
In Vitro Effects of Lactobacillus plantarum LN66 and Antibiotics Used Alone or in Combination on Helicobacter pylori Mature Biofilm

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Medium and Growth Conditions
2.2. MICs of Antibiotics
2.3. Collection of LN66 CFS
2.4. Characterization of L. plantarum LN66
2.5. MIC of LN66 CFS
2.6. FICs of Antibiotics and CFS
2.7. Construction of Biofilm
2.8. Analysis of CFS Effectiveness after Different Treatments
2.9. Drug Sensitivity before and after Biofilm Formation
2.10. Effect of CFS Combined with Antibiotics on H. pylori Biofilm Viability
2.11. Effect of CFS Combined with Antibiotics on H. pylori Biofilm EPS
2.12. Effect of CFS Combined with Antibiotics on H. pylori Biofilm SEM Observations
2.13. Effect of CFS Combined with Antibiotics on H. pylori Biofilm CLSM Observations
2.14. Effect of CFS Combined with Antibiotics on H. pylori Biofilm Virulence Gene Expression
2.15. Effect of CFS Combined with Antibiotics on H. pylori Biofilm Live Cell Dispersion
2.16. Effect of CFS Combined with Antibiotics on H. pylori Planktonic Cell Viability
2.17. Statistical Analysis
3. Results
3.1. MICs and Characterization of LN66
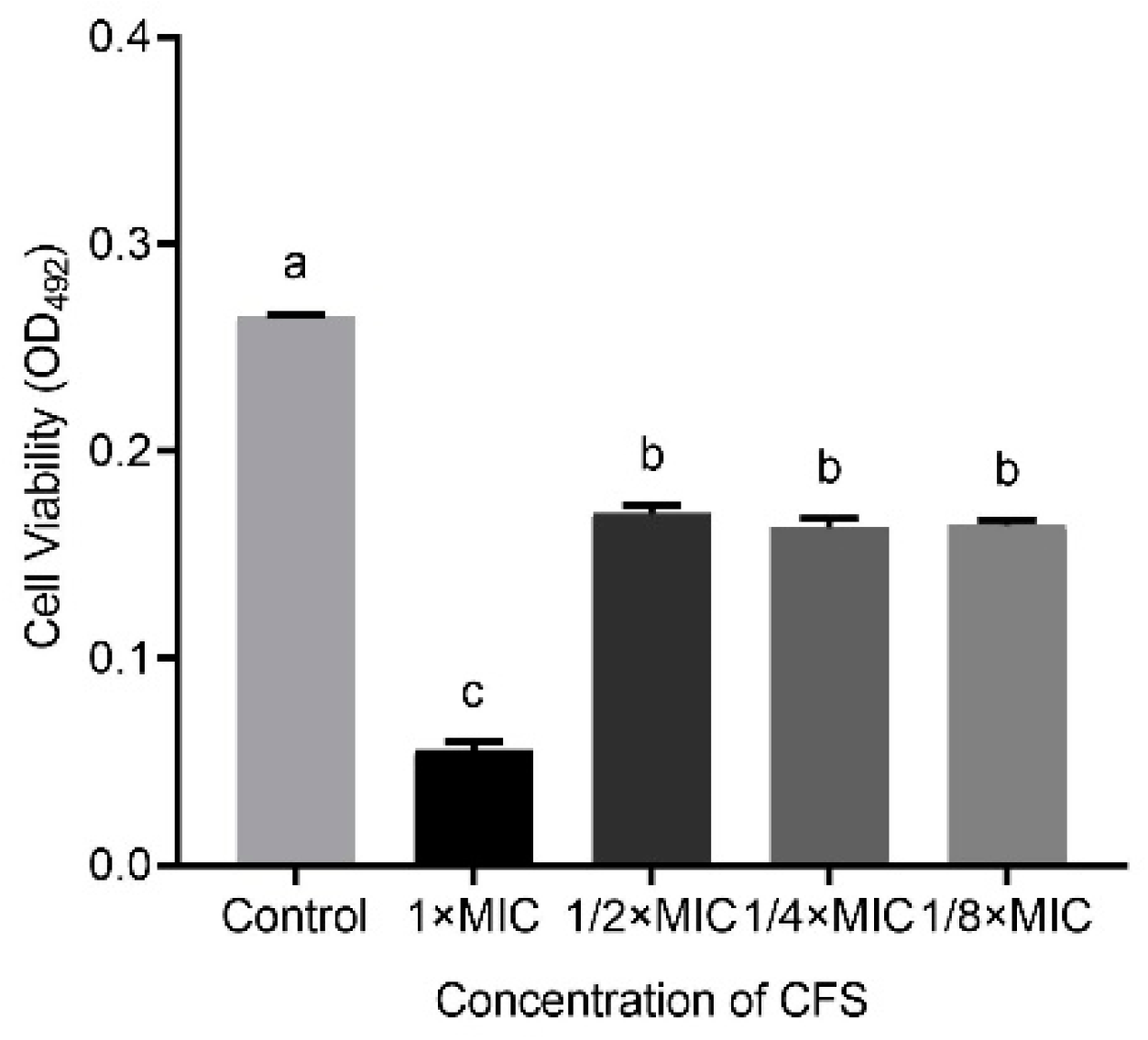
3.2. The Eradication Effect of LN66 CFS on H. pylori Mature Biofilms Is Derived Mainly from Heat-Stable Peptides
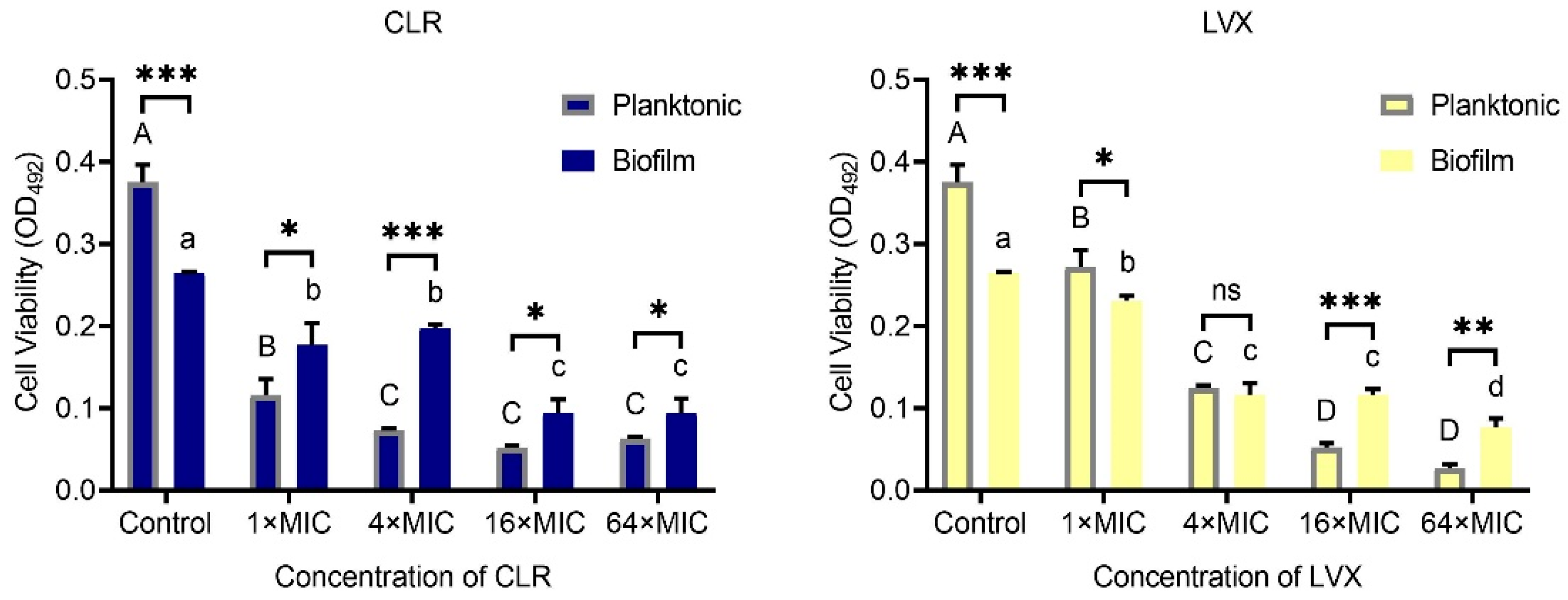
3.3. H. pylori Sensitivity to High Concentrations of CLR and LVX Is Reduced after Mature Biofilm Formation
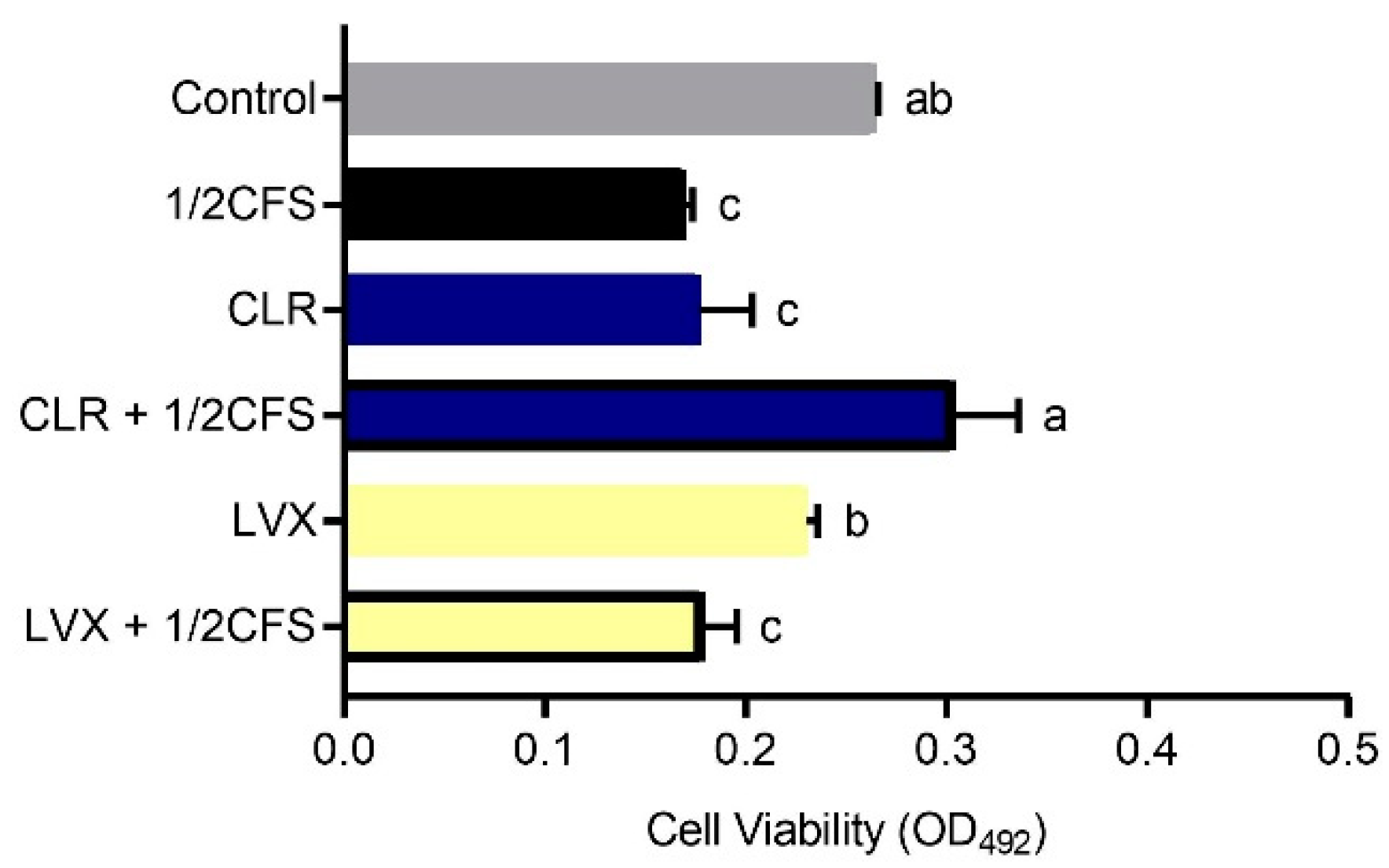
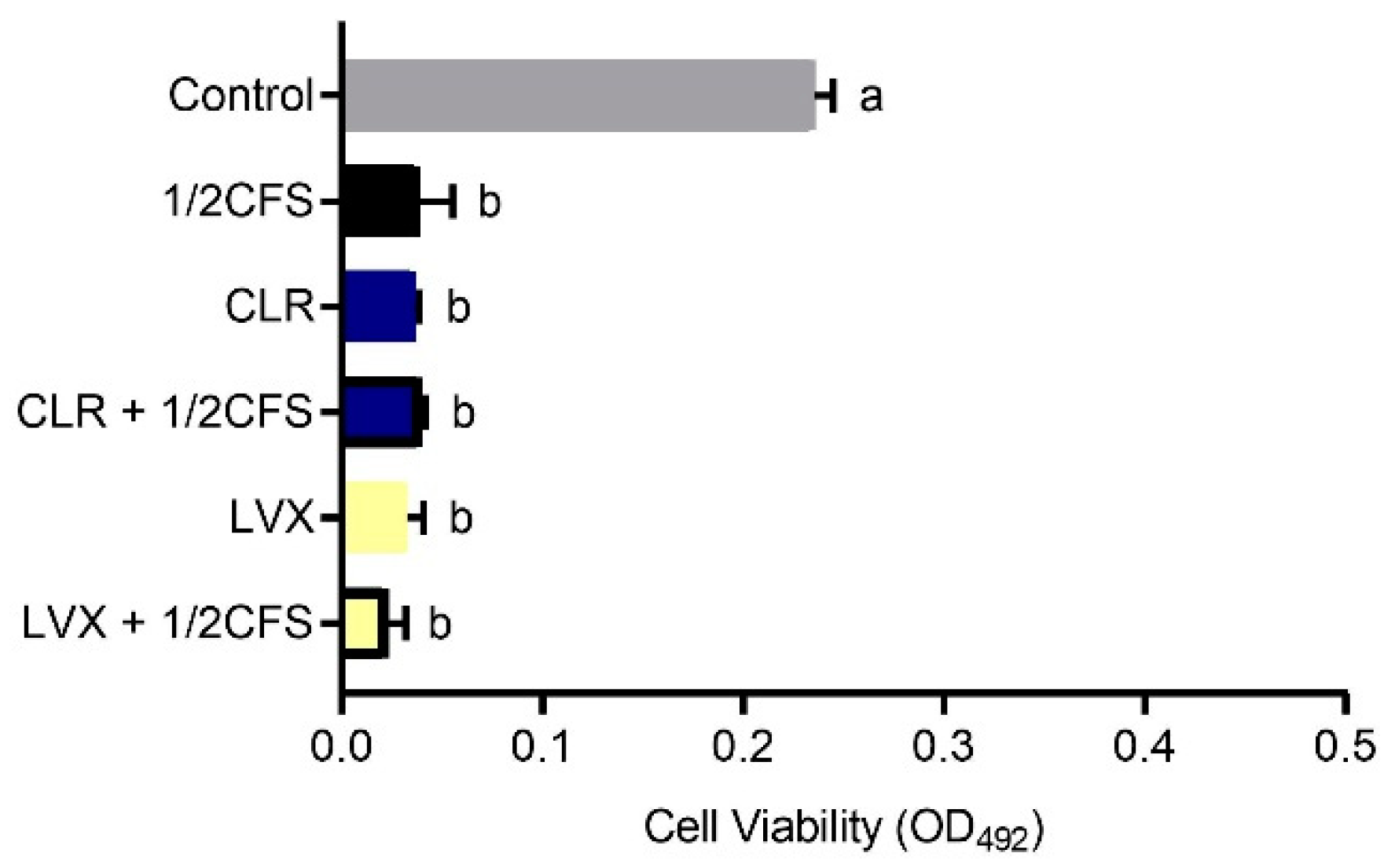
3.4. Combination of CFS and Different Antibiotics Can Influence the Viability of H. pylori Mature Biofilms
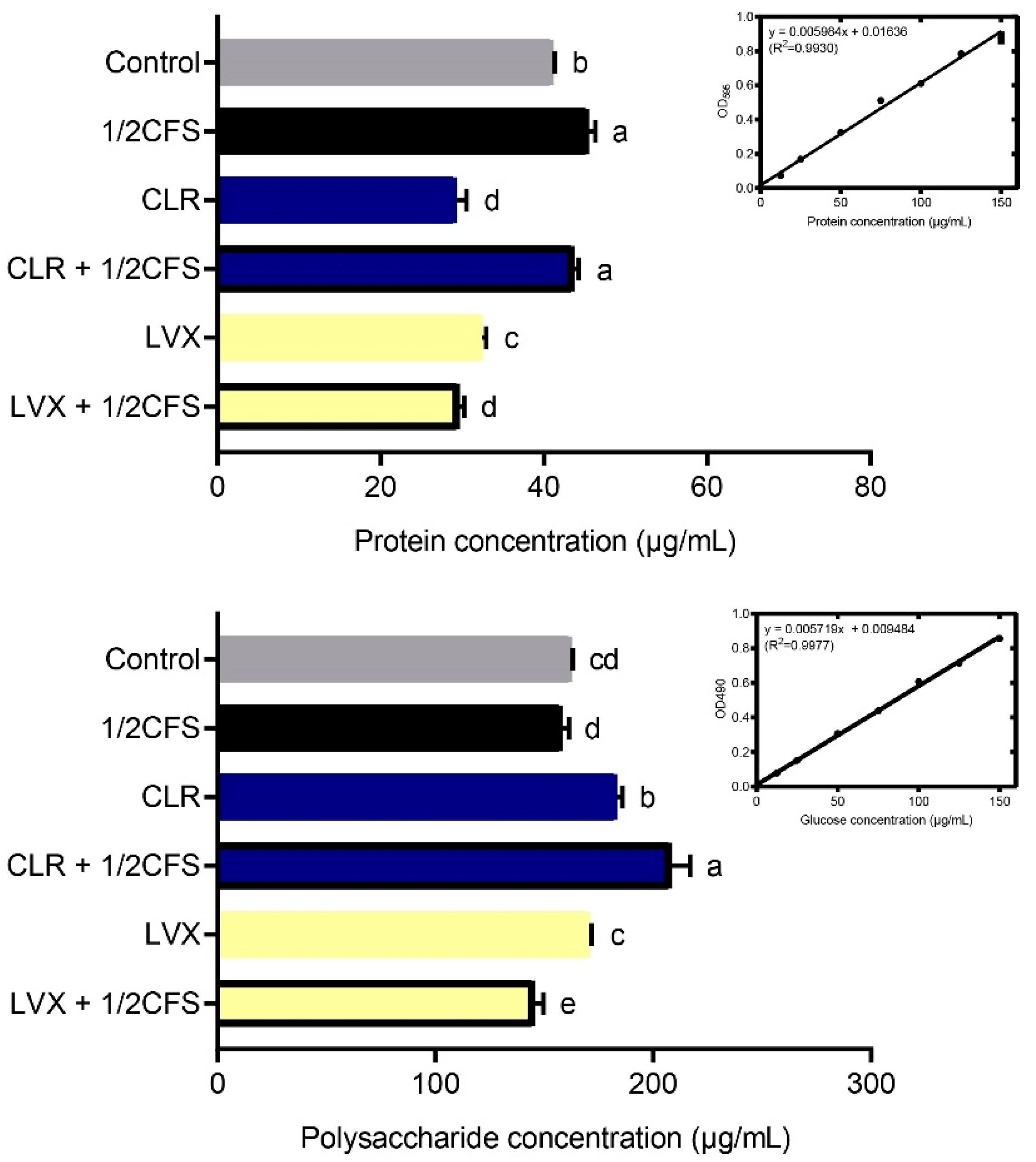
3.5. Combination of CFS and Different Antibiotics Can Affect the Protein and Polysaccharide Content of H. pylori Mature Biofilm EPS
3.6. SEM Observation of H. pylori Mature Biofilm Cell Morphology
3.7. CLSM Observation of H. pylori Mature Biofilm Structure
3.8. Combined Treatment with CFS and Different Antibiotics Can Affect Virulence Gene Expression in H. pylori Biofilm Cells
3.9. Combined Treatment with CFS and Different Antibiotics Can Inhibit Live H. pylori Mature Biofilm Cell Dispersion
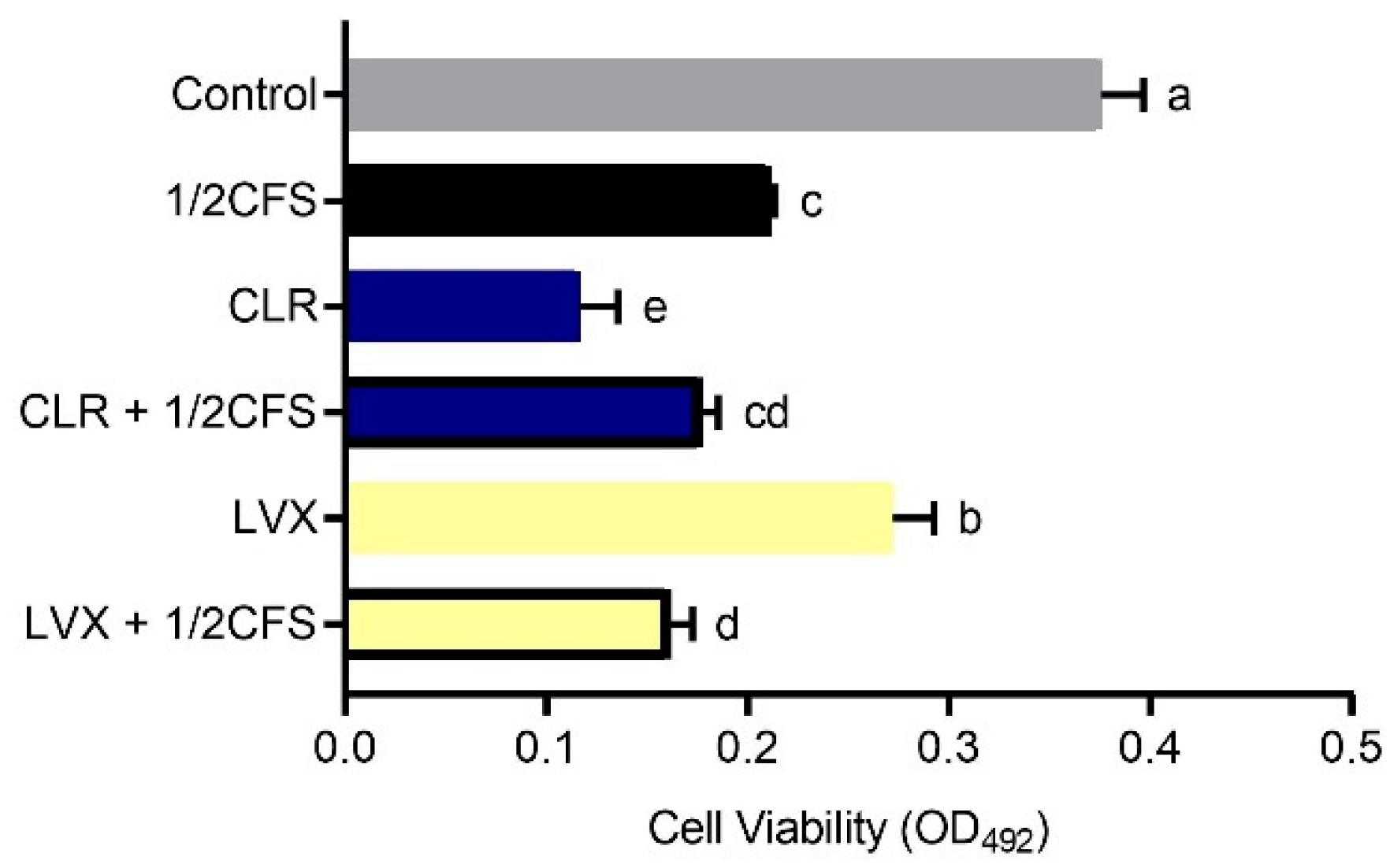
3.10. Combined Treatment with CFS and Different Antibiotics Can Influence the Viability of H. pylori Planktonic Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Connor, A.; O’Morain, A.O.C.A.; Ford, A.O.A.C. Population screening and treatment of Helicobacter pylori infection. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Wroblewski, L.E.; Peek, R.M.; Wilson, K.T. Helicobacter pylori and Gastric Cancer: Factors That Modulate Disease Risk. Clin. Microbiol. Rev. 2010, 23, 713–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, J.; Yang, H. Using Probiotics as Supplementation for Helicobacter pylori Antibiotic Therapy. Int. J. Mol. Sci. 2020, 21, 1136. [Google Scholar] [CrossRef] [Green Version]
- Hathroubi, S.; Servetas, S.L.; Windham, I.; Merrell, D.S.; Ottemann, K.M. Helicobacter pylori Biofilm Formation and Its Poten-tial Role in Pathogenesis. Microbiol. Mol. Biol. Rev. 2018, 82, e00001–e00018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Chen, X.; Shen, Y.; Li, H.; Zou, Y.; Yuan, G.; Hu, P.; Hu, H. Mucus penetration enhanced lipid polymer nanoparticles improve the eradication rate of Helicobacter pylori biofilm. J. Control. Release 2019, 300, 52–63. [Google Scholar] [CrossRef]
- Mathur, H.; Field, D.; Rea, M.C.; Cotter, P.D.; Hill, C.; Ross, R.P. Fighting biofilms with lantibiotics and other groups of bacteriocins. npj Biofilms Microbiomes 2018, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yang, H. Transcriptome Analysis of the Clostridioides difficile Response to Different Doses of Bifidobacterium breve. Front. Microbiol. 2020, 11, 1863. [Google Scholar] [CrossRef] [PubMed]
- Barzegari, A.; Kheyrolahzadeh, K.; Khatibi, S.M.H.; Sharifi, S.; Memar, M.Y.; Vahed, S.Z. The Battle of Probiotics and Their Derivatives against Biofilms. Infect. Drug Resist. 2020, 13, 659–672. [Google Scholar] [CrossRef] [Green Version]
- Moghadam, S.S.; Khodaii, Z.; Zadeh, S.F.; Ghooshchian, M.; Aghmiyuni, Z.F.; Shabestari, T.M.; Ghoshchiyan, M. Synergistic or Antagonistic Effects of Probiotics and Antibiotics- Alone or in Combination- on Antimicrobial-Resistant Pseudomonas aeruginosa Isolated from Burn Wounds. Arch. Clin. Infect. Dis. 2018, 13, 63121. [Google Scholar] [CrossRef] [Green Version]
- Miao, J.; Xin, L.; Wu, Z.; Guo, B. Studies on Modified MRS Eliminating the Inhibited Growth of Helicobacter pylori Caused by MRS Self. J. Dairy Sci. Technol. 2005, 4, 154–156. (In Chinese) [Google Scholar]
- De Souza, B.M.S.; Borgonovi, T.F.; Casarotti, S.N.; Todorov, S.D.; Penna, A.L.B. Lactobacillus casei and Lactobacillus fermentum Strains Isolated from Mozzarella Cheese: Probiotic Potential, Safety, Acidifying Kinetic Parameters and Viability under Gastrointestinal Tract Conditions. Probiotics Antimicrob. Proteins 2019, 11, 382–396. [Google Scholar] [CrossRef]
- Zuo, F.; Yu, R.; Feng, X.; Chen, L.; Zeng, Z.; Khaskheli, G.B.; Ma, H.; Chen, S. Characterization and in vitro properties of po-tential probiotic Bifidobacterium strains isolated from breast-fed infant feces. Ann. Microbio. 2015, 66, 1027–1037. [Google Scholar] [CrossRef]
- Al-Malkey, M.K.; Ismeeal, M.C.; Al-Hur, F.J.A.; Mohammed, S.W.; Nayyef, H.J. Antimicrobial effect of probiotic Lactobacillus spp. on Pseudomonas aeruginosa. J. Contemp. Med. Sci. 2017, 3, 218–223. [Google Scholar]
- Bomfim, V.B.; Neto, J.H.P.L.; Leite, K.S.; de Andrade Vieira, É.; Iacomini, M.; Mellinger-Silva, C.; Dos Santos, K.M.; Cardarelli, H.R. Partial characterization and antioxidant activity of exopolysaccharides produced by Lactobacillus plantarum CNPC003. LWT 2020, 127, 109349. [Google Scholar] [CrossRef]
- Pelyuntha, W.; Chaiyasut, C.; Kantachote, D.; Sirilun, S. Cell-free supernatants from cultures of lactic acid bacteria isolated from fermented grape as biocontrol againstSalmonellaTyphi andSalmonellaTyphimurium virulence via autoinducer-2 and biofilm interference. PeerJ 2019, 7, e7555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yang, H. Effect of Bifidobacterium breve in Combination with Different Antibiotics on Clostridium difficile. Front. Microbiol. 2018, 9, 2953. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Cai, Y.; Chen, Z.; Gao, S.; Geng, X.; Li, Y.; Li, Y.; Jia, J.; Sun, Y. Bifunctional enzyme SpoT is involved in biofilm for-mation of Helicobacter pylori with multidrug resistance by upregulating efflux pump Hp1174 (gluP). Antimicrob. Agents Chemother. 2018, 62, e00957-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Liu, X.M.; Tian, F.; Zhang, Q.; Zhang, H.P.; Zhang, H.; Chen, W. Antagonistic activities of lactobacilli against Helicobacter pylori growth and infection in human gastric epithelial cells. J. Food Sci. 2012, 77, 9–14. [Google Scholar] [CrossRef]
- Cerca, N.; Martins, S.; Cerca, F.; Jefferson, K.K.; Pier, G.B.; Oliveira, R.; Azeredo, J. Comparative assessment of antibiotic susceptibility of coagulase-negative staphylococci in biofilm versus planktonic culture as assessed by bacterial enumeration or rapid XTT colorimetry. J. Antimicrob. Chemother. 2005, 56, 331–336. [Google Scholar] [CrossRef]
- Kang, J.; Liu, L.; Liu, Y.; Wang, X. Ferulic Acid Inactivates Shigella flexneri through Cell Membrane Destructieon, Biofilm Retardation, and Altered Gene Expression. J. Agric. Food Chem. 2020, 68, 7121–7131. [Google Scholar] [CrossRef]
- Franklin, M.J.; Chang, C.; Akiyama, T.; Bothner, B. New Technologies for Studying Biofilms. Microbiol. Spectr. 2015, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Wang, X.; Ling, F.; Wang, H.; Zhang, P.; Shao, S. Atractylodes lancea volatile oils attenuated Helicobacter pylori NCTC11637 growth and biofilm. Microb. Pathog. 2019, 135, 103641. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Bassler, B.L. Surviving as a Community: Antibiotic Tolerance and Persistence in Bacterial Biofilms. Cell Host Microbe 2019, 26, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Wang, C.; Chen, Z.; Xu, Z.; Li, H.; Li, W.; Sun, Y. Transporters HP0939, HP0497, and HP0471 participate in intrinsic multidrug resistance and biofilm formation in Helicobacter pylori by enhancing drug efflux. Helicobacter 2020, 25, e12715. [Google Scholar] [CrossRef]
- Thung, I.; Aramin, H.; Vavinskaya, V.; Gupta, S.; Park, J.Y.; Crowe, S.E.; Valasek, M.A. Review article: The global emergence of Helicobacter pylori antibiotic resistance. Aliment. Pharmacol. Ther. 2016, 43, 514–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coticchia, J.M.; Sugawa, C.; Tran, V.R.; Gurrola, J.; Kowalski, E.; Carron, M.A. Presence and density of Helicobacter pylori bio-films in human gastric mucosa in patients with peptic ulcer disease. J. Gastrointest. Surg. 2006, 10, 883–889. [Google Scholar] [CrossRef]
- Suez, J.; Zmora, N.; Segal, E.; Elinav, E. The pros, cons, and many unknowns of probiotics. Nat. Med. 2019, 25, 716–729. [Google Scholar] [CrossRef]
- Jung, S.; Park, O.J.; Kim, A.R.; Ahn, K.B.; Lee, D.; Kum, K.Y.; Yun, C.H.; Han, S.H. Lipoteichoic acids of lactobacilli inhibit En-terococcus faecalis biofilm formation and disrupt the preformed biofilm. J. Microbiol. 2019, 57, 310–315. [Google Scholar] [CrossRef]
- Song, H.; Zhang, J.; Qu, J.; Liu, J.; Yin, P.; Zhang, G.; Shang, D. Lactobacillus rhamnosus GG microcapsules inhibit Escherichia coli biofilm formation in coculture. Biotechnol. Lett. 2019, 41, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Xie, Q.; Xu, D.; Guo, Y.; Tao, X.; Wei, H.; Wan, C. Antagonistics of Lactobacillus plantarum ZDY2013 against Helicobacter pylori SS1 and its infection in vitro in human gastric epithelial AGS cells. J. Biosci. Bioeng. 2018, 126, 458–463. [Google Scholar] [CrossRef]
- Chen, M.E.; Su, C.H.; Yang, J.S.; Lu, C.C.; Hou, Y.C.; Wu, J.B.; Hsu, Y.M. Baicalin, Baicalein, and Lactobacillus Rhamnosus JB3 Alleviated Helicobacter pylori Infections in vitro and in vivo. J. Food. Sci. 2018, 83, 3118–3125. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Tsai, W.-H.; Wu, H.-Y.; Chen, C.-Y.; Yeh, W.-L.; Chen, Y.-H.; Hsu, H.-Y.; Chen, W.-W.; Chen, Y.-W.; Chang, W.-W.; et al. Probiotic Lactobacillus spp. act Against Helicobacter pylori-induced Inflammation. J. Clin. Med. 2019, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Tian, F.; Liu, X.; Zhao, J.; Zhang, H.-P.; Chen, W.; Zhang, H. In Vitro screening of lactobacilli with antagonistic activity against Helicobacter pylori from traditionally fermented foods. J. Dairy Sci. 2010, 93, 5627–5634. [Google Scholar] [CrossRef]
- Homan, M.; Orel, R. Are probiotics useful in Helicobacter pylori eradication? World J. Gastroenterol. 2015, 21, 10644–10653. [Google Scholar] [CrossRef]
- Kadkhodaei, S.; Siavoshi, F.; Akbari Noghabi, K. Mucoid and coccoid Helicobacter pylori with fast growth and antibiotic re-sistance. Helicobacter 2020, 25, e12678. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.S.; Hur, J.W.; Yu, M.A.; Cheigh, C.I.; Kim, K.N.; Hwang, J.K.; Pyun, Y.R. Antagonism of Helicobacter pylori by bacteri-ocins of lactic acid bacteria. J. Food Prot. 2003, 66, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, P.; Shen, Y.; Zou, Y.; Yuan, G.; Hu, H. Rhamnolipid-involved antibiotics combinations improve the eradication of Helicobacter pylori biofilm in vitro: A comparison with conventional triple therapy. Microb. Pathog. 2019, 131, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Hanafi, A.; Lee, W.C.; Loke, M.F.; Teh, X.; Shaari, A.; Dinarvand, M.; Lehours, P.; Mégraud, F.; Leow, A.H.R.; Vadivelu, J.; et al. Molecular and Proteomic Analysis of Levofloxacin and Metronidazole Resistant Helicobacter pylori. Front. Microbiol. 2016, 7, 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salama, N.R.; Hartung, M.L.; Muller, A. Life in the human stomach: Persistence strategies of the bacterial pathogen Helicobacter pylori. Nat. Rev. Microbiol. 2013, 11, 385–399. [Google Scholar] [CrossRef]
- Urrutia-Baca, V.H.; Escamilla-García, E.; De La Garza-Ramos, M.A.; Tamez-Guerra, P.; Gomez-Flores, R.; Urbina-Ríos, C.S. In Vitro Antimicrobial Activity and Downregulation of Virulence Gene Expression on Helicobacter pylori by Reuterin. Probiotics Antimicrob. Proteins 2018, 10, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Ciccaglione, A.F.; Di Giulio, M.; Di Lodovico, S.; Di Campli, E.; Cellini, L.; Marzio, L. Bovine lactoferrin enhances the effi-cacy of levofloxacin-based triple therapy as first-line treatment of Helicobacter pylori infection: An in vitro and in vivo study. J. Antimicrob. Chemother. 2019, 74, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Di Lodovico, S.; Napoli, E.; Di Campli, E.; Di Fermo, P.; Gentile, D.; Ruberto, G.; Nostro, A.; Marini, E.; Cellini, L.; Di Giulio, M. Pistacia vera L. oleoresin and levofloxacin is a synergistic combination against resistant Helicobacter pylori strains. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bugli, F.; Palmieri, V.; Torelli, R.; Papi, M.; De Spirito, M.; Cacaci, M.; Galgano, S.; Masucci, L.; Paroni Sterbini, F.; Vella, A.; et al. In vitro effect of clarithromycin and alginate lyase against Helicobacter pylori biofilm. Biotechnol. Prog. 2016, 32, 1584–1591. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequences (5′-3′) |
|---|---|
| cagA-F | GCAAGTGGTTTGGGTGGTGTAGG |
| cagA-R | CCGCCGAGATCATCAATCGTAGC |
| vacA-F | AGCGAGCGGGCGTTCTTTATTG |
| vacA-R | GGTATCCGTGCCAGCCTTAAACTC |
| 16s rRNA-F | GGCGACCTGCTGGAACATTACTG |
| 16s rRNA-R | CATCGTTTAGGGCGTGGACTACC |
| Antibiotics | MICs (μg/mL) |
|---|---|
| CLR | 0.063 |
| LVX | 0.250 |
| Combinations | FICs | Effect |
|---|---|---|
| CLR + CFS | 3 (0.125 μg/mL, 12.5%) | Antagonist |
| LVX + CFS | 1 (0.125 μg/mL, 6.25%) | Additive |
| Group | Live Cells (nm) | Dead Cells (nm) |
|---|---|---|
| Control | 12.7 ± 1.5a | 2.3 ± 0.6B |
| 1/2CFS | 6.7 ± 0.6c | 1.7 ± 0.6B |
| CLR | 7.3 ± 0.6c | 5.7 ± 2.1A |
| CLR + 1/2CFS | 11.7 ± 1.2ab | 2.7 ± 0.6AB |
| LVX | 9.3 ± 1.5bc | 2.0 ± 1.0B |
| LVX + 1/2CFS | 6.7 ± 1.2c | 4.3 ± 1.5AB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, J.; Yang, H. In Vitro Effects of Lactobacillus plantarum LN66 and Antibiotics Used Alone or in Combination on Helicobacter pylori Mature Biofilm. Microorganisms 2021, 9, 424. https://doi.org/10.3390/microorganisms9020424
Ji J, Yang H. In Vitro Effects of Lactobacillus plantarum LN66 and Antibiotics Used Alone or in Combination on Helicobacter pylori Mature Biofilm. Microorganisms. 2021; 9(2):424. https://doi.org/10.3390/microorganisms9020424
Chicago/Turabian StyleJi, Jianfu, and Hong Yang. 2021. "In Vitro Effects of Lactobacillus plantarum LN66 and Antibiotics Used Alone or in Combination on Helicobacter pylori Mature Biofilm" Microorganisms 9, no. 2: 424. https://doi.org/10.3390/microorganisms9020424
APA StyleJi, J., & Yang, H. (2021). In Vitro Effects of Lactobacillus plantarum LN66 and Antibiotics Used Alone or in Combination on Helicobacter pylori Mature Biofilm. Microorganisms, 9(2), 424. https://doi.org/10.3390/microorganisms9020424