
SARS-CoV-2 Evolution and Spike-Specific CD4+ T-Cell Response in Persistent COVID-19 with Severe HIV Immune Suppression

 , , , , , , , ,
, , , , , , , ,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Plasma RNA
2.1.1. SARS-CoV-2 RNA Extraction in Plasma
2.1.2. Droplet Digital Polymerase Chain Reaction (PCR) Analysis
2.2. SARS-CoV-2 Genomic Analysis from Nasopharyngeal Samples
2.2.1. Complementary DNA (cDNA) Synthesis and Multiplex Amplification
2.2.2. Library Construction and Genome Sequencing
2.2.3. Mutation Calling
2.3. T-Cell Response Analysis
2.3.1. Cell Stimulation
2.3.2. Immunophenotyping and Intracellular Cytokine Staining
3. Results
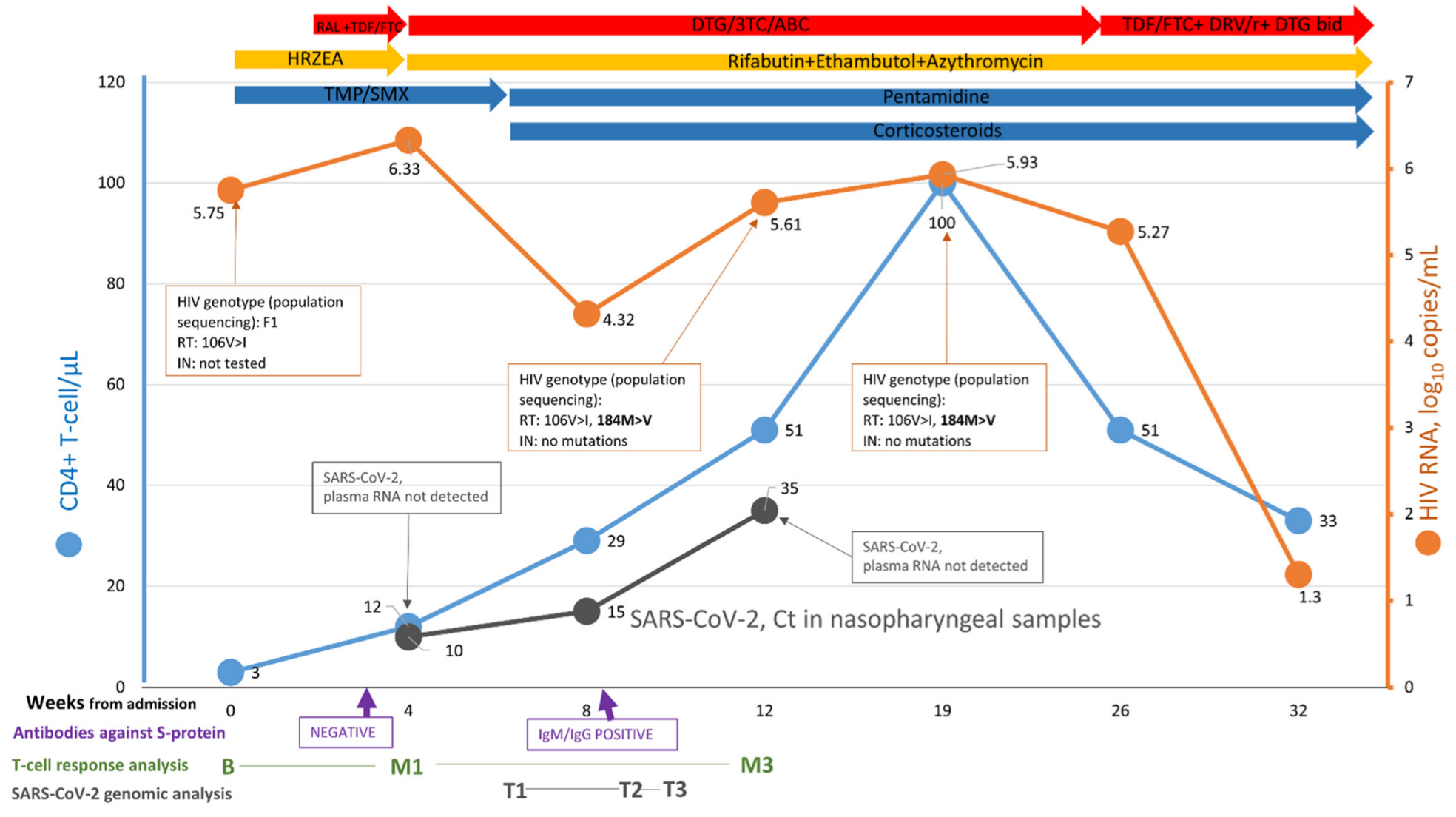
3.1. Case Report
3.2. Human Immunodeficiency Virus (HIV) Genomic Analysis
3.3. SARS-CoV-2 Plasma RNA
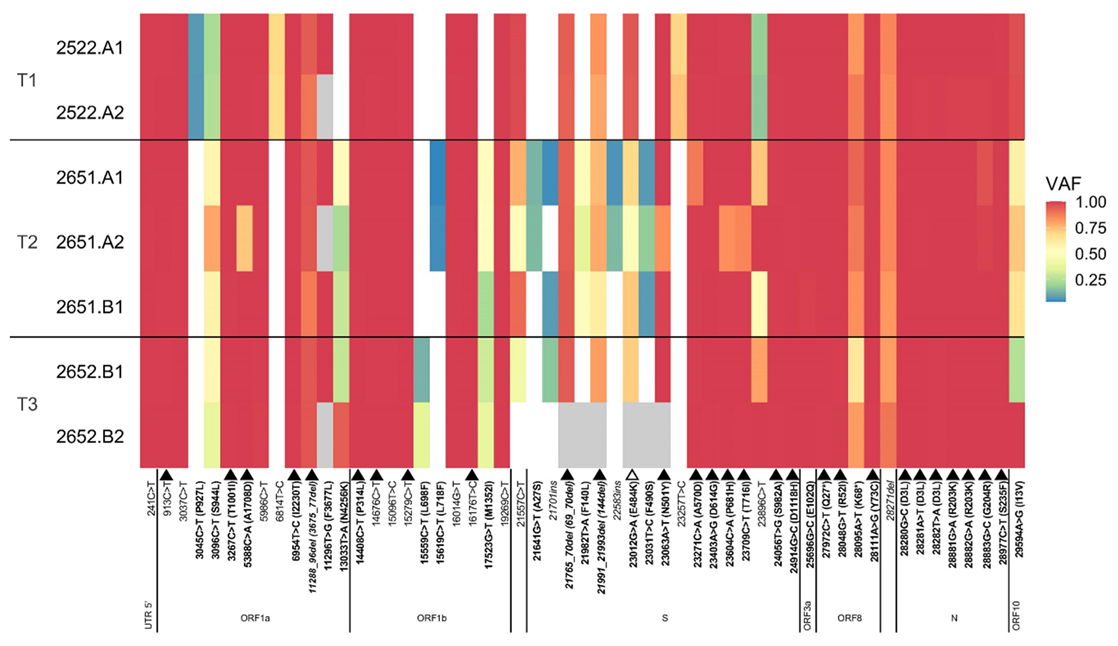
3.4. SARS-CoV-2 Genomic Analysis from Nasopharyngeal Samples
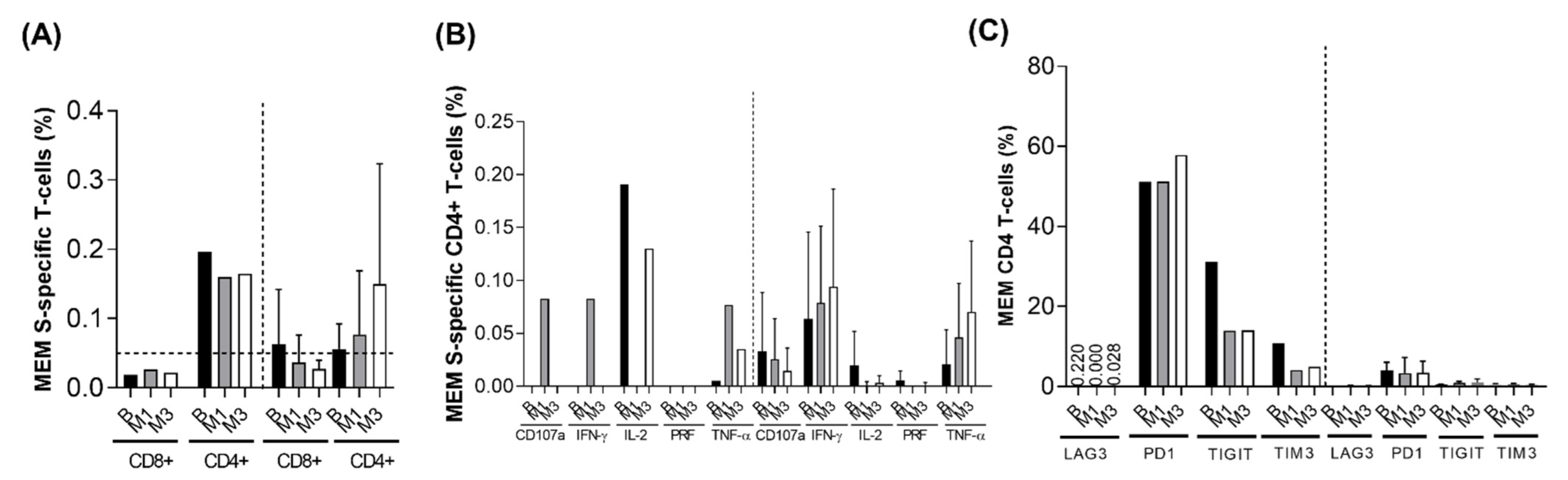
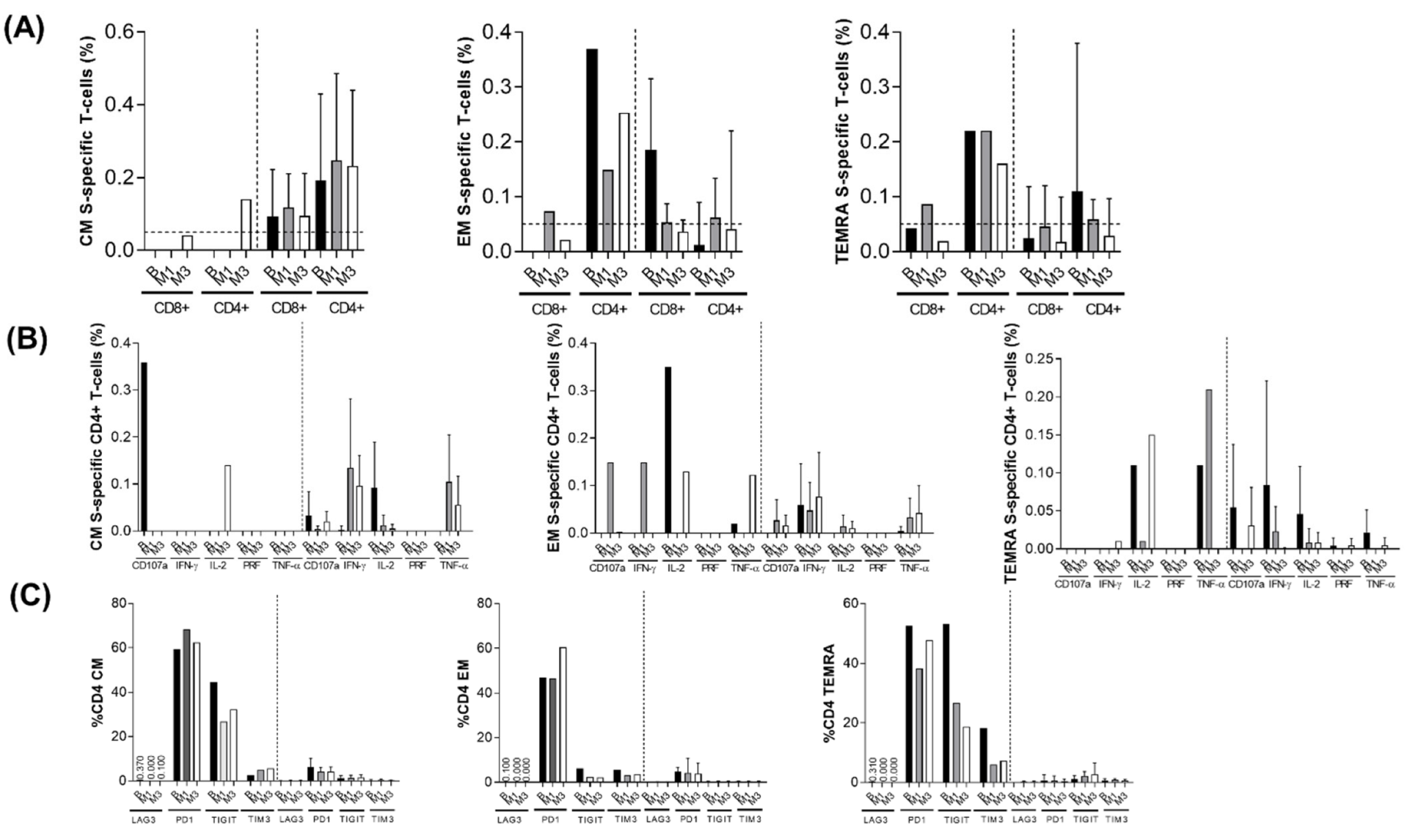
3.5. T-Cell Response Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karim, F.; Moosa, M.Y.S.; Gosnell, B.I.; Cele, S.; Giandhari, J.; Pillay, S.; Tegally, H.; Wilkinson, E.; San, J.E.; Msomi, N.; et al. Persistent SARS-CoV-2 infection and intra-host evolution in association with advanced HIV infection. MedRxiv, 2021; in press. [Google Scholar] [CrossRef]
- Quick, J.; Grubaugh, N.D.; Pullan, S.T.; Claro, I.M.; Smith, A.D.; Gangavarapu, K.; Oliveira, G.; Robles-Sikisaka, R.; Rogers, T.F.; Beutler, N.A.; et al. Multiplex PCR method for MinION and Illumina sequencing of Zika and other virus genomes directly from clinical samples. Nat. Protoc. 2017, 12, 1261–1276. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. Available online: https://arxiv.org/abs/1303.3997 (accessed on 8 August 2021).
- Grubaugh, N.D.; Gangavarapy, K.; Quick, J.; Matteson, N.L.; De Jesus, J.G.; Main, B.J.; Tan, A.L.; Paul, L.M.; Brackney, D.E.; Grewal, S.; et al. An amplicon-based sequencing framework for accurately measuring intrahost virus diversity using PrimalSeq and iVar. Genome Biol. 2019, 20, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broad Institute. Picard Tools. Available online: https://broadinstitute.github.io/picard/ (accessed on 8 August 2021).
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, Á.; Scher, E.; Underwood, A.; Jackson, B.; Hill, V.; McCrone, J.T.; Colquhoun, R.; Ruis, C.; Abu-Dahab, K.; Taylor, B.; et al. Assignment of epidemiological lineages in an emerging pandemic using the pangolin tool. Virus Evol. 2021, 7, veab064. [Google Scholar] [CrossRef]
- Ferrando-Martínez, S.; Casazza, J.P.; Leal, M.; Machmach, K.; Muñoz-Fernández, M.A.; Viciana, P.; Koup, R.A.; Ruiz-Mateos, E. Differential Gag-specific polyfunctional T cell maturation patterns in HIV-1 elite controllers. J. Virol. 2012, 86, 3667–3674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fromentin, R.; Bakeman, W.; Lawani, M.B.; Khoury, G.; Hartogensis, W.; DaFonseca, S.; Killian, M.; Epling, L.; Hoh, R.; Sinclair, E.; et al. CD4+ T Cells Expressing PD-1, TIGIT and LAG-3 Contribute to HIV Persistence during ART. PLoS Pathog. 2016, 12, e1005761. [Google Scholar] [CrossRef]
- Pardons, M.; Baxter, A.E.; Massanella, M.; Pagliuzza, A.; Fromentin, R.; Dufour, C.; Leyre, L.; Routy, J.-P.; Kaufmann, D.E.; Chomont, N. Single-cell characterization and quantification of translation-competent viral reservoirs in treated and untreated HIV infection. PLoS Pathog. 2019, 15, e1007619. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Molina, B.; Ferrando-Martinez, S.; Tarancon-Diez, L.; Hernandez-Quero, J.; Genebat, M.; Vidal, F.; Muñoz-Fernández, M.A.; Leal, M.; Koup, R.; Ruiz-Mateos, E. Immune Correlates of Natural HIV Elite Control and Simultaneous HCV Clearance-Supercontrollers. Front Immunol. 2018, 9, 2897. [Google Scholar] [CrossRef] [Green Version]
- Aydillo, T.; Gonzalez-Reiche, A.S.; Aslam, S.; van de Guchte, A.; Khan, Z.; Obla, A.; Dutta, J.; van Bakel, H.; Aberg, J.; García-Sastre, A. Shedding of viable SARS-CoV-2 after Immunosuppressive Therapy for Cancer. N. Engl. J. Med. 2020, 383, 2586–2588. [Google Scholar] [CrossRef]
- Baang, J.H.; Smith, C.; Mirabelli, C.; Valesano, A.L.; Manthei, D.M.; Bachman, M.A.; Wobus, C.E.; Adams, M.; Washer, L.; Martin, E.T.; et al. Prolonged Severe Acute Respiratory Syndrome Coronavirus 2 Replication in an Immunocompromised Patient. J. Infect. Dis. 2021, 223, 23–27. [Google Scholar] [CrossRef]
- Avanzato, V.A.; Matson, M.J.; Seifert, S.N.; Pryce, R.; Williamson, B.N.; Anzick, S.L.; Barbian, K.; Judson, S.D.; Fischer, E.R.; Martens, C.; et al. Case Study: Prolonged Infectious SARS-CoV-2 Shedding from an Asymptomatic Immunocompromised Individual with Cancer. Cell 2020, 183, 1901–1912.e9. [Google Scholar] [CrossRef]
- Sepulcri, C.; Dentone, C.; Mikulska, M.; Bruzzone, B.; Lai, A.; Fenoglio, D.; Bozzano, F.; Bergna, A.; Parodi, A.; Altosole, T.; et al. The longest persistence of viable SARS-CoV-2 with recurrence of viremia and relapsing symptomatic COVID-19 in an immunocompromised patient: A case study. Open Forum Infect. Dis. 2021, ofab217. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.C.; Crain, C.R.; Quiu, X.; Hanage, W.; Li, J.Z. SARS-CoV-2 Sequence Characteristics of COVID-19 Persistence and Reinfection. Clin. Infect. Dis. 2021, ciab380. [Google Scholar] [CrossRef]
- Hensley, M.K.; Bain, W.G.; Jacobs, J.; Nambulli, S.; Parikh, U.; Cillo, A.; Staines, B.; Heaps, A.; Sobolewski, M.D.; Rennick, L.J.; et al. Intractable COVID-19 and Prolonged SARS-CoV-2 Replication in a CAR-T-cell Therapy Recipient: A Case Study. Clin. Infect. Dis. 2021, 73, e815–e821. [Google Scholar] [CrossRef]
- Truong, T.T.; Ryutov, A.; Pandey, U.; Yee, R.; Goldberg, L.; Bhojwani, D.; Aguayo-Hirald, P.; Pinsky, B.A.; Pekosz, A.; Shen, L.; et al. Increased viral variants in children and young adults with impaired humoral immunity and persistent SARS-CoV-2 infection: A consecutive case series. Multicent. Study EBioMed. 2021, 67, 103355. [Google Scholar] [CrossRef]
- Kemp, S.A.; Collier, D.A.; Datir, R.P.; Ferreira, I.A.T.M.; Gayed, S.; Jahun, A.; Hosmillo, M.; Rees-Spear, C.; MIcochova, P.; Lumb, I.U.; et al. SARS-CoV-2 evolution during treatment of chronic infection. Nature 2021, 592, 277–282. [Google Scholar] [CrossRef]
- Khatamzas, E.; Rehn, A.; Muenchhoff, M.; Hellmuth, J.; Gaitzsch, E.; Weiglein, T.; Georgi, E.; Scherer, C.; Stecher, S.; Weigert, O.; et al. Emergence of multiple SARS-CoV-2 mutations in an immunocompromised host. MedRxiv, 2021; in press. [Google Scholar] [CrossRef]
- Lynch, M.; Macori, G.; Fanning, S.; O’Regan, E.; Hunt, E.; O’Callaghan, D.; McCullagh, B.; Jennings, C.; Fortune, A. Genomic Evolution of SARS-CoV-2 Virus in Immunocompromised Patient, Ireland. Emerg. Infect. Dis. 2021, 27, 2499–2501. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Choudhary, M.C.; Regan, J.; Sparks, J.A.; Padera, R.F.; Qiu, X.; Solomon, I.H.; Kuo, H.-H.; Boucau, J.; Bowman, K.; et al. Persistence and evolution of SARS-CoV-2 in an immunocompromised host. N. Engl. J. Med. 2020, 383, 2291–2293. [Google Scholar] [CrossRef]
- Clark, S.A.; Clark, L.E.; Pan, J.; Coscia, A.; McKay, L.G.A.; Shankar, S.; Johnson, R.I.; Brusic, V.; Choudhary, M.C.; Regan, J.; et al. SARS-CoV-2 evolution in an immunocompromised host reveals shared neutralization escape mechanisms. Cell 2021, 184, 2605–2617.e18. [Google Scholar] [CrossRef] [PubMed]
- Tarhini, H.; Recoing, A.; Bridier-Nahmias, A.; Rahi, M.; Lambert, C.; Martres, P.; Lucet, J.-C.; Rioux, C.; Bouzid, D.; Lebourgeois, S.; et al. Long-term severe acute respiratory syndrome coronavirus 2 (SAR-CoV-2) infectiousness among three immunocompromised patients: From prolonged viral shedding to SARS-CoV-2 superinfection. J. Infect. Dis. 2021, 223, 1522–1527. [Google Scholar] [CrossRef]
- Hoffman, S.A.; Costales, C.; Sahoo, M.K.; Palanisamy, S.; Yamamoto, F.; Huang, C.; Verghese, M.; Solis, D.A.; Sibai, M.; Subramanian, A.; et al. SARS-CoV-2 Neutralization Resistance Mutations in Patient with HIV/AIDS, California, USA. Emerg. Infect. Dis. 2021, 27, 2720–2723. [Google Scholar] [CrossRef] [PubMed]
- Corey, L.; Beyrer, C.; Cohen, M.S.; Michael, N.L.; Bedford, T.; Rolland, M. SARS-CoV-2 variants in patients with immunosuppression. N. Engl. J. Med. 2021, 385, 562–566. [Google Scholar] [CrossRef]
- Martinot, M.; Jary, A.; Fafi-Kremer, S.; Leducq, V.; Delagreverie, H.; Garnier, M.; Pacanowski, J.; Mékinian, A.; Pirenne, F.; Tiberghien, P.; et al. Emerging RNA-dependent RNA polymerase mutation in a Remdesivir-treated B-cell immunodeficient patient with protracted coronavirus disease 2019. Clin. Infect. Dis. 2021, 73, e1762–e1765. [Google Scholar] [CrossRef] [PubMed]
- Ambrosioni, J.; Blanco, J.L.; Reyes-Urueña, J.M.; Davies, M.-A.; Sued, O.; Marcos, M.A.; Martínez, E.; Bertagnolio, S.; Alcamí, J.; Miró, J.M. Overview of SARS-CoV-2 infection in adults living with HIV. Lancet HIV 2021, 8, e294–e305. [Google Scholar] [CrossRef]
- Dandachi, D.; Geiger, G.; Montgomery, M.W.; Karmen-Tuohy, S.; Golzy, M.; Antar, A.A.R.; Llibre, J.M.; Camazine, M.; Díaz-De Santiago, A.; Carlucci, P.M.; et al. Characteristics, Comorbidities, and Outcomes in a Multicenter Registry of Patients with HIV and Coronavirus Disease-19. Clin. Infect. Dis. 2021, 73, e1964–e1972. [Google Scholar] [CrossRef] [PubMed]
- Bouaré, F.; Laghmari, M.; Etouche, F.N.; Arjdal, B.; Saidi, I.; Hajhouji, F.; Ghannane, H.; Amro, L.; Tassi, N.; Benali, S.A. Unusual association of COVID-19, pulmonary tuberculosis and human immunodeficiency virus, having progressed favorably under treatment with chloroquine and rifampin. Pan Afr. Med. J. 2020, 35 (Suppl. 2), 110. [Google Scholar] [CrossRef] [PubMed]
- Mang, S.; Kaddu-Mulindwa, D.; Metz, C.; Becker, A.; Seiler, F.; Smola, S.; Maβmann, A.; Becker, S.L.; Papan, C.; Bals, R.; et al. Pneumocystis jirovecii Pneumonia and Severe Acute Respiratory Syndrome Coronavirus 2 Coinfection in a Patient With Newly Diagnosed HIV-1 Infection. Clin. Infect. Dis. 2021, 72, 1487–1489. [Google Scholar] [CrossRef]
- Merchant, E.A.; Flint, K.; Barouch, D.H.; Blair, B.M. Co-infection with coronavirus disease 2019, previously undiagnosed human immunodeficiency virus, Pneumocystis jirovecii pneumonia and cytomegalovirus pneumonitis, with possible immune reconstitution inflammatory syndrome. IDCases 2021, 24, e01153. [Google Scholar] [CrossRef]
- Wang, M.; Luo, L.; Bu, H.; Xia, H. One case of coronavirus disease 2019 (COVID-19) in a patient co-infected by HIV with a low CD4+ T-cell count. Int. J. Infect. Dis. 2020, 96, 148–150. [Google Scholar] [CrossRef]
- Yousaf, M.; Hameed, M.; Alsoub, H.; Khatib, M.; Jamal, W.; Ahmad, M. COVID-19: Prolonged viral shedding in an HIV patient with literature review of risk factors for prolonged viral shedding and its implications for isolation strategies. Clin. Case Rep. 2021, 9, 1397–1401. [Google Scholar] [CrossRef]
- Maan, I. Prolonged SARS-CoV-2 shedding in a person living with advanced HIV and diffuse large B-cell lymphoma. In Proceedings of the 18th European AIDS Conference, London, UK, 27–30 October 2021. Clinical Case Session. [Google Scholar]
- Vizcarra, P.; Pérez-Elías, M.J.; Quereda, C.; Moreno, A.; Vivancos, M.J.; Dronda, F.; Casado, J.L. Description of COVID-19 in HIV-infected individuals: A single-centre, prospective cohort. Lancet HIV 2020, 7, e554–e564. [Google Scholar] [CrossRef]
- Huang, J.; Xie, N.; Hu, X.; Yan, H.; Ding, J.; Liu, P.; Ma, H.; Ruan, L.; Li., G.; He, N.; et al. Epidemiological, virological and serological features of COVID-19 cases in people living with HIV in Wuhan City: A population-based cohort study. Clin. Infect. Dis. 2021, 73, e2086–e2094. [Google Scholar]
- Gutmann, C.; Takov, K.; Burnap, S.A.; Singh, B.; Ali, H.; Theofilatos, K.; Reed, E.; Hasman, M.; Nabeebaccus, A.; Fish, M.; et al. SARS-CoV-2 RNAemia and proteomic trajectories inform prognostication in COVID-19 patients admitted to intensive care. Nat. Commun. 2021, 12, 3406. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.L.; Staines, B.; Bain, W.; Da Silva Castanha, P.; Naqvi, A.; Boltz, V.F.; Marques, E.T.; Denny, T.; Woods, C.; Morris, A.; et al. COVID-19 outcome: Insights from quantification of viremia and neutralizing antibody. In Proceedings of the 28th Virtual Conference on Retroviruses and Opportunistic Infections (CROI), Boston, MA, USA, 6–10 March 2021. Abstract No. 116. [Google Scholar]
- Sharov, K.S. HIV/SARS-CoV-2 co-infection: T cell profile, cytokine dynamics and role of exhausted lymphocytes. Int. J. Infect. Dis. 2021, 102, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Gómez, A.; Vitalle, J.; Gasca-Capote, M.C.; Gutiérrez-Valencia, A.; Trujillo-Rodríguez, M.; Serna-Gallego, A.; Muñoz-Muela, E.; Jiménez-Leon, M.R.; Benhnia, M.R.I.; Rivas-Jeremias, I.; et al. Dendritic cell deficiencies persist seven months after SARS-CoV-2 Infection. Cell Mol. Immunol. 2021, 18, 2128–2139. [Google Scholar] [CrossRef]
- Kim, M.H.; Salloum, S.; Wang, J.Y.; Wong, L.P.; Regan, J.; Lefteri, K.; Manickas-Hill, Z.; Gao, C.; Li, J.Z.; Sadreyev, R.I.; et al. Type I, II, and III Interferon Signatures Correspond to Coronavirus Disease 2019 Severity. J. Infect. Dis. 2021, 224, 777–782. [Google Scholar] [CrossRef]
- Pernas, B.; Grandal, M.; Mena, A.; Castro-Iglesias, A.; Cañizares, A.; Wyles, D.L.; López-Calvo, S.; Pértega, S.; Rodríguez-Osorio, I.; Pedreira, J.D.; et al. High prevalence of subtype F in newly diagnosed HIV-1 persons in northwest Spain and evidence for impaired treatment response. AIDS 2014, 28, 1837–1840. [Google Scholar] [CrossRef] [Green Version]
- Cid-Silva, P.; Margusino-Framiñán, L.; Balboa-Barreiro, V.; Martín-Herranz, I.; Castro-Iglesias, Á.; Pernas-Souto, B.; Llibre, J.M.; Poveda, E. Initial treatment response among HIV subtype F infected participants who started antiretroviral therapy based on integrase inhibitors. AIDS 2018, 32, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Lepik, K.J.; Harrigan, P.R.; Yip, B.; Wang, L.; Robbins, M.A.; Zhang, W.W.; Toy, J.; Akagi, L.; Lima, V.D.; Guillemi, S.; et al. Emergent drug resistance with integrase strand transfer inhibitor-based regimens. AIDS 2017, 31, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Fulcher, J.A.; Du, Y.; Zhang, T.-H.; Sun, R.; Landovitz, R.J. Emergence of integrase resistance mutations during initial therapy containing dolutegravir. Clin. Infect. Dis. 2018, 67, 791–794. [Google Scholar] [CrossRef]
- Pena, M.J.; Chueca, N.; D’Avolio, A.; Zarzalejos, J.M.; Garcia, F. Virological failure in HIV to triple therapy with dolutegravir-based firstline treatment: Rare but possible. Open Forum Infect. Dis. 2019, 6, ofy332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lübke, N.; Jensen, B.; Hüttig, F.; Feldt, T.; Walker, A.; Thielen, A.; Däumer, M.; Obermeier, M.; Kaiser, R.; Knops, E.; et al. Failure of dolutegravir first-line ART with selection of virus carrying R263K and G118R. N. Engl. J. Med. 2019, 381, 887–889. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, S.; Daniel, J.; Forsyth, S.; Smit, E. First reported case of integrase (R263K, G163R) and reverse transcriptase (M184V)-transmitted drug resistance from a drug-naive patient failing Triumeq. AIDS 2018, 32, 1905–1907. [Google Scholar] [CrossRef]
- Chamberlain, N.; Mena, L.; Brock, J.B. Case report: Emergent Resistance in a treatment-naïve person with human immunodeficiency virus under Bictegravir-based therapy. Open Forum Infect. Dis. 2021, 8, ofab297. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvarez, H.; Ruiz-Mateos, E.; Juiz-González, P.M.; Vitallé, J.; Viéitez, I.; Vázquez-Friol, M.d.C.; Torres-Beceiro, I.; Pérez-Gómez, A.; Gallego-García, P.; Estévez-Gómez, N.; et al. SARS-CoV-2 Evolution and Spike-Specific CD4+ T-Cell Response in Persistent COVID-19 with Severe HIV Immune Suppression. Microorganisms 2022, 10, 143. https://doi.org/10.3390/microorganisms10010143
Álvarez H, Ruiz-Mateos E, Juiz-González PM, Vitallé J, Viéitez I, Vázquez-Friol MdC, Torres-Beceiro I, Pérez-Gómez A, Gallego-García P, Estévez-Gómez N, et al. SARS-CoV-2 Evolution and Spike-Specific CD4+ T-Cell Response in Persistent COVID-19 with Severe HIV Immune Suppression. Microorganisms. 2022; 10(1):143. https://doi.org/10.3390/microorganisms10010143
Chicago/Turabian StyleÁlvarez, Hortensia, Ezequiel Ruiz-Mateos, Pedro Miguel Juiz-González, Joana Vitallé, Irene Viéitez, María del Carmen Vázquez-Friol, Isabel Torres-Beceiro, Alberto Pérez-Gómez, Pilar Gallego-García, Nuria Estévez-Gómez, and et al. 2022. "SARS-CoV-2 Evolution and Spike-Specific CD4+ T-Cell Response in Persistent COVID-19 with Severe HIV Immune Suppression" Microorganisms 10, no. 1: 143. https://doi.org/10.3390/microorganisms10010143
APA StyleÁlvarez, H., Ruiz-Mateos, E., Juiz-González, P. M., Vitallé, J., Viéitez, I., Vázquez-Friol, M. d. C., Torres-Beceiro, I., Pérez-Gómez, A., Gallego-García, P., Estévez-Gómez, N., De Chiara, L., Poveda, E., Posada, D., & Llibre, J. M. (2022). SARS-CoV-2 Evolution and Spike-Specific CD4+ T-Cell Response in Persistent COVID-19 with Severe HIV Immune Suppression. Microorganisms, 10(1), 143. https://doi.org/10.3390/microorganisms10010143