
| 1:1600 | >3.25 × 10−4 |
| Final Tested Dilution of WH | Final mg/mL GAE (Gallic Acid Equivalent) | Final mg/mL HAMA (Dry Weight Equivalent) |
|---|---|---|
| 1:16 | 0.79 | 1.08 |
| 1:20 | 0.633 | 0.865 |
| 1:26 ** | 0.48 | 0.665 |
| 1:40 | 0.31 | 0.435 |
| 1:80 * | 0.158 | 0.216 |
| 1:200 | 0.0633 | 0.0865 |
| 1:400 | 0.031 | 0.0435 |
| 1:800 | 0.0158 | 0.0216 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasooly, R.; Molnar, A.; Do, P.; Morroni, G.; Brescini, L.; Cirioni, O.; Giacometti, A.; Apostolidis, E. Witch Hazel Significantly Improves the Efficacy of Commercially Available Teat Dips. Pathogens 2020, 9, 92. https://doi.org/10.3390/pathogens9020092
Rasooly R, Molnar A, Do P, Morroni G, Brescini L, Cirioni O, Giacometti A, Apostolidis E. Witch Hazel Significantly Improves the Efficacy of Commercially Available Teat Dips. Pathogens. 2020; 9(2):92. https://doi.org/10.3390/pathogens9020092
Chicago/Turabian StyleRasooly, Reuven, Adel Molnar, Paula Do, Gianluca Morroni, Lucia Brescini, Oscar Cirioni, Andrea Giacometti, and Emmanouil Apostolidis. 2020. "Witch Hazel Significantly Improves the Efficacy of Commercially Available Teat Dips" Pathogens 9, no. 2: 92. https://doi.org/10.3390/pathogens9020092
APA StyleRasooly, R., Molnar, A., Do, P., Morroni, G., Brescini, L., Cirioni, O., Giacometti, A., & Apostolidis, E. (2020). Witch Hazel Significantly Improves the Efficacy of Commercially Available Teat Dips. Pathogens, 9(2), 92. https://doi.org/10.3390/pathogens9020092