Rapid Spread of Classical Swine Fever Virus among South Korean Wild Boars in Areas near the Border with North Korea

Abstract
:1. Introduction
2. Results
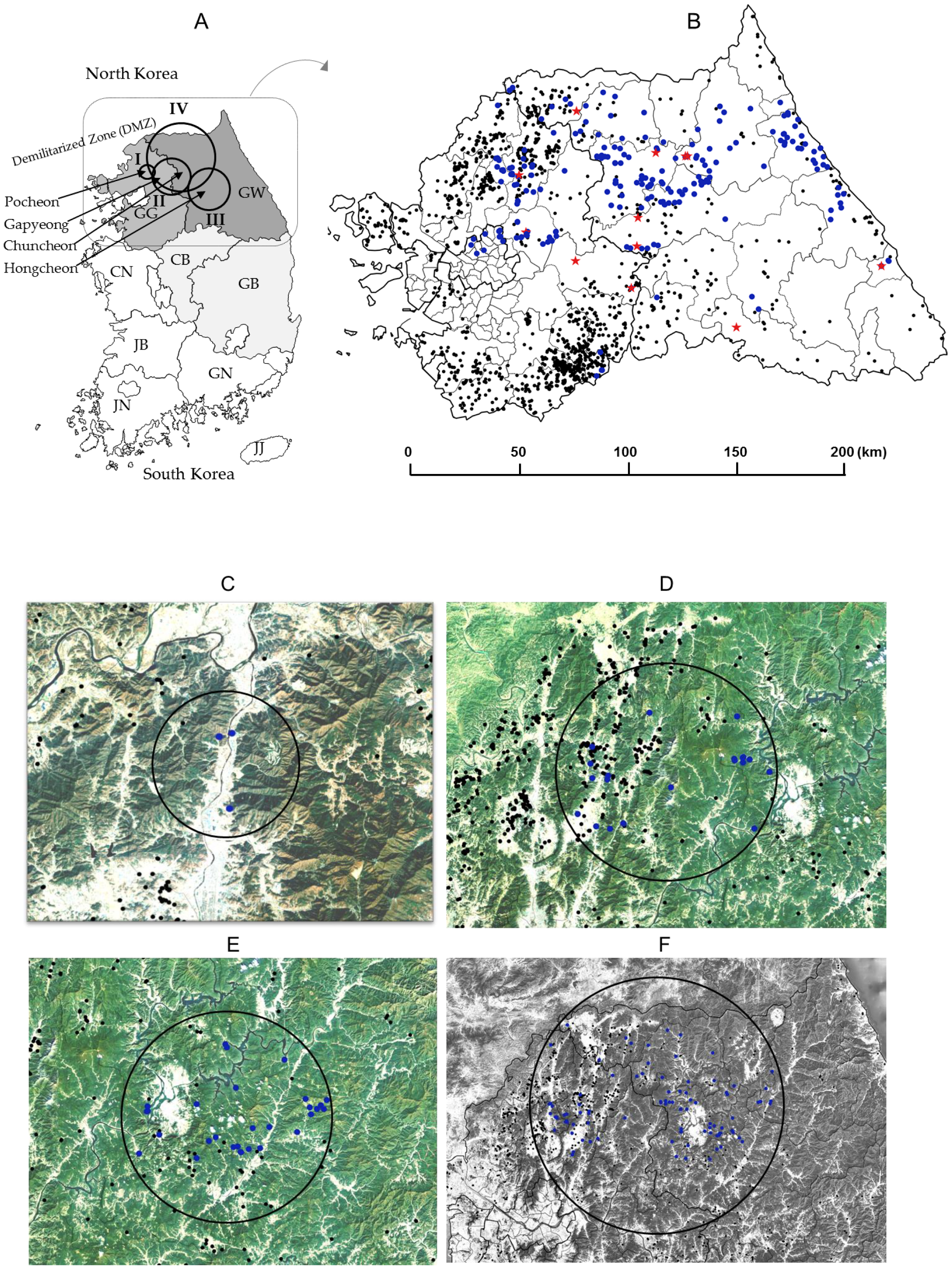
2.1. CSF Antibody Prevalence According to Province
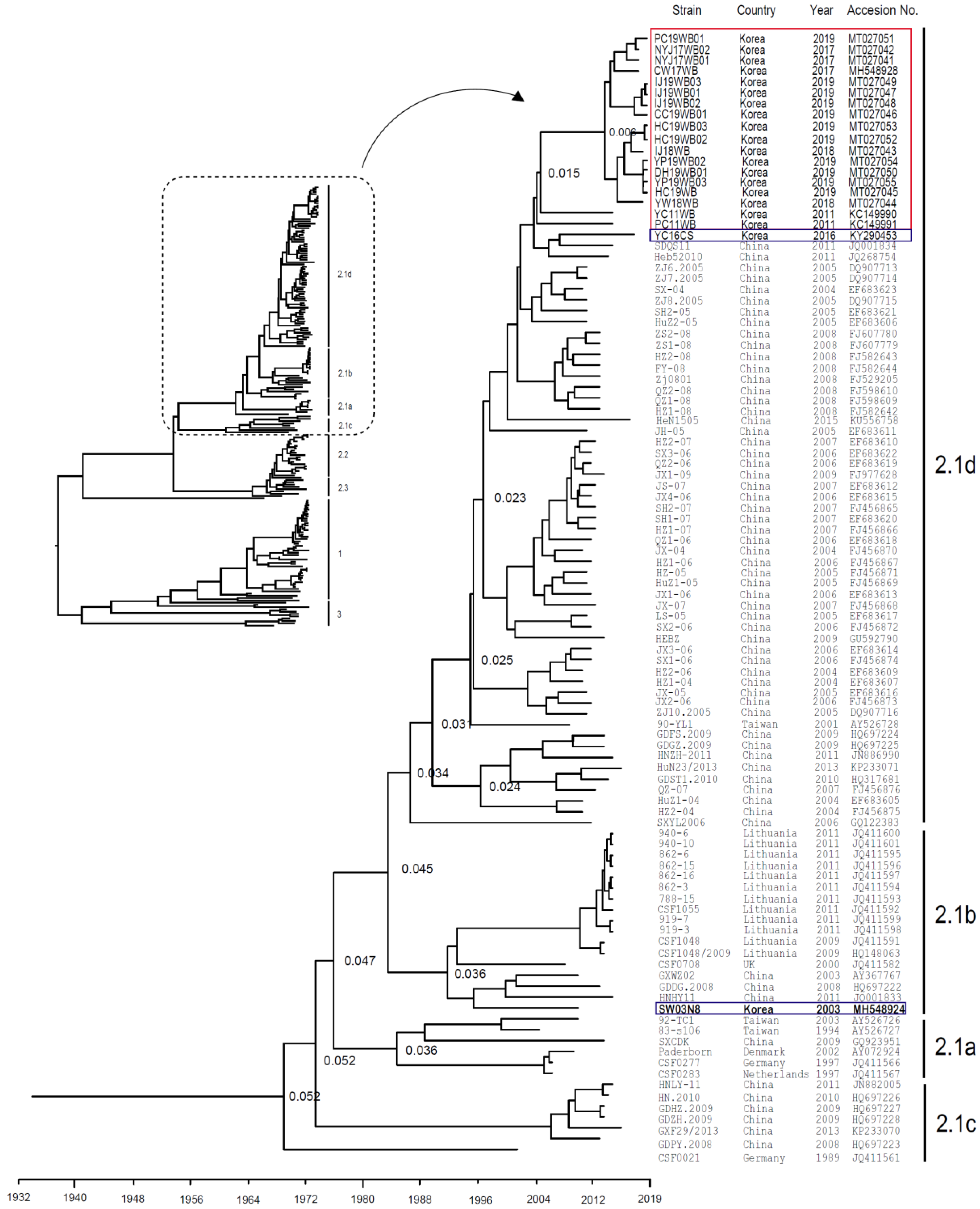
2.2. Genetic Analysis of CSFVs Isolated from Wild Boars
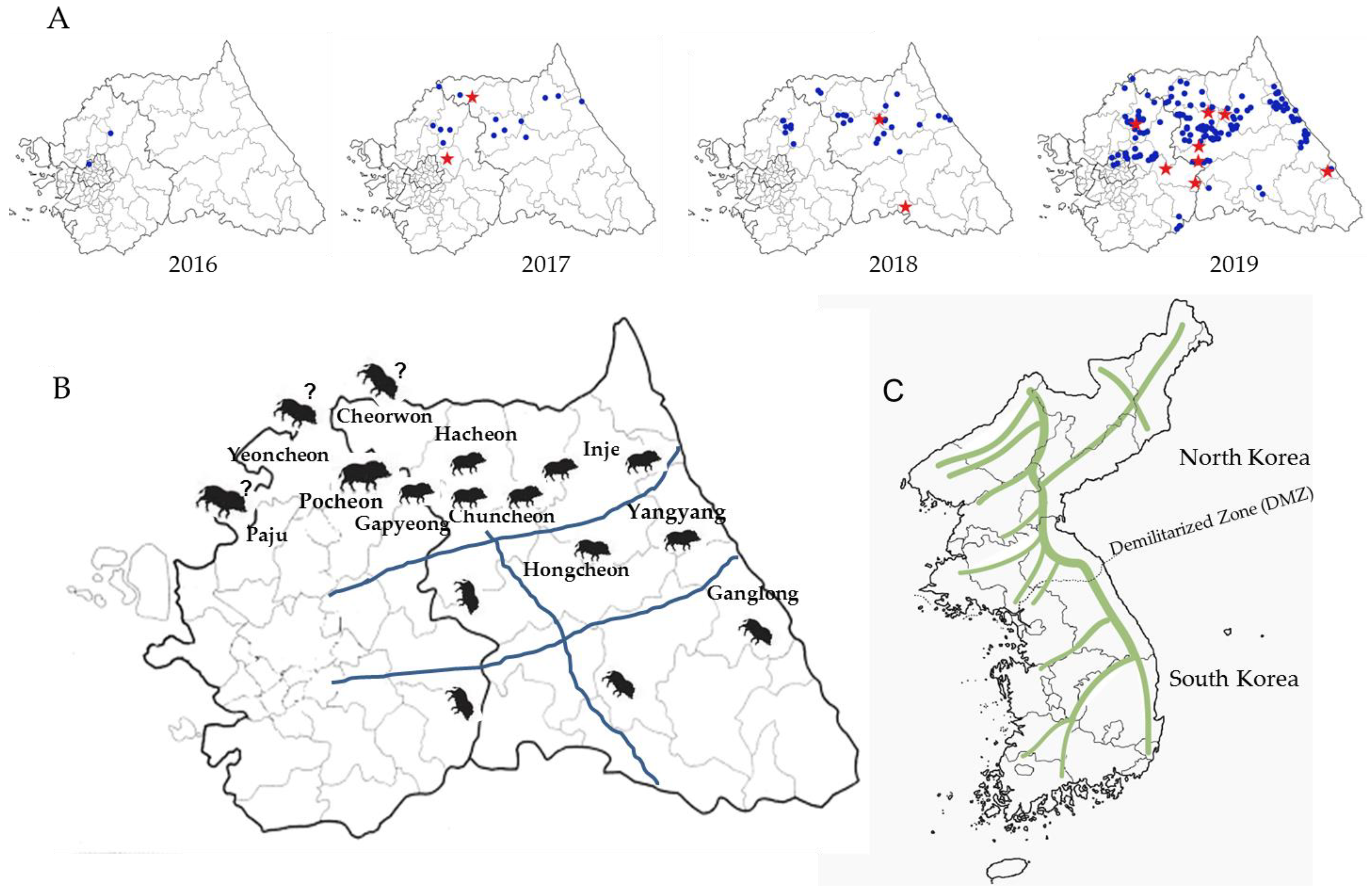
2.3. Space–Time Clusters
2.4. Relationship among CSF Sero-Positive and Age
3. Discussion
4. Materials and Methods
4.1. Sample Collection, RT-PCR, and Phylogenetic Analysis
4.2. Spatiotemporal Cluster Analysis
4.3. CSFV Ab B-ELISA and SN Tests
4.4. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Ethical Approval and Consent to Participate
References
- Edwards, S.; Fukusho, A.; Lefèvre, P.C.; Lipowski, A.; Pejsak, Z.; Roehe, P.; Westergaard, J. Classical swine fever: The global situation. Vet. Microbiol. 2000, 73, 103–119. [Google Scholar] [CrossRef]
- Moennig, V. The control of classical swine fever in wild boar. Front. Microbiol. 2015, 6, 1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritzemeier, J.; Teuffert, J.; Greiser-Wilke, I.; Staubach, C.; Schlüter, H.; Moennig, V. Epidemiology of classical swine fever in Germany in the 1990s. Vet. Microbiol. 2000, 77, 29–41. [Google Scholar] [CrossRef]
- Postel, A.; Moennig, V.; Becher, P. Classical swine fever in Europe—The current situation. Berl. Munch. Tierarztl. Wochenschr. 2013, 126, 468–475. [Google Scholar] [PubMed]
- Staubach, C.; Höreth-Böntgen, D.; Blome, S.; Fröhlich, A.; Blicke, J.; Jahn, B.; Teuffert, J.; Kramers, M. Descriptive summary of the classical swine fever control in wild boar in Germany since 2005. Berl. Munch. Tierarztl. Wochenschr. 2013, 126, 491–499. [Google Scholar] [PubMed]
- Zanardi, G.; Macchi, C.; Sacchi, C.; Rutili, D. Classical swine fever in wild boar in the Lombardy region of Italy from 1997 to 2002. Vet. Rec. 2003, 152, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Fromont, E.; Pontier, D.; Crucière, C.; Hars, J.; Barrat, J.; Pacholek, X.; Artois, M. Incidence and persistence of classical swine fever in free-ranging wild boar (Sus scrofa). Epidemiol. Infect. 2005, 133, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Kameyama, K.I.; Nishi, T.; Yamada, M.; Masujin, K.; Morioka, K.; Kokuho, T.; Fukai, K. Experimental infection of pigs with a classical swine fever virus isolated in Japan for the first time in 26 years. J. Vet. Med. Sci. 2019, 81, 1277–1284. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Jurado, C.; Bosch, J.; Ito, M.; Sanchez-Vizcaino, J.M.; Isoda, N.; Sakoda, A.Y. Role of Wild Boar in the Spread of Classical Swine Fever in Japan. Pathogens 2019, 8, 206. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.K.; Lim, S.I.; Kim, J.J.; Cho, Y.Y.; Song, J.Y.; Cho, I.S.; Hyun, B.H.; Choi, S.H.; Kim, S.H.; Park, E.H.; et al. Surveillance of classical swine fever in wild boar in South Korea from 2010–2014. J. Vet. Med. Sci. 2016, 77, 1667–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, D.J.; Lim, S.I.; Choe, S.; Kim, K.S.; Cha, R.M.; Cho, I.S.; Song, J.Y.; Hyun, B.H.; Park, B.K. Evolutionary dynamics of classical swine fever virus in South Korea: 1987–2017. Vet. Microbiol. 2018, 225, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Rho, P. Using habitat suitability model for the wild boar (Sus scrofa Linnaeus) to select wildlife passage sites in extensively disturbed temperate forests. J. Ecol. Environ. 2015, 38, 163–173. [Google Scholar] [CrossRef] [Green Version]
- 2018 Korean Wildlife Survey; Biological Resources Research Department, National Institute of Biological Resources: Incheon, Korea, 2018.
- Food and Agriculture Organization of the United Nations (FAO). ASF Situation in Asia Update 5 March 2020. Available online: http://www.fao.org/ag/againfo/programmes/en/empres/ASF/Situation_update.html (accessed on 5 March 2020).
- Schnyder, M.; Stärk, K.D.; Vanzetti, T.; Salman, M.D.; Thor, B.; Schleiss, W.; Griot, C. Epidemiology and control of an outbreak of classical swine fever in wild boar in Switzerland. Vet. Rec. 2002, 150, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Staubach, C.; Blome, S.; Guberti, V.; Thulke, H.H.; Vos, A.; Koenen, F.; LePotier, M.F. Controlling of CSFV in European wild boar using oral vaccination: A review. Front. Microbiol. 2015, 6, 1141. [Google Scholar] [CrossRef] [PubMed]
- Depner, K.; Gortazar, C.; Guberti, V.; Masiulis, M.; More, S.; O_l_sevskis, E.; Thulke, H.H.; Viltrop, A.; Wo_zniakowski, G.; Abrahantes, J.C.; et al. Epidemiological analyses of African swine fever in the Baltic States and Poland. EFSA J. 2017, 15, 5068. [Google Scholar]
- Lim, S.I.; Choe, S.; Kim, K.S.; Jeoung, H.Y.; Cha, R.M.; Park, G.S.; Shin, J.; Park, G.N.; Cho, I.S.; Song, J.Y.; et al. Assessment of the efficacy of an attenuated live marker classical swine fever vaccine (Flc-LOM-BErns) in pregnant sows. Vaccine 2019, 37, 3598–3604. [Google Scholar] [CrossRef] [PubMed]
- Paton, D.J.; McGoldrick, A.; Greiser-Wilke, I.; Parchariyanon, S.; Song, J.Y.; Liou, P.P.; Stadejek, T.; Lowings, J.P.; Björklund, H.; Belák, S. Genetic typing of classical swine fever virus. Vet. Microbiol. 2000, 73, 137–157. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigFree v.1.4.0. 2012. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 20 November 2019).
- Kulldorff, M.; Heffernan, R.; Hartman, J.; Assunção, R.; Mostashari, F. A space–time permutation scan statistic for disease outbreak detection. PLoS Med. 2005, 2, e59. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | No. of APa /No. of CWBb | Gender | Age (Months) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Male | Female | UKc | 0–12 | 13–24 | 25–36 | 37–48 | 48–70 | UKc | ||
| 2016 | 7/1683 | 2/584 | 1/417 | 4/682 | 0/369 | 3/499 | 0/88 | 0/26 | 0/19 | 4/682 |
| 2017 | 20/1670 | 9/912 | 11/757 | 0/1 | 3/630 | 16/762 | 0/171 | 0/70 | 1/36 | 0/1 |
| 2018 | 47/1320 | 28/740 | 19/580 | 0/0 | 14/479 | 23/608 | 4/138 | 6/64 | 0/31 | 0/0 |
| 2019 | 200/2297 | 84/1126 | 101/1045 | 15/126 | 34/909 | 94/902 | 32/186 | 16/95 | 4/33 | 20/172 |
| Total | 3.9 d (274/ 6970) | 3.6 (123/ 3362) | 4.7 (132/ 2799) | 2.3 (19/ 809) | 2.1 (51/ 2387) | 4.9 (136/ 2771) | 6.1 (36/ 583) | 8.6 (22/ 255) | 4.2 (5/ 119) | 2.8 (24/ 855) |
| Year | No. of APa /No. of CWBb | Percentage (%) for Region (No. of Antibody Positive/No. of Wild Boars Tested) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| GW | GG | GN | GB | JN | JB | CN | CB | JJ | UKc | ||
| 2016 | 7/1683 | 0.6 (1/148) | 1.6 (5/302) | 0 (0/334) | 0 (0/410) | 0 (0/71) | 0 (0/59) | 0.3 (1/261) | 0 (0/96) | 0 (0/2) | 0 (0/0) |
| 2017 | 20/1670 | 4.7 (12/251) | 4.6 (6/129) | 0.5 (1/195) | 0.3 (1/301) | 0 (0/189) | 0 (0/112) | 0 (0/221) | 0 (0/270) | 0 (0/0) | 0 (0/2) |
| 2018 | 47/1320 | 16.5d (33/200) | 9.2 (9/97) | 0.5 (1/196) | 0.8 (2/237) | 0 (0/150) | 0 (0/97) | 0 (0/172) | 1.3 (2/149) | 0 (0/22) | 0 (0/0) |
| 2019 | 200/2297 | 21.2e (129/608) | 14.3f (65/453) | 0.3 (1/292) | 0.3 (1/275) | 1.4 (1/69) | 0 (0/71) | 0.3 (1/289) | 0.9 (2/203) | 0 (0/37) | 0 (0/0) |
| Spatiotemporal Cluster Analysis of CSF Antibody Distribution | |||
|---|---|---|---|
| Cluster | I | II | III |
| Observed notifications | 5 | 22 | 34 |
| Expected notifications | 0.35 | 6.84 | 11.59 |
| Duration (days) | 181 | 60 | 61 |
| Start date | 30/11/2017 | 01/03/2019 | 30/12/2018 |
| End date | 29/05/2018 | 29/04/2019 | 28/02/2019 |
| Radius (km) | 3.87 | 23.05 | 24.49 |
| p-value | 0.038 | 0.0018 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choe, S.; Cha, R.M.; Yu, D.-S.; Kim, K.-S.; Song, S.; Choi, S.-H.; Jung, B.-I.; Lim, S.-I.; Hyun, B.-H.; Park, B.-K.; et al. Rapid Spread of Classical Swine Fever Virus among South Korean Wild Boars in Areas near the Border with North Korea. Pathogens 2020, 9, 244. https://doi.org/10.3390/pathogens9040244
Choe S, Cha RM, Yu D-S, Kim K-S, Song S, Choi S-H, Jung B-I, Lim S-I, Hyun B-H, Park B-K, et al. Rapid Spread of Classical Swine Fever Virus among South Korean Wild Boars in Areas near the Border with North Korea. Pathogens. 2020; 9(4):244. https://doi.org/10.3390/pathogens9040244
Chicago/Turabian StyleChoe, SeEun, Ra Mi Cha, Dae-Sung Yu, Ki-Sun Kim, Sok Song, Sung-Hyun Choi, Byung-Il Jung, Seong-In Lim, Bang-Hun Hyun, Bong-Kyun Park, and et al. 2020. "Rapid Spread of Classical Swine Fever Virus among South Korean Wild Boars in Areas near the Border with North Korea" Pathogens 9, no. 4: 244. https://doi.org/10.3390/pathogens9040244
APA StyleChoe, S., Cha, R. M., Yu, D. -S., Kim, K. -S., Song, S., Choi, S. -H., Jung, B. -I., Lim, S. -I., Hyun, B. -H., Park, B. -K., & An, D. -J. (2020). Rapid Spread of Classical Swine Fever Virus among South Korean Wild Boars in Areas near the Border with North Korea. Pathogens, 9(4), 244. https://doi.org/10.3390/pathogens9040244