
Next-Generation Sequencing-Based Study of Helicobacter pylori Isolates from Myanmar and Their Susceptibility to Antibiotics

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. H. pylori Infection Status and Gastritis Histology Score
2.3. Antibiotic Susceptibility Test
2.4. Molecular Analysis of H. pylori Resistant Strains
2.5. Data Analysis
3. Results
3.1. H. pylori Infection Status
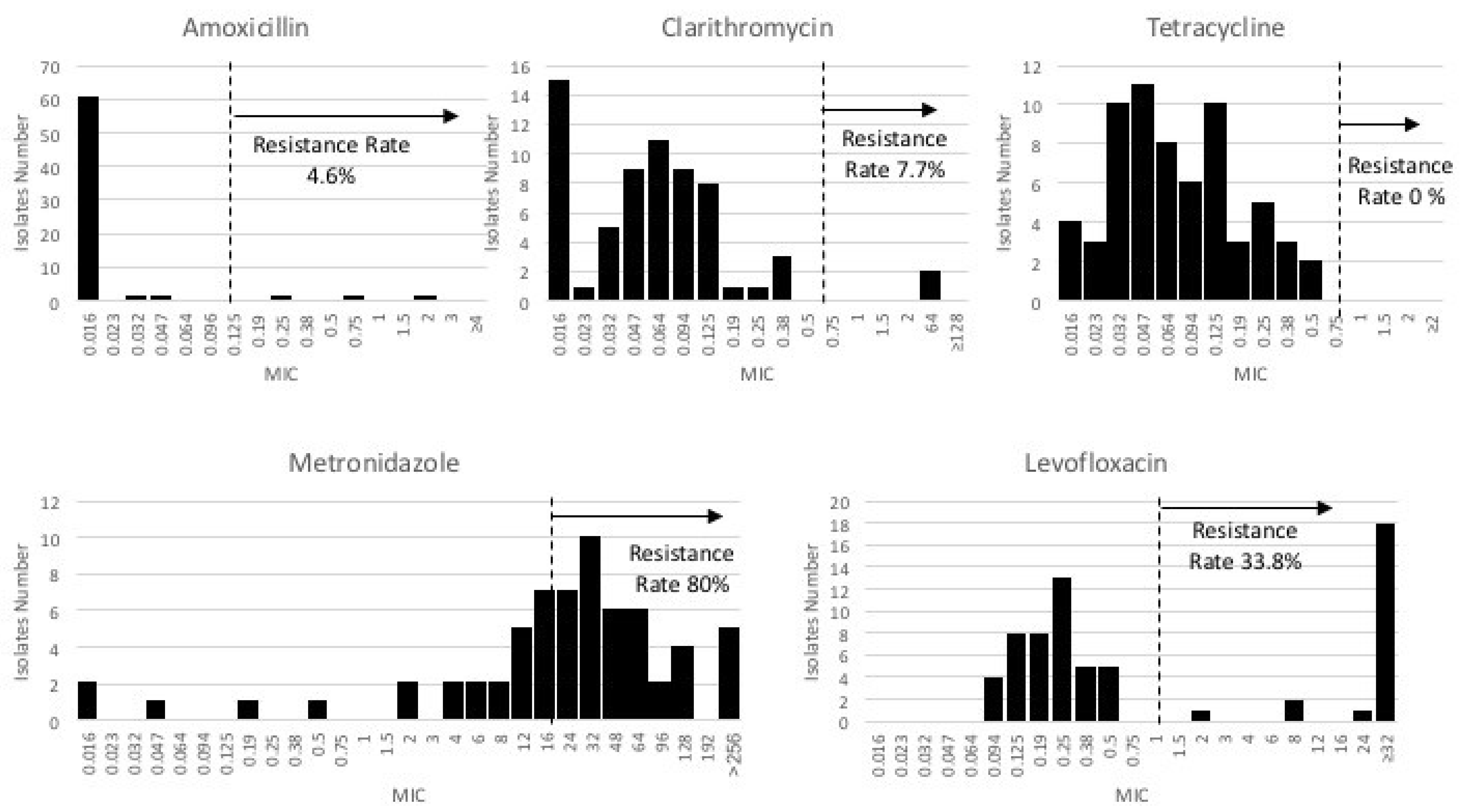
3.2. Antibiotic Susceptibility Test
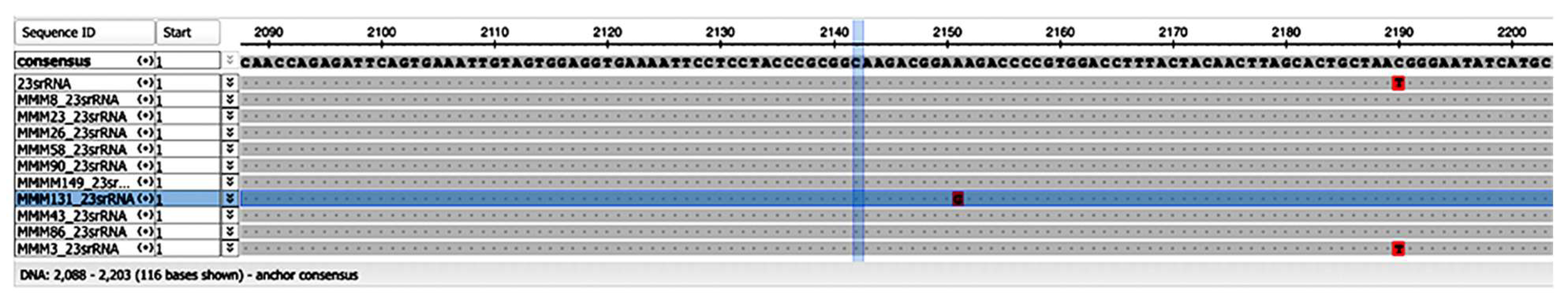
3.3. Genetic Mutation Study
3.3.1. Amoxicillin Resistance
3.3.2. Clarithromycin Resistance
3.3.3. Levofloxacin Resistance
3.3.4. Metronidazole Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marshall, B.J.; Warren, J.R. Unidentified curved bacilli in the stomach of patients with gastritis and peptic ulceration. Lancet 1984, 1, 1311–1315. [Google Scholar] [CrossRef]
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global prevalence of Helicobacter pylori infection: Systematic review and meta-analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [Green Version]
- Nagy, P.; Johansson, S.; Molloy-Bland, M. Systematic review of time trends in the prevalence of Helicobacter pylori infection in China and the USA. Gut Pathog. 2016, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Malaty, H.M.; Graham, D.Y. Importance of childhood socioeconomic status on the current prevalence of Helicobacter pylori infection. Gut 1994, 35, 742–745. [Google Scholar] [CrossRef] [Green Version]
- Moayyedi, P.; Axon, A.T.R.; Feltbower, R.; Duffett, S.; Crocombe, W.; Braunholtz, D.; Richards, I.D.G.; Dowell, A.C.; Forman, D. Relation of adult lifestyle and socioeconomic factors to the prevalence of Helicobacter pylori infection. Int. J. Epidemiol. 2002, 31, 624–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, D.Y.; Dore, M.P.; Lu, H. Understanding treatment guidelines with bismuth and non-bismuth quadruple Helicobacter pylori eradication therapies. Expert Rev. Anti-Infective Ther. 2018, 16, 679–687. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, V.; Giorgio, F.; Hassan, C.; Manes, G.; Vannella, L.; Panella, C.; Ierardi, E.; Zullo, A. Worldwide H. pylori antibiotic resistance: A systematic review. J. Gastrointest. Liver Dis. 2010, 19, 409–414. [Google Scholar]
- Miftahussurur, M.; Cruz, M.; Subsomwong, P.; Abreu, J.A.J.; Hosking, C.; Nagashima, H.; Akada, J.; Yamaoka, Y. Clarithromycin-based triple therapy is still useful as an initial treatment for Helicobacter pylori infection in the Dominican Republic. Am. J. Trop. Med. Hyg. 2017, 96, 1050–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myint, T.; Shiota, S.; Vilaichone, R.K.; Aye, T.T.; Matsuda, M.; Tran, T.T.H.; Uchida, T.; Mahachai, V.; Yamaoka, Y. Prevalence of Helicobacter pylori infection and atrophic gastritis in patients with dyspeptic symptoms in Myanmar. World J. Gastroenterol. WJG 2015, 21, 629. [Google Scholar] [CrossRef]
- Vilaichone, R.-K.; Quach, D.T.; Yamaoka, Y.; Sugano, K.; Mahachai, V. Prevalence and pattern of antibiotic resistant strains of Helicobacter pylori infection in ASEAN. Asian Pac. J. Cancer Prev. 2018, 19, 1411–1413. [Google Scholar] [CrossRef] [PubMed]
- Syam, A.F.; Miftahussurur, M.; Makmun, D.; Nusi, I.A.; Zain, L.H.; Akil, F.; Uswan, W.B.; Simanjuntak, D.; Uchida, T.; Adi, P.; et al. Risk factors and prevalence of Helicobacter pylori in five largest islands of Indonesia: A preliminary study. PLoS ONE 2015, 10, e0140186. [Google Scholar] [CrossRef]
- Miftahussurur, M.; Syam, A.F.; Nusi, I.A.; Makmun, D.; Waskito, L.A.; Zein, L.H.; Akil, F.; Uwan, W.B.; Simanjuntak, D.; Wibawa, I.D.N.; et al. Surveillance of Helicobacter pylori antibiotic susceptibility in Indonesia: Different resistance types among regions and with novel genetic mutations. PLoS ONE 2016, 11, e0166199. [Google Scholar] [CrossRef] [PubMed]
- Eucast, T. European Committee on Antimicrobial Susceptibility Testing, Breakpoint Tables for Interpretation of MICs and Zone Diameters; European Society of Clinical Microbiology and Infectious Diseases: Basel, Switzerland, 2015. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Altschup, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kazutaka, K.; Misakwa, K.; Kei-ichi, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Page, A.J.; Taylor, B.; Delaney, A.J.; Soares, J.; Seemann, T.; Keane, J.A.; Harris, S.R. SNP-sites: Rapid efficient extraction of SNPs from multi-FASTA alignments. Microb. Genom. 2016, 2, e000056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerrits, M.M.; Schuijffel, D.; van Zwet, A.A.; Kuipers, E.J.; Vandenbroucke-Grauls, C.M.J.E.; Kusters, J.G. Alterations in Penicillin-Binding Protein 1A Confer Resistance to β-Lactam Antibiotics in Helicobacter pylori. Antimicrob. Agents Chemother. 2002, 46, 2229–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimbara, E.; Noguchi, N.; Kawai, T.; Sasatsu, M. Mutations in penicillin-binding proteins 1, 2 and 3 are responsible for amoxicillin resistance in Helicobacter pylori. J. Antimicrob. Chemother. 2008, 61, 995–998. [Google Scholar] [CrossRef] [Green Version]
- Stone, G.G.; Shortridge, D.; Flamm, R.K.; Versalovic, J.; Beyer, J.; Idler, K.; Zulawinski, L.; Tanaka, S.K. Identification of a 23S rRNA gene mutation in clarithromycin-resistant Helicobacter pylori. Helicobacter 1996, 1, 227–228. [Google Scholar] [CrossRef] [PubMed]
- Cabral JH, M.; Jackson, A.P.; Smith, C.V.; Shikotra, N.; Maxwell, A.; Liddington, R.C. Crystal structure of the breakage–reunion domain of DNA gyrase. Nature 1997, 388, 903–906. [Google Scholar] [CrossRef] [PubMed]
- Wohlkonig, A.; Chan, P.F.; Fosberry, A.P.; Homes, P.; Huang, J.; Kranz, M.; Leydon, V.R.; Miles, T.J.; Pearson, N.D.; Perera, R.; et al. Structural basis of quinolone inhibition of type IIA topoisomerases and target-mediated resistance. Nat. Struct. Mol. Biol. 2010, 17, 1152–1153. [Google Scholar] [CrossRef]
- Camorlinga-Ponce, M.; Gómez-Delgado, A.; Aguilar-Zamora, E.; Torres, R.C.; Giono-Cerezo, S.; Escobar-Ogaz, A.; Torres, J. Phenotypic and genotypic antibiotic resistance patterns in Helicobacter pylori strains from ethnically diverse population in México. Front. Cell. Infect. Microbiol. 2021, 10, 539115. [Google Scholar] [CrossRef] [PubMed]
- Dattoli, V.C.C.; Veiga, R.V.; Da Cunha, S.S.; Pontes-De-Carvalho, L.C.; Barreto, M.L.; Alcântara-Neves, N.M. Seroprevalence and potential risk factors for Helicobacter pylori infection in Brazilian children. Helicobacter 2010, 15, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Savoldi, A.; Carrara, E.; Graham, D.Y.; Conti, M.; Tacconelli, E. Prevalence of antibiotic resistance in Helicobacter pylori: A systematic review and meta-analysis in World Health Organization regions. Gastroenterology 2018, 155, 1372–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, Y.-T.; Liou, J.-M.; El-Omar, E.; Wu, J.-Y.; Leow, A.H.R.; Goh, K.L.; Das, R.; Lu, H.; Lin, J.-T.; Tu, Y.-K.; et al. Primary antibiotic resistance in Helicobacter pylori in the Asia-Pacific region: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2017, 2, 707–715. [Google Scholar] [CrossRef]
- Čižman, M. The use and resistance to antibiotics in the community. Int. J. Antimicrob. Agents 2003, 21, 297–307. [Google Scholar] [CrossRef]
- Organization world Health. UN-Water Global Analysis and Assessment of Sanitation and Drinking-Water (GLAAS) 2017 Report: Financing Universal Water, Sanitation and Hygiene Under the Sustainable Development Goals; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Sukri, A.; Lopes, B.S.; Hanafiah, A. The Emergence of multidrug-resistant Helicobacter pylori in Southeast Asia: A systematic review on the trends and intervention strategies using antimicrobial peptides. Antibiotics 2021, 10, 1061. [Google Scholar] [CrossRef] [PubMed]
- Malfertheiner, P.; Megraud, F.; O’morain, C.A.; Gisbert, J.P.; Kuipers, E.J.; Axon, A.T.; Bazzoli, F.; Gasbarrini, A.; Atherton, J.; Graham, D.Y.; et al. Management of Helicobacter pylori infection—the Maastricht V/Florence consensus report. Gut 2017, 66, 6–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gisbert, J.P.; Morena, F. Systematic review and meta-analysis: Levofloxacin-based rescue regimens after Helicobacter pylori treatment failure. Aliment Pharmacol. Ther. 2006, 23, 35–44. [Google Scholar] [CrossRef]
- Fock, K.M.; Katelaris, P.; Sugano, K.; Ang, T.L.; Hunt, R.; Talley, N.J.; Lam, S.K.; Xiao, S.D.; Tan, H.J.; Wu, C.Y.; et al. Second Asia–Pacific consensus guidelines for Helicobacter pylori infection. J. Gastroenterol. Hepatol. 2009, 24, 1587–1600. [Google Scholar] [CrossRef]
- Tshibangu-Kabamba, E.; Yamaoka, Y. Helicobacter pylori infection and antibiotic resistance—from biology to clinical implications. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Tshibangu-Kabamba, E.; de Jesus Ngoma-Kisoko, P.; Tuan, V.P.; Matsumoto, T.; Akada, J.; Kido, Y.; Tshimpi-Wola, A.; Tshiamala-Kashala, P.; Ahuka-Mundeke, S.; Ngoyi, D.M.; et al. Next-generation sequencing of the whole bacterial genome for tracking molecular insight into the broad-spectrum antimicrobial resistance of Helicobacter pylori clinical isolates from the Democratic Republic of Congo. Microorganisms 2020, 8, 887. [Google Scholar] [CrossRef]
- Tuan, V.P.; Narith, D.; Tshibangu-Kabamba, E.; Dung, H.D.Q.; Viet, P.T.; Sokomoth, S.; Binh, T.T.; Sokhem, S.; Tri, T.D.; Ngov, S.; et al. A Next-generation sequencing-based approach to identify genetic determinants of antibiotic resistance in Cambodian Helicobacter pylori clinical isolates. J. Clin. Med. 2019, 8, 858. [Google Scholar] [CrossRef] [Green Version]
- Cambau, E.; Allerheiligen, V.; Coulon, C.; Corbel, C.; Lascols, C.; Deforges, L.; Soussy, C.-J.; Delchier, J.-C.; Megraud, F. Evaluation of a new test, genotype HelicoDR, for molecular detection of antibiotic resistance in Helicobacter pylori. J. Clin. Microbiol. 2009, 47, 3600–3607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishizawa, T.; Suzuki, H. Mechanisms of Helicobacter pylori antibiotic resistance and molecular testing. Front. Mol. Biosci. 2014, 1, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liou, J.-M.; Chang, C.-Y.; Sheng, W.-H.; Wang, Y.-C.; Chen, M.-J.; Lee, Y.-C.; Hung, H.-W.; Chian, H.; Chang, S.-C.; Wu, M.-S.; et al. Genotypic resistance in Helicobacter pylori strains correlates with susceptibility test and treatment outcomes after levofloxacin- and clarithromycin-based therapies. Antimicrob. Agents Chemother. 2011, 55, 1123–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.M.; Jung, H.C.; Song, I.S. Distribution of fluoroquinolone MICs in Helicobacter pylori strains from Korean patients. J. Antimicrob. Chemother. 2005, 56, 965–967. [Google Scholar] [CrossRef] [Green Version]
- Rimbara, E.; Noguchi, N.; Kawai, T.; Sasatsu, M. Fluoroquinolone resistance in Helicobacter pylori: Role of mutations at position 87 and 91 of gyrA on the level of resistance and identification of a resistance conferring mutation in gyrB. Helicobacter 2012, 17, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Hanafiah, A.; Binmaeil, H.; Ali, R.A.R.; Rose, I.M.; Lopes, B.S. Molecular characterization and prevalence of antibiotic resistance in Helicobacter pylori isolates in Kuala Lumpur, Malaysia. Infect. Drug Resist. 2019, 12, 3051–3061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziver-Sarp, T.; Yuksel-Mayda, P.; Saribas, S.; Demiryas, S.; Gareayaghi, N.; Ergin, S.; Tasci, I.; Ozbey, D.; Bal, K.; Erzin, Y.; et al. Point mutations at gyrA and gyrB genes of levofloxacin resistant Helicobacter pylori strains and dual resistance with clarithromycin. Clin. Lab. 2021, 67, 10. [Google Scholar] [CrossRef]
- Zhang, Y.; Wen, Y.; Xiao, Q.; Zheng, W.; Long, G.; Chen, B.; Shu, X.; Jiang, M. Mutations in the antibiotic target genes related to clarithromycin, metronidazole and levofloxacin resistance in Helicobacter pylori strains from children in China. Infect. Drug Resist. 2020, 13, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, E.-G.; Debowski, A.W.; Webberley, K.M.; Peters, F.; Lamichhane, B.; Loke, M.-F.; Vadivelu, J.; Tay, C.-Y.; Marshall, B.J.; Wise, M.J. Analysis of core protein clusters identifies candidate variable sites conferring metronidazole resistance in Helicobacter pylori. Gastroenterol. Rep. 2019, 7, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Solcà, N.M.; Bernasconi, M.V.; Piffaretti, J.-C. Mechanism of metronidazole resistance in Helicobacter pylori: Comparison of the rdxA gene sequences in 30 strains. Antimicrob. Agents Chemother. 2000, 44, 2207–2210. [Google Scholar] [CrossRef] [Green Version]
- Mendz, G.L.; Mégraud, F. Is the molecular basis of metronidazole resistance in microaerophilic organisms understood? Trends Microbiol. 2002, 10, 370–375. [Google Scholar] [CrossRef]
- Marais, A.; Bilardi, C.; Cantet, F.; Mendz, G.L.; Mégraud, F. Characterization of the genes rdxA and frxA involved in metronidazole resistance in Helicobacter pylori. Res. Microbiol. 2003, 154, 137–144. [Google Scholar] [CrossRef]
- Aftab, H.; Miftahussurur, M.; Subsomwong, P.; Ahmed, F.; Khan, A.A.; Yamaoka, Y. Helicobacter pylori antibiotic susceptibility patterns in Bangladesh: Emerging levofloxacin resistance. J. Infect. Dev. Ctries. 2016, 10, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Jenks, P.J.; Edwards, D.I. Metronidazole resistance in Helicobacter pylori. Int. J. Antimicrob. Agents 2002, 19, 1–7. [Google Scholar] [CrossRef]
- De Palma, G.Z.; Mendiondo, N.; Wonaga, A.; Viola, L.; Ibarra, D.; Campitelli, E.; Salim, N.; Corti, R.; Goldman, C.; Catalano, M. Occurrence of mutations in the antimicrobial target genes related to levofloxacin, clarithromycin, and amoxicillin resistance in Helicobacter pylori isolates from Buenos Aires city. Microb. Drug Resist. 2017, 23, 351–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.J.; Kim, J.G. Substitutions in penicillin-binding protein 1 in amoxicillin-resistant Helicobacter pylori strains isolated from Korean patients. Gut Liver 2013, 7, 655–660. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, R.B.; Lopes, R.A.B.; Lopes, G.A.D.C.; Ho, T.H.; Sperança, M.A. Low Helicobacter pylori primary resistance to clarithromycin in gastric biopsy specimens from dyspeptic patients of a city in the interior of São Paulo, Brazil. BMC Gastroenterol. 2013, 13, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deyi, V.Y.M.; Burette, A.; Bentatou, Z.; Maaroufi, Y.; Bontems, P.; Lepage, P.; Reynders, M. Practical use of GenoType® HelicoDR, a molecular test for Helicobacter pylori detection and susceptibility testing. Diagn. Microbiol. Infect. Dis. 2011, 70, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Fauzia, K.A.; Miftahussurur, M.; Syam, A.F.; Waskito, L.A.; Doohan, D.; Rezkitha, Y.A.A.; Matsumoto, T.; Tuan, V.P.; Akada, J.; Yonezawa, H.; et al. Biofilm formation and antibiotic resistance phenotype of Helicobacter pylori clinical isolates. Toxins 2020, 12, 473. [Google Scholar] [CrossRef] [PubMed]
- Ierardi, E.; Losurdo, G.; Mileti, A.; Paolillo, R.; Giorgio, F.; Principi, M.; Di Leo, A. The puzzle of coccoid forms of Helicobacter pylori: Beyond basic science. Antibiotics 2020, 9, 293. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristic | n | Single Antibiotic Resistance (%) | ||||
|---|---|---|---|---|---|---|
| AMX | CAM | LVX | MNZ | TCN | ||
| Total | 65 | 3 (4.6) | 5 (7.7) | 22 (33.8) | 52 (80.0) | 0 (0.0) |
| Sex | ||||||
| Male | 45 | 3 (6.7) | 4 (8.9) | 13 (28.9) | 35 (77.8) | 0 (0.0) |
| Female | 20 | 0 (0.0) | 1 (5.0) | 9 (45.0) | 17 (85.0) | 0 (0.0) |
| Location | ||||||
| Mawlamyine | 44 | 3 (6.8) | 3 (6.8) | 12(27.3) | 34 (77.3) | 0 (0.0) |
| Yangon | 21 | 0 (0.0) | 2 (9.5) | 10 (47.6) | 18 (85.7) | 0 (0.0) |
| Ethnicity | ||||||
| Bamer | 53 | 2 () | 2 (3.8) | 17 (32.1) | 43 (81.1) | 0 (0.0) |
| Kayen | 4 | 0 (0.0) | 1 (25.0) | 2 (50.0) | 4 (100) | 0 (0.0) |
| Mon | 5 | 1 (.0) | 2 (40.0) | 3 (60.0) | 3 (60.0) | 0 (0.0) |
| Others | 3 | 0 (0.0) | 0 (0.0) | 0 (0.0) | 2 (66.7) | 0 (0.0) |
| Susceptibility Test Result | n (%) |
|---|---|
| Number of strains | 65 |
| Sensitive | 8 (12.3) |
| Single-drug resistance | 35 (53.8) |
| AMX | 0 (0.0) |
| CAM | 1 (1.5) |
| LVX | 2 (3.1) |
| MNZ | 32 (49.2) |
| TCN | 0 (0.0) |
| Double-drug resistance | 19 (29.2) |
| AMX + CAM | 1 (5.3) |
| AMX + MNZ | 1 (5.3) |
| AMX + LVX | 1 (5.3) |
| MNZ + LVX | 16 (84.2) |
| Triple-drug resistance | 3 (4.6) |
| CAM + MNZ + LVX | 3 (100) |
| Gene | Strain | Sensitive (n = 5) | MMM3 | MMM37 | MMM25 |
|---|---|---|---|---|---|
| pbp1 | MIC (mg/L) | 2 | 0.25 | 0.75 | |
| Position | |||||
| 45 | V/I | V/I | |||
| 414 | S/R | V/R | |||
| 465 | D-Del | D/K | |||
| 471 | V/M | ||||
| 564 | N/Y |
| Gene | Strain | Sensitive (n = 5) | MMM3 | MMM43 | MMM86 | MMM131 | MMM149 |
|---|---|---|---|---|---|---|---|
| MIC (mg/L) | 0.38 | 16 | 8 | 128 | 64 | ||
| 23S rRNA | Position | ||||||
| 64 | G/A | ||||||
| 186 | T/C | ||||||
| 228 | G/A | ||||||
| 242 | T-Del | ||||||
| 243 | T-Del | ||||||
| 247 | A-Ins | ||||||
| 248 | T/C | T/C | T/C | ||||
| 425 | A/G | ||||||
| 513 | C/T | ||||||
| 559 | C/T | ||||||
| 763 | A/G | ||||||
| 822 | G/A | ||||||
| 1286 | A/G | ||||||
| 1312 | C/T | ||||||
| 2219 | C/T | ||||||
| 2937 | G/A | ||||||
| rpl22 | 16 | T/C | |||||
| 20 | G/A | ||||||
| 69 | T/C | T/C | |||||
| 125 | C/T | ||||||
| 135 | G/A | ||||||
| 147 | G/T | ||||||
| 237 | GCG-Ins | C/T | |||||
| 267 | A/G |
| Gene | Strain | MMM33 | MMM37 | MMM43 | MMM44 | MMM47 | MMM54 | MMM62 | MMM131 | MMM135 | MMM145 | MMM149 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC (mg/L) | 32 | >32 | >32 | >32 | >32 | >32 | >32 | >32 | >32 | 2 | >32 | |
| gyrA | Position | |||||||||||
| 3 | D/G | D/G | ||||||||||
| 20 | ||||||||||||
| 91 | D/N | D/N | D/N | D/G | D/Y | D/N | D/N | |||||
| 172 | V/I | |||||||||||
| 208 | G/E | G/R | G/E | |||||||||
| 210 | D/N | |||||||||||
| 230 | K/Q | |||||||||||
| 234 | I/V | |||||||||||
| 246 | V/M | |||||||||||
| 285 | ||||||||||||
| 407 | A/V | A/V | A/V | |||||||||
| 497 | D/E | |||||||||||
| 517 | V/M | |||||||||||
| 524 | A/V | A/V | ||||||||||
| 557 | I/T | |||||||||||
| 612 | R/C | |||||||||||
| 635 | R/K | |||||||||||
| 661 | A/T | |||||||||||
| 668 | T/A | |||||||||||
| 684 | I/M | I/M | I/M | |||||||||
| 688 | T/A | T/A | ||||||||||
| 703 | V/I | |||||||||||
| 709 | S/N | S/N | S/N | |||||||||
| 712 | G/S | G/S | G/S | |||||||||
| 735 | ||||||||||||
| 760 | D/M | |||||||||||
| Truncated | 1-285 | 1-735 | ||||||||||
| gyrB | 64 | E/G | ||||||||||
| 160 | S/N | |||||||||||
| 215 | T/A | |||||||||||
| 230 | A/T | |||||||||||
| 240 | S/A | |||||||||||
| 479 | S/G | |||||||||||
| 573 | N/G | |||||||||||
| 584 | A/V | |||||||||||
| 614 | V/I | V/I | ||||||||||
| 620 | L/S | |||||||||||
| 676 | M/V | |||||||||||
| 679 | N/H | |||||||||||
| Truncated | 1-1476 | 1-1476 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subsomwong, P.; Doohan, D.; Fauzia, K.A.; Akada, J.; Matsumoto, T.; Yee, T.T.; Htet, K.; Waskito, L.A.; Tuan, V.P.; Uchida, T.; et al. Next-Generation Sequencing-Based Study of Helicobacter pylori Isolates from Myanmar and Their Susceptibility to Antibiotics. Microorganisms 2022, 10, 196. https://doi.org/10.3390/microorganisms10010196
Subsomwong P, Doohan D, Fauzia KA, Akada J, Matsumoto T, Yee TT, Htet K, Waskito LA, Tuan VP, Uchida T, et al. Next-Generation Sequencing-Based Study of Helicobacter pylori Isolates from Myanmar and Their Susceptibility to Antibiotics. Microorganisms. 2022; 10(1):196. https://doi.org/10.3390/microorganisms10010196
Chicago/Turabian StyleSubsomwong, Phawinee, Dalla Doohan, Kartika Afrida Fauzia, Junko Akada, Takashi Matsumoto, Than Than Yee, Kyaw Htet, Langgeng Agung Waskito, Vo Phuoc Tuan, Tomohisa Uchida, and et al. 2022. "Next-Generation Sequencing-Based Study of Helicobacter pylori Isolates from Myanmar and Their Susceptibility to Antibiotics" Microorganisms 10, no. 1: 196. https://doi.org/10.3390/microorganisms10010196
APA StyleSubsomwong, P., Doohan, D., Fauzia, K. A., Akada, J., Matsumoto, T., Yee, T. T., Htet, K., Waskito, L. A., Tuan, V. P., Uchida, T., Matsuhisa, T., & Yamaoka, Y. (2022). Next-Generation Sequencing-Based Study of Helicobacter pylori Isolates from Myanmar and Their Susceptibility to Antibiotics. Microorganisms, 10(1), 196. https://doi.org/10.3390/microorganisms10010196